


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
水稻斑点叶突变体 spl21的鉴定与基因定位
[宋莉欣1, 2  , 黄奇娜
, 黄奇娜1 , 奉保华1 , 施勇烽1 , 张晓波1 , 徐霞1 , 王惠梅1 , 李小红1 , 赵宝华2, * , 吴建利1, * ]
, 黄奇娜, 吴建利]
|
|


第一作者联系方式: E-mail: songlixin321@foxmail.com
通过双环氧丁烷(diepoxybutane)诱变籼稻品种IR64获得一个稳定遗传的红褐色斑点叶突变体 spl21( spotted-leaf 21)。大田条件下, 突变体播种后约2周叶片上开始出现红褐色斑点, 随后部分斑点融合, 从叶尖开始发黄枯萎, 并沿叶片两侧边缘向下扩散, 严重时叶片大部分或整体枯死。突变体 spl21与野生型IR64相比, 其株高、穗长、有效穗数、实粒数、结实率和千粒重等农艺性状均显著降低。组织化学分析表明, 叶片斑点处及周围有H2O2沉积。突变还导致叶绿素 a、叶绿素 b和类胡萝卜素含量极显著降低, 叶片光合能力明显下降; 此外, 突变体中CAT、SOD、APX和可溶性蛋白含量均极显著降低, POD活性则极显著升高。遗传分析表明, 突变体表型受1对隐性核基因控制。通过图位克隆法最终将该基因定位于第12染色体长臂下端介于InDel-8和RM28746之间约87 kb的区段内, 暂名 spl21( t), 本研究为该基因的克隆与功能研究奠定了基础。
The rice spotted-leaf 21 mutant ( spl21) was isolated from a diepoxybutane-induced IR64 mutant bank. Under field conditions, the red-brown spots appeared on the leaves of mutant seedlings in two weeks after sowing. Subsequently, a portion of spots merged and the leaf tips became yellowish, wilted and spread downwards along both edges of the leaf blade leading to the death of the whole leaf blade when the symptom was severe. Accumulation of H2O2 was detected in and around the spots. Major agronomic traits including plant height, length of panicle, number of panicles, number of filled grains, seed setting-rate, and 1000-grain weight were markedly affected in the mutant. The contents of chlorophyll a, b, carotenoid and photosynthetic parameters were significantly reduced