

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
油菜素内酯对春玉米灌浆期叶片光合功能与产量的调控效应
[王庆燕1  , 管大海
, 管大海1, 2 , 潘海波1 , 李建民1 , 段留生1 , 张明才1, * , 李召虎1 ]
, 管大海, 李召虎|
|


第一作者联系方式: E-mail: wangqyan@aliyun.com
以玉米品种“郑单958”为材料, 在大田条件下, 采用植物生长调节物质油菜素内酯(brassinolide, BR)对苞叶和穗位叶喷施处理, 研究了BR对玉米穗位叶功能、籽粒灌浆及产量的调控作用。结果表明, 灌浆期随生育进程, 玉米穗位叶叶绿素含量、光合速率、磷酸烯醇式丙酮酸羧化酶(PEPCase)、1,5-二磷酸核酮糖羧化酶(RuBPCase)以及蔗糖磷酸合酶和蔗糖合酶的活性均显著下降。同时, 籽粒蔗糖含量显著降低, 但淀粉含量和粒重均显著增加。与对照相比, BR处理显著增加玉米穗位叶叶绿素含量, 提高光合速率, 增强PEPCase、RuBPCase、蔗糖磷酸合酶和蔗糖合酶的活性。BR处理显著增加籽粒蔗糖和淀粉积累, 提高玉米籽粒干物质积累。在产量构成上, BR显著缩短秃尖长度, 增加穗粒数和千粒重, 显著提高产量。本研究说明, 灌浆期喷施BR可提高玉米叶源的活性, 延长叶片光合功能持续期, 促进籽粒灌浆和物质积累, 从而实现增产。
A field experiment was conducted to investigate the effect of plant growth regulator brassinolide (BR) on the regulation of physiological activities of ear leaf, the grain filling and yield in spring maize ( Zea mays L., cv. Zhengdan 958). Husks and ear leaves were treated with BR at 15 days after anthesis by foliar spraying. The results showed that the chlorophyll content, photosynthetic rate and activities of PEP carboxylaes, RuBP carboxylase, sucrose synthase and sucrose phosphate synthase in ear leaf as well as source content in kernel were significantly reduced, while starch accumulation and dry weight of kernel were increased during grain filling. BR significantly increased the content of chlorophyll and photosynthetic rate, enhanced the activities of PEP carboxylaes and RuBP carboxylase, definitely resulting in enhanced CO2 assimilation. BR markedly increased the activities of sucrose synthase and sucrose phosphate synthase in ear leaf. And in kernel, the accumulation of sucrose and starch as well as the grain filling rate were accelerated by BR. BR treatment significantly reduced the length of ear barren tip, and increased the number of kernels per ear, which led to higher yield. These results suggest that BR treatment at filling stage may be a good strategy to get a high yield in maize, by enhancing activity of resource, delaying sencence and stimulating the flow of assimilate from the source to the sink.