in the mutant as compared with the wild type. Furthermore, the activities of CAT, SOD, APX, and soluble protein contents were significantly lower than those of the wild type while the activity of POD was apparently higher than that of the wild type. The mutant trait was controlled by a single recessive nuclear gene, tentatively termed spl21( t), located on the long arm of chromosome 12. The population and data achieved in the present study would facilitate the isolation and functional analysis of spl21( t).
植物斑点叶(spotted leaf)突变体是在没有明显逆境、机械和农药损伤或者病原菌侵染的条件下, 由植物体在叶片或叶鞘上自发形成斑点的突变体[1]。斑点叶突变体分为两类, 一类是斑点处没有发生细胞死亡, 色斑仅仅是由于色素的沉积形成, 例如spl30[2]; 另一类是斑点处发生细胞死亡, 类似于无毒病原菌侵染造成的损伤, 因此通常又称为类病斑突变体(lesion mimic mutant), 在多种植物中已有广泛的报道[2, 3, 4, 5, 6, 7, 8]。
在水稻中, 斑点叶性状虽然复杂多样, 包括不同色泽、形状和大小的斑点(红色、褐色、橙黄色和白色), 不同的分布位置(叶片、叶鞘、枝梗和谷壳), 但其遗传控制相对简单, 绝大多数受单隐性基因控制, 少数受单显性基因控制, 双基因控制和母本遗传的情况较为少见[1]。迄今为止, 已经报道了约80个水稻斑点叶基因, 分布于除第9染色体外的其他11条染色体上, 但不少斑点叶基因的等位性还有待测定[8, 9, 10, 11, 12, 13]。
目前, 已经克隆了18个水稻斑点叶基因, 包括spl7[14]、spl11[15]、spl18[16]、spl28[17]、OsLSD1[18]、OsACDR1[19]、OsNPR1[20]、OsPtila[21]、RLIN1[22]、nls1-1D[23]、cea62[24]、spl5[25]、OsLMS[26]、noe1[27]、fgl[28]、sl[29]、RLS1[30]和LMR[31]。研究发现, 包括水稻等植物中已克隆的基因, 编码许多不同类型的蛋白, 例如锌指蛋白[5, 18]、热激转录蛋白[14]、膜相关蛋白[17]、离子通道调节蛋白等[32]; 涉及多种生理生化代谢途径, 比如卟啉代谢[33]、脂肪酸/脂质代谢[34]、酚类物质代谢[5]以及泛素化作用等[15]。由此可见, 水稻斑点叶基因具有广泛的生物学功能, 其机制也十分复杂。充分挖掘、鉴定更多的突变体, 分离相关基因并阐明各个基因的功能, 才能更加全面了解斑点叶或类病斑发生的机理, 同时也为研究水稻等植物的程序性细胞死亡和抗病机制、探讨类病斑抗性的育种价值提供参考。
本研究利用籼稻IR64经过双环氧丁烷(Diepoxybutane)诱变获得一个稳定遗传的斑点叶突变体spl21[35], 其农艺性状和生理生化特征均发生了显著变化。突变体褐色斑点叶性状受1对隐性核基因控制, 该基因暂被命名为spl21(t), 定位于第12染色体长臂上, 是一个不同于已克隆的新斑点叶基因。研究结果为该斑点叶基因的克隆和功能分析奠定了基础。
突变体spl21经过多代自交, 其褐色斑点表型在浙江和海南均能稳定遗传。2014年将该spl21和野生型IR64种植于杭州富阳实验基地, 每个材料4行共24株, 株行距17 cm × 20 cm, 成熟期分别调查株高、穗长、单株有效分蘖数、结实率、千粒重等农艺性状, 3次重复, 取其平均值。
采用Arnon和Wellburn等[36, 37]的方法, 在大田条件下, 分别测定苗期、分蘖期和抽穗期的野生型IR64、突变体spl21带斑点和不带斑点叶片中叶绿素a、叶绿素b和类胡萝卜素的含量。处理过的样品分别测定在470 nm、645 nm和663 nm三个波长下的吸光值, 然后根据公式分别计算spl21和IR64的色素含量。
从田间取同时期的spl21红褐斑叶片、无斑叶片和对照IR64, 采用Thordal-Christensen等[38]的方法, 利用二氨基联苯胺(diamino benzidine, DAB)染色检测突变体类病斑部位过氧化氢发生情况。
用便携式光合仪Li-6400进行叶片光合作用指标的测定。在大田条件下, 天气晴朗的上午8:00— 10:00分别测定分蘖盛期突变体spl21和野生型IR64的叶片净光合速率(photosynthetic rate, Pn)、气孔导度(stomatal conductance, Gs)、胞间CO2浓度(intercellular CO2concentration, Ci)、蒸腾速率(transpiration rate, Tr)。使用光合仪Li-6400的红蓝光源, 光量子密度为1200 μ mol m-2 s-1, 流速为500 μ mol s-1。每个部位测定3片叶, 取平均值。
参照赵世杰等[39]方法, 从田间分别选取分蘖盛期spl21和IR64的主茎倒二叶和倒三叶, 抽提后, 对其过氧化氢酶(catalase, CAT)、过氧化物酶(peroxidase, POD)、超氧化物歧化酶(superoxide dismutase, SOD)、抗坏血酸过氧化物酶(ascorbate peroxidase, APX)、可溶性蛋白含量(soluble protein contents)和丙二醛(malonaldehyde, MDA)含量等与衰老生理生化相关指标进行测定。
以spl21为母本, 分别以Moroberekan、LH422和Jefferson为父本进行杂交, 观察F1的表型; 调查来源于spl21/Moroberekan、spl21/LH422和spl21/Jefferson三个组合的F1自交获得的F2群体性状的分离情况; 同时调查来源于spl21/Moroberekan的3个F3分离株系, 用于遗传验证。