华北平原是我国重要的玉米产区, 其生产的玉米占全国产量的33%以上[1]。该地区以灌溉农业为主, 由于常年过量的地下水开采, 导致水资源匮缺, 已严重妨碍了华北地区农业的持续发展。为了保护有限的水资源, 将耗水量大的冬小麦种植面积减少, 增加春玉米种植已成为大家关注的热点问题。然而, 由于华北地区春玉米灌浆期处于阴雨寡照、高温高湿的7月至8月, 易造成玉米早衰[2], 因此增强玉米叶片源活性, 加快灌浆速率, 延长灌浆期对该地区春玉米增产具有重要意义。
玉米籽粒发育过程受激素间平衡和协同作用调控, 退化玉米籽粒中生长素(IAA)水平较低, 与IAA水平有关的过氧化物酶及同工酶明显高于正常籽粒, 而败育籽粒中细胞分裂素(CTKs)低于正常水平[3]。赤霉素(GA)可以促进乳熟期同化物的调运, 脱落酸(ABA)可以促进玉米籽粒中同化物的卸载, 提高光合产物向籽粒中的运输[4]。油菜素内酯(BR)是一种高活性的甾醇类植物激素。众多研究发现, BR及其类似物的生理功能非常广泛, 涉及细胞伸长与分裂、光合作用、维管分化和逆境反应等[5, 6]。同时, BR对植物生殖生长也具有重要调控作用。Ye等[7]发现BR可以通过调控花粉数量、活性和散粉效率来调节拟南芥雄性可育性。Hartwig等[8]研究发现BR合成缺陷突变体和抑制剂均可导致玉米雄花的雌性化。Jiang等[9]发现BR在调控拟南芥花粉发育的同时, 对籽粒大小、形状和生物量积累也起着重要作用, 这表明BR可以促进籽粒生长, 增加籽粒生物量积累。Morinaka等[10]在研究BR对水稻籽粒的调控上也得到了相似结论。然而, 有关BR在春玉米灌浆期叶片功能与籽粒发育的调控研究鲜有报道。本文在大田条件下, 采用BR叶面喷施处理, 研究BR对春玉米灌浆期穗位叶功能和籽粒灌浆及产量的调控, 探讨BR对春玉米灌浆过程的调控效应, 为华北地区春玉米高产提供新的技术手段。
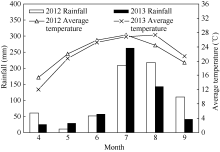
试验于2012— 2013年在河北省沧州市吴桥县中国农业大学吴桥试验站(37° 41′ N, 116° 37′ E)进行。试验地土质为沙壤土, 肥力中等, 含有机质2.5%、全氮0.16%、碱解氮179.2 mg kg-1、速效磷83.7 mg kg-1、速效钾161.0 mg kg-1。2年玉米生长季气象条件如图1。以郑单958为试验材料, 种植密度为60 000株 hm-2, 行距为60 cm, 采用春玉米单作, 播前深翻旋耕, 肥料全部基施, 施用量为纯N 220 kg hm-2、P2O5和K2O各90 kg hm-2。吐丝期标记吐丝时间一致的植株, 吐丝后15 d苞叶和穗位叶叶面喷施0.1 mg L-1 BR。试验采用随机区组设计, 重复5次, 小区面积6.0 m × 4.8 m。
 | 图1 2012-2013年玉米生长季月降雨量和平均温度Fig. 1 Monthly rainfall distribution (bar) and mean temperature (line) during maize growing season in 2012-2013 |
处理后每隔5 d取长势均匀的植株3株, 以果穗中部部分籽粒用于灌浆速率测定, 另一部分籽粒及穗位叶经液氮速冻, -20℃保存, 用于酶活性测定。
1.2.1 穗位叶叶绿素含量及光合速率测定 标记每区长势均匀的植株5株。处理后每隔5 d用叶绿素仪(SPAD-502, Konika Minolta Sensing Inc., Japan)测定标记植株穗位叶SPAD值。用LI-6400便携式光合作用测定系统(LI-COR, 美国), 设定开放式气路和365 µ mol mol-1 CO2, 测定穗位叶净光合速率(Pn)。
1.2.2 穗位叶PEPCase和RuBPCase活性测定
按李立人等[11]的方法并略加修改提取酶液。取玉米穗位叶0.5 g, 置研钵中, 加入3 mL预冷的提取缓冲液(100 mmol L-1 Tris-HCl, 5%甘油, 1% PVP, 1 mmol L-1 EDTA, 10 mmol L-1巯基乙醇, pH 8.2), 冰浴中研磨至匀浆, 于4℃下15 000× g离心20 min, 取上清液, 以分光光度法测定RuBPCase活性[12], 施教耐等[13]的方法测定PEPCase活性。
1.2.3 蔗糖合酶和磷酸蔗糖合酶活性测定 参考Dougles等[14]的方法稍作修改提取酶液。称取0.5 g叶片, 置预冷的研钵中, 分批加入5 mL提取缓冲液(100 mmol L-1 Tris-HCl, 5 mmol L-1 MgCl2, 2 mmol L-1 EDTA-Na, 2%乙二醇, 0.2%牛血清蛋白, 2% PVP, 5 mmol L-1 DTT, pH 7.0), 冰浴研磨成匀浆, 4℃下10 000× g离心20 min, 取上清液2 mL装入透析袋, 置于透析缓冲液(25 mmol L-1 Tris-HCl, 2.5 mmol L-1 MgCl2, 1 mmol L-1 EDTA-Na, 1%乙二醇, 1 mmol L-1 DTT, pH 7.0)中4℃过夜, 期间更换透析液3次, 将透析后酶液定容至5 mL备用。