采用简易法[39]从水稻叶片中提取基因组DNA, 用于PCR。PCR体系包括1 µ g的模板DNA、1 µ L (10 µ mol L-1)引物、0.2 µ L (10 mmol L-1) dNTPs、1 µ L 10× PCR缓冲液、0.25 µ L Taq DNA聚合酶(2 U μ L-1), 加ddH2O补足10 µ L。在Biometra PCR仪上进行扩增反应, 反应条件为94℃预变性2 min; 94℃ 30 s, 55℃ 30 s, 72℃ 40 s, 共35个循环; 72℃延伸5 min。反应产物经6%非变性聚丙烯酰胺凝胶电泳、银染显色, 分析检测样品间的多态性。
采用DNA分池法(bulk segregation analysis)根据F2表型随机选取红褐斑点叶与正常叶单株各10株, 每单株以等量叶片混合, 各自提取构成斑点叶DNA池与正常叶DNA池。
初定位采取两个DNA池和随机选取的36株红褐色斑点叶单株。选取均匀分布于12条染色体上的微卫星(SSR)标记筛选亲本spl21与Moroberekan之间的多态性, 亲本间有多态的标记进一步用于两池的筛选, 然后两池间有多态的标记用随机选取的36株红褐色斑点叶单株进一步验证。所用SSR标记的引物序列下载自Gramene数据库(http://www.gramene.org/), 引物由上海生工生物工程有限公司合成。
根据初步定位结果, 采用841个F2褐色斑点叶单株进行精细定位。在目标区间进行SSR标记加密, 并利用国际公共数据库RGP (http://rgp.dna.affrc.go.jp/E/toppage.html), Gramene (http://gramene.org/genome_browser/index.html)和中国科学院华大基因研究中心(http://rice.genomics.org.cn/rice/index2.jsp)已公布的粳稻日本晴和籼稻93-11全基因组序列, 比对分析日本晴和93-11间序列的差别, 借助生物学软件Primer Premier5.0和DNAStar5.0设计插入/缺失(InDel)引物, 筛选多态性标记用于精细定位斑点叶基因。引物合成、PCR条件和电泳分析条件同上。
在杭州富阳大田自然条件下, 突变体spl21播种约2周后, 从第一叶叶尖开始出现大小不同、形状不规则的红褐色斑点, 然后逐渐扩散至全叶, 到后期部分斑点融合, 从叶尖开始枯萎, 并沿叶片两侧边缘向下扩散, 严重时叶片大部分或整体枯死(图1)。突变体spl21的抽穗期比野生型晚约10 d, 突变体的穗长与野生型相比存在显著差异; 而株高、有效穗数、实粒数、结实率和千粒重与野生型存在极显著差异(表1)。突变体较矮的原因是第1、第2、第3、第4和第5节间比野生型极显著缩短(图2)。
 | 图1 突变体表型A: 野生型IR64幼苗; B: 突变体spl21幼苗; C: 苗期叶片 (左: IR64; 右: spl21); D: 分蘖期IR64和spl21。Fig. 1 Phenotype of the mutantA: seedling of IR64; B: seedling of the mutant; C: leaves at the seedling stage (Left: IR64; Right: spl21); D: plants of the wild type and the mutant at the tillering stage. |
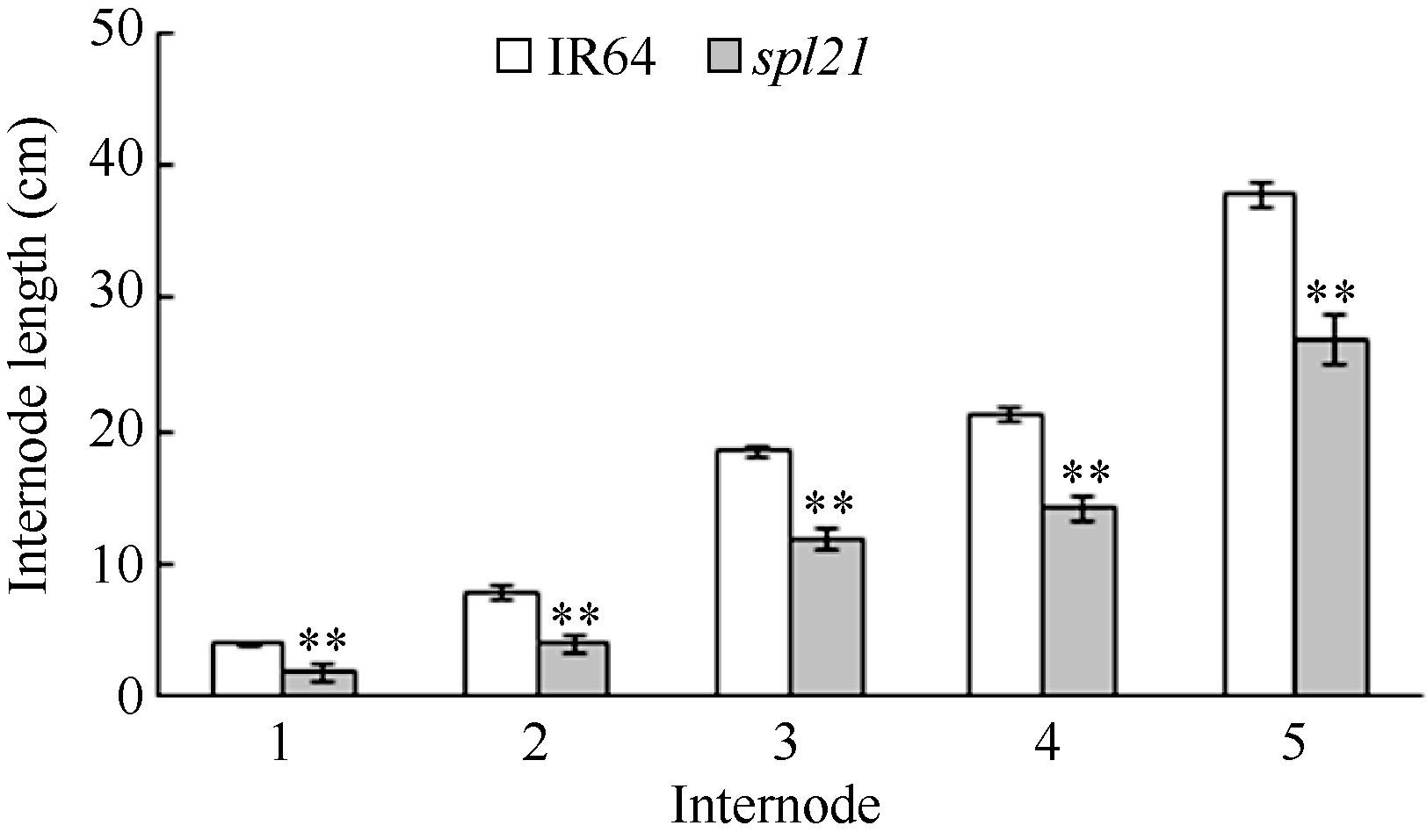 | 图2 野生型IR64与突变体spl21的节间长度Fig. 2 Internode length of the wild type IR64 and mutantspl21 |
光合色素含量测定结果表明, 苗期突变体spl21不带斑点叶片与野生型IR64叶片间叶绿素a、叶绿素b、类胡萝卜素含量均无显著差异; 而spl21带斑点叶片与IR64相比, 叶绿素a、叶绿素b、类胡萝卜素的含量均极显著下降, 但与IR64相比, 叶绿素a/b比值没有差异(表2)。分蘖期和抽穗期的结果和苗期一致(见附表1、附表2和附表3)。由于不同时期突变体叶绿素a/b比值与野生型IR64均没有差异, 说明突变体光合色素含量下降可能与总活性细胞有关, 而与叶绿素代谢缺陷无关。
| 表1 突变体和野生型的农艺性状 Table 1 Performance of agronomic traits in the mutant and wild type |
| 表2 苗期叶片光合色素含量 Table 2 Pigment contents at the seedling stage |
二氨基联苯胺(DAB)染色可以在细胞水平上检测活性氧物质的产生与积累。如图3显示, spl21不带斑点叶片和野生型IR64叶片没有被DAB染色(图3-A~D); spl21带斑点叶片斑点处及周围被染成红棕色(图3-E, F)。说明突变体中具有较多的过氧化氢产生。
突变体spl21分蘖盛期主茎倒二叶(无斑点)的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)均与野生型无差异(图4); 而倒三叶(有斑点)的净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)与野生型IR64相比均极显著降低, 但胞间CO2浓度(Ci)则极显著升高(图4)。以上结果说明, 随着突变体spl21在分蘖盛期叶片枯萎表型的加重, 叶片的水分含量相应减少, 气孔导度降低, 蒸腾作用相应减弱, 而胞间CO2浓度却开始积累增加, 最终导致叶片的光合速率降低。
 | 图3 突变体spl21和野生型IR64叶片DAB染色结果A: IR64染色之前; B: IR64染色之后; C: spl21无斑叶片(spl21-NO)DAB染色之前; D: spl21无斑叶片(spl21-NO)DAB染色之后; E: spl21褐斑叶片(spl21-SPL)DAB染色之前; F: spl21褐斑叶片(spl21-SPL)DAB染色之后。Fig. 3 DAB staining of leaves in the mutant spl21 and wild type IR64A: IR64 before DAB staining; B: IR64 after DAB staining; C: spl21-NO before DAB staining; D: spl21-NO after DAB staining; E: spl21-SPL before DAB staining; F:spl21-SPL after DAB staining. |
突变体spl21与野生型IR64相比, 主茎倒二叶(无褐斑)的CAT、POD、SOD、APX和可溶性蛋白含量均无差异; 但倒三叶(有褐斑)的CAT、SOD、APX和可溶性蛋白含量均极显著降低(图5-A, C, D, E), POD活性则极显著升高(图5-B)。突变体spl21主茎倒二叶、倒三叶的MDA含量均略高于野生型IR64, 但无显著差异(图5-F), 说明突变体主要活性氧清除酶活性的改变可能与细胞死亡有关, 但MDA含量未明显改变, 又不同于典型的叶片衰老。因此, spl21是否是一个早衰的突变体还有待于进一步研究。
以spl21为母本, Moroberekan、LH422和Jefferson为父本分别构建了spl21/Moroberekan、spl21/LH422和spl21/Jefferson三个群体, 考察各自的F1植株表型, 发现所有的F1都表现为正常绿色叶, 说明褐色斑点叶性状受隐性基因控制。观察3个群体的F2单株, 发现其均出现明显的分离, 分别表现亲本的性状, 没有中间类型的植株。spl21/Moroberekan的F2群体中共考察824株, 其中野生型个体624株, 突变型个体200株, 经卡平方检验, 野生型与突变型个体的分离比符合3:1 (χ 2=2.606< χ 20.05=3.84); spl21/ LH422的F2群体中共考察524株, 其中野生型个体377株, 突变型个体147株, 经卡平方检验, 野生型与突变型个体的分离比符合3:1 (χ 2=0.233< χ 20.05= 3.84); spl21/Jefferson的F2群体中共考察705株, 其中野生型个体549株, 突变型个体156株, 经卡平方检验, 野生型与突变型个体的分离比也符合3:1 (χ 2=3.102< χ 20.05=3.84)。另外, 来源于spl21/Moroberekan的3个F3分离株系的正常株和突变株比例也都符合3:1 (见附表4), 以上结果说明该突变体性状受1对隐性核基因控制(表3)。
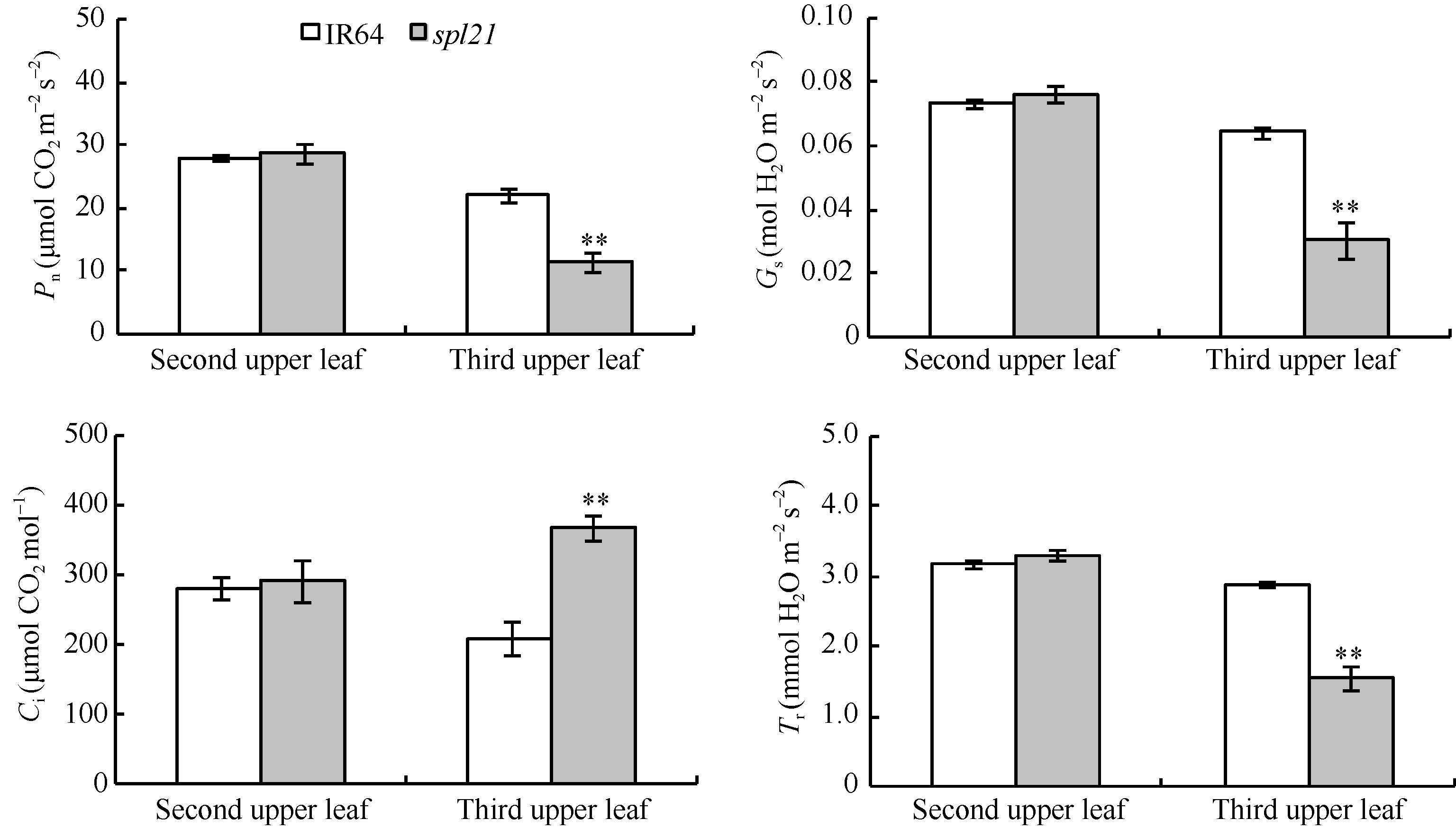 | 图4 野生型IR64和突变体spl21光合指标* * 极显著差异(P≤ 0.01)。* * Significant at P≤ 0.01.Fig. 4 Photosynthetic parameters of the wild type IR64 and mutant typespl21 |
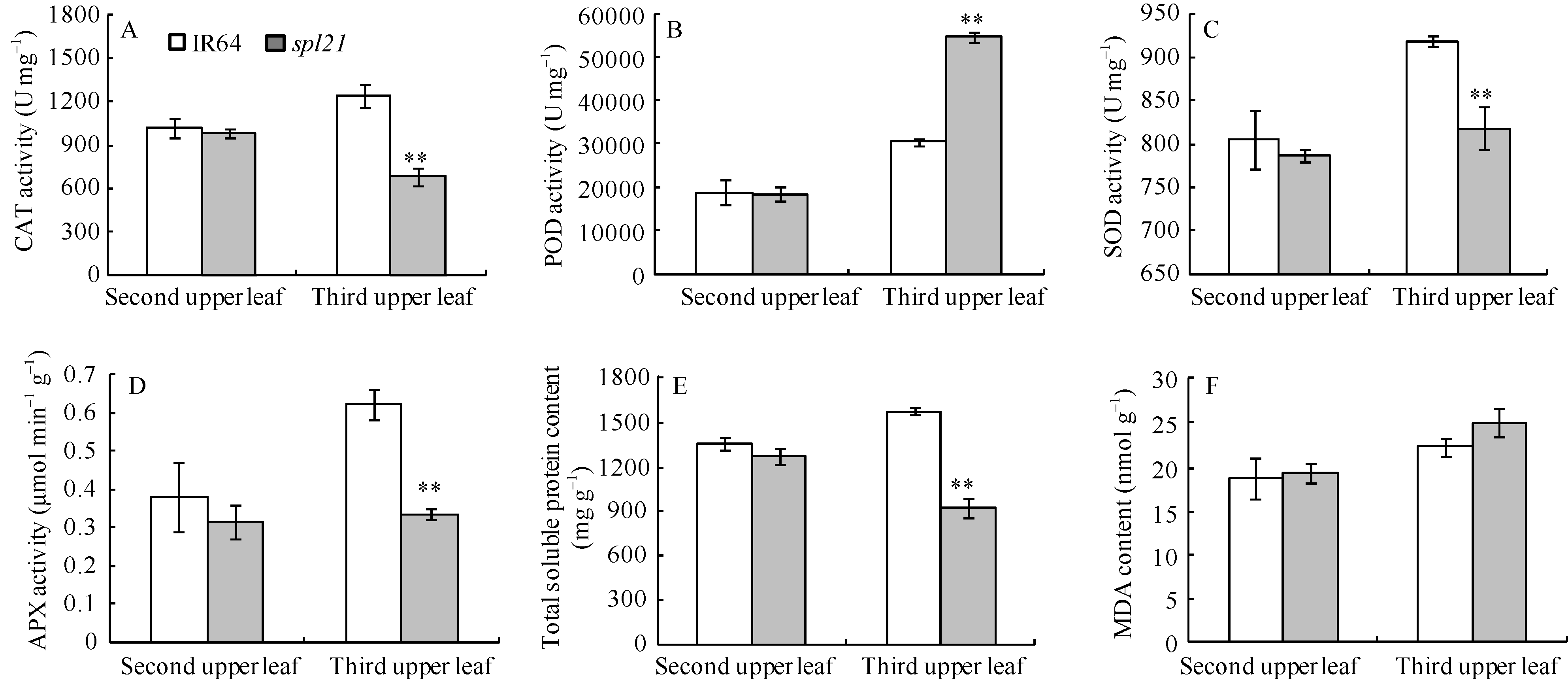 | 图5 野生型IR64和突变体spl21生理生化指标A: 过氧化氢酶(CAT)活性; B: 过氧化物酶(POD)活性; C: 超氧化物歧化酶(SOD)活性; D: 抗坏血酸过氧化物酶(APX)活性; E: 可溶性蛋白含量; F: 丙二醛(MDA)含量。* * 极显著差异(P≤ 0.01)。Fig. 5 Physiological and biochemical indexes of wild type IR64 and mutant typespl21A: CAT activity; B: POD activity; C: SOD activity; D: APX activity; E: total soluble protein content; F: MDA content. * * Significant at P≤ 0.01. |
| 表3 褐色斑点叶突变体spl21的遗传分析 Table 3 Genetic analysis of spl21 mutant |
在第12染色体上有7个SSR标记(RM28438、RM28466、RM28537、RM235、RM17、RM28828和RM28621)在亲本间和DNA池之间均有多态性, 随后用随机选取的36个斑点叶单株验证发现, RM235有2个交换单株, RM17和RM28828均只有1个交换单株, 表明该斑点叶基因与这3个标记紧密连锁, 初步表明该基因位于第12染色体长臂上, 暂名spl21(t)。
为精细定位该褐色斑点叶基因, 利用已有标记和新发展的SSR标记对F2群体中的另外841株突变型植株进行基因型分析, 最终将该褐色斑点叶基因定位于第12染色体长臂下端RM28723和RM28746之间约200 kb的区段内。根据籼粳稻序列差异, 借助生物学软件Primer Premier5.0和DNAStar5.0在RM28723和RM28746中间新发展了39个InDel标记(见附表5), 最终将spl21定位于InDel-8和RM28746之间约87 kb区间内(图6)。根据Gramene网站的注释, 该区间共有13个开放阅读框, 尚无斑点叶基因的报道, 因此, 本研究定位的斑点叶基因spl21(t)可能是一个新基因。
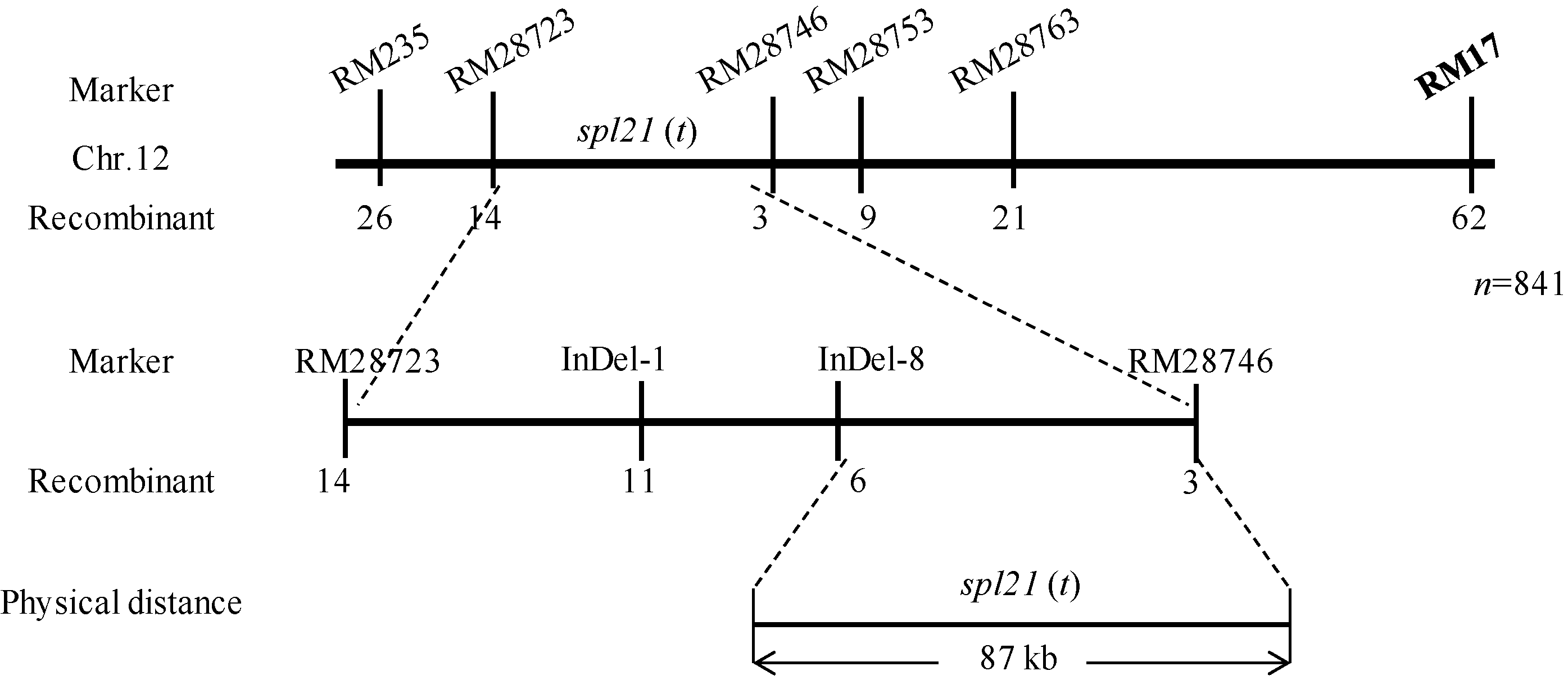 | 图6 spl21(t)在第12染色体上的物理定位Fig. 6 Physical location of spl21(t) on chromosome 12 |
目前, 国内外已经报道了大量水稻斑点叶突变体的研究, 表明水稻斑点叶突变体除产生色泽、形状和大小不同斑点外, 还通常伴随着重要农艺性状、光合色素代谢以及抗病性等的改变[1, 4, 13]。
本研究通过双环氧丁烷(diepoxybutane)诱变获得一个褐色斑点叶突变体, 该突变体在播种约2周后开始出现红褐色斑点, 之后部分斑点融合并从叶尖开始枯萎, 严重时叶片大部分或整体枯死, 抽穗后突变体植株衰老明显。突变体spl21在苗期、分蘖期和抽穗期与野生型IR64相比, 叶绿素a、叶绿素b、类胡萝卜素的含量均极显著下降, 但叶绿素a/b的比值却无显著差异, 说明突变体斑点的产生可能与测定时的活细胞数目有关, 而与叶绿素代谢缺陷无关, 这与之前报道的spl30(t)明显不同[2]。另外, 突变体spl21叶片净光合速率、气孔导度和蒸腾速率均比野生型极显著降低, 但胞间CO2浓度有所增加, 其原因与突变体大部分斑点叶片快速枯萎, 叶片失水严重有关。突变体植株在抽穗后叶片快速枯萎, 似有早衰的迹象, 因此我们怀疑spl21可能是一个早衰突变体。在分蘖盛期突变体spl21倒三叶的CAT、SOD和APX活性极显著降低, 此时叶片已经产生斑点但还未枯萎, 不过可能已经不能有效清除不断积累的过氧化氢或其他活性氧自由基, 导致加速叶片衰老; 同时, 突变体的可溶性蛋白含量极显著降低, 不足以提供植物体生长所需, 也会导致叶片加速衰老死亡; 突变体倒三叶的POD活性极显著升高, 可能是斑点的产生导致细胞壁及细胞器机械损伤, 从而削弱结合态POD的结合能力, 致使较多结合态的POD从细胞壁上游离出来, 引起游离态POD的大幅上升[41]。有趣的是突变体倒三叶MDA含量升高, 但未达到显著程度, 有别于植物正常衰老过程。因此, spl21是否是一个早衰的突变体还有待进一步研究。
到目前为止, 在第12染色体上已经定位了sl[29]、spl11[15]、spl29(t)[42]、spl30(t)[43]、syl1[44]、splt[45]、spl31[46]、lmm3[47]等8个斑点叶相关基因, 其中sl和spl11基因已被克隆。syl1与spl(t)位于第12染色体短臂上, lmm3、spl29(t)与spl31相距较近, 都位于第12染色体着丝粒附近, spl30(t)位于长臂着丝粒与标记RM3739之间, 而本研究定位的spl21(t)靠近长臂末端, 两者之间相隔约100 Mb, 因此, 初步确定spl21(t)是一个新斑点叶基因。本研究定位的spl21(t)位于第12染色体InDel-8和RM28746之间约87 kb区段内, 该区段内共13个开放阅读框, 其中LOC_ Os12g42290、LOC_Os12g42320、LOC_Os12g42390、LOC_Os12g42410和LOC_Os12g42360编码不同的假定蛋白, LOC_Os12g42300编码钾离子流出逆向转运蛋白, LOC_Os12g42310编码丝/苏蛋白磷酸酶BSL2, LOC_Os12g42330编码舌下腺粘蛋白前体, LOC_Os12g42340编码含OsFBX464-F-box结构域的蛋白, LOC_Os12g42370编码未知结构域的GTP酶、LOC_Os12g42380编码表达蛋白、LOC_Os12g42400编码核转录因子Y亚基、LOC_Os12g42420编码DNA结合蛋白, 这些基因均未被克隆。目前, spl21(t)候选基因的鉴定正在进行中。
红褐斑突变体spl21属于扩散型斑点叶突变体, 该性状受一个新的单隐性核基因控制, 位于第12染色体长臂末端。该基因的突变除导致突变体株高、穗长、结实率以及千粒重等重要农艺性状显著降低外, 还导致光合色素和光合效率极显著降低。此外, 衰老相关指标包括活性氧自由基清除系统中的APX、CAT、SOD酶活性和可溶性蛋白含量均显著降低, 但MDA含量并未明显改变, 有别于正常的衰老过程。深入研究spl21(t)基因的功能将有助于对植物衰老现象本质的理解。
| 附表1 分蘖期叶片光合色素含量 Supplementary table 1 Pigment contents at the tillering stage |
| 附表2 抽穗期剑叶光合色素含量 Supplementary table 2 Pigment contents of flag leaves at the heading stage |
| 附表3 抽穗期倒二叶光合色素含量 Supplementary table 3 Pigment contents of the second upper leaf at the heading stage |
| 附表4 来源于spl21/Moroberenkan的F3分离株系的遗传分析 Supplementary table 3 Genetic analysis of F3 segregation lines derived from spl21/Moroberekan |
| 附表5 用于spl21(t)基因定位的分子标记 Supplementary table 3 Polymorphic markers used for spl21(t) gene mapping |
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|