参照苏丽英等[15]的方法稍作修改测定蔗糖合酶活性, 向测定试管中加入0.4 mL酶反应液, 0.1 mL UDPG和0.05 mL透析后的酶液, 加H2O至1 mL, 于30℃水浴反应10 min, 沸水浴3 min中止反应, 对照用蒸馏水代替UDPG。各反应试管中加入0.1 mL 2 mol L-1 NaOH, 沸水浴10 min, 冷却至室温。加3.5 mL 30% HCl, 1 mL 0.1%间苯二酚, 摇匀后于80℃保温10 min, 冷却后480 nm处比色, 测定蔗糖生成的量。记录并计算蔗糖合酶活性。
参考Wardlaw[16]的方法略做修改测定蔗糖磷酸合酶活性, 在酶反应液中用10 mol L-1果糖-6-磷酸代替果糖, 其余均按蔗糖合酶的方法测定。
1.2.4 籽粒灌浆干物质积累测定 取玉米中部籽100粒, 测定鲜重和体积; 然后于恒温干燥箱105℃杀青0.5 h, 80℃烘至恒重, 测定干重。
1.2.5 籽粒淀粉及蔗糖含量测定 取1.2.4中烘干籽粒, 粉碎, 过0.5 mm筛, 取0.5 g粉末, 用蒽酮比色法[17]测定籽粒总淀粉含量, 间苯二酚法[18]测定籽粒蔗糖含量。
1.2.6 产量及其构成因素 成熟期收获每区中间2行测产, 选取代表性果穗10穗, 室内考种, 测定穗长、穗粗、秃尖长、穗行数、行粒数和千粒重(14%含水量)等。
用SPPS 17.0统计分析, 采用t测验法进行显著性检验, Microsoft Excel 2003作图。
BR处理可显著缩短秃尖长, 增加玉米穗粒数和千粒重, 提高产量(表1)。如2012年和2013年生长季BR处理玉米秃尖长分别比对照减小32.8%和28.9%, 而穗粒数分别比对照增加4.1%和5.8%。BR处理显著提高了2012年春玉米季千粒重, 但在2013年处理和对照间千粒重差异不显著。在两个生长季中, BR处理显著提高了春玉米产量, 分别比对照提高了9.8%和7.3%。
| 表1 BR对玉米产量构成因素与产量的影响 Table 1 Effect of BR on yield components and yield in maize |
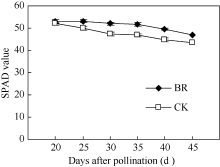
2.2.1 BR对穗位叶叶绿素含量的调控 BR处理对玉米穗位叶叶绿素含量影响显著, 如图2所示, 授粉后20~45 d, BR与CK穗位叶SPAD值均呈下降趋势, 但相同时期下BR处理显著高于CK, 且BR处理SPAD值下降速度低于CK。授粉后25 d BR处理穗位叶SPAD值与授粉后20 d基本持平, 而CK穗位叶SPAD值比20 d低4.2%。BR处理授粉后30 d、35 d、40 d和45 d分别比授粉后20 d下降1.9%、2.3%、6.4%和11.4%; 而CK则分别下降8.9%、10.0%、14.0%和16.1%。
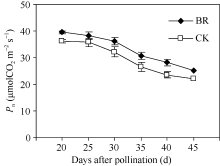
2.2.2 BR对穗位叶光合速率的调控 BR对玉米穗位叶光合速率的调控趋势与叶绿素含量相似, 也表现为显著提高效应。授粉后20~45 d, BR处理与CK的穗位叶光合速率均呈下降趋势。但授粉后20 d、25 d、30 d、35 d、40 d和45 d的BR处理穗位叶光合速率分别比CK高9.1%、7.0%、12.8%、15.4%、19.5%和13.6% (图3)。
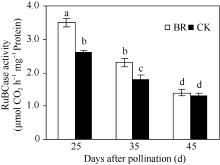
2.2.3 BR对穗位叶PEPCase和RuBPCase活性的调控 PEPCase和RuBPCase是玉米碳同化过程的关键酶。在授粉后25~45 d, 玉米穗位叶PEPCase (图4)和RuBPCase (图5)活性均呈下降趋势, 但BR处理显著高于CK。授粉后25 d、35 d和45 d, 分别比CK提高了16.7%、23.2%、44.4%与34.6%、27.8%、7.7%。
 | 图2 BR处理对玉米穗位叶叶绿素含量的影响BR: 0.1 mg L-1油菜素内酯; CK: 对照。误差线为标准差。Fig. 2 Effect of BR on chlorophyll content in maize ear leafBR: 0.1 mg L-1 brassinolide; CK: control. Error bars represent standard deviations. |
 | 图3 BR处理对玉米穗位叶光合速率的影响BR: 0.1 mg L-1油菜素内酯; CK: 对照。误差线为标准差。Fig. 3 Effect of BR on photosynthetic rate in maize ear leafBR: 0.1 mg L-1 brassinolide; CK: control. Error bars represent standard deviations. |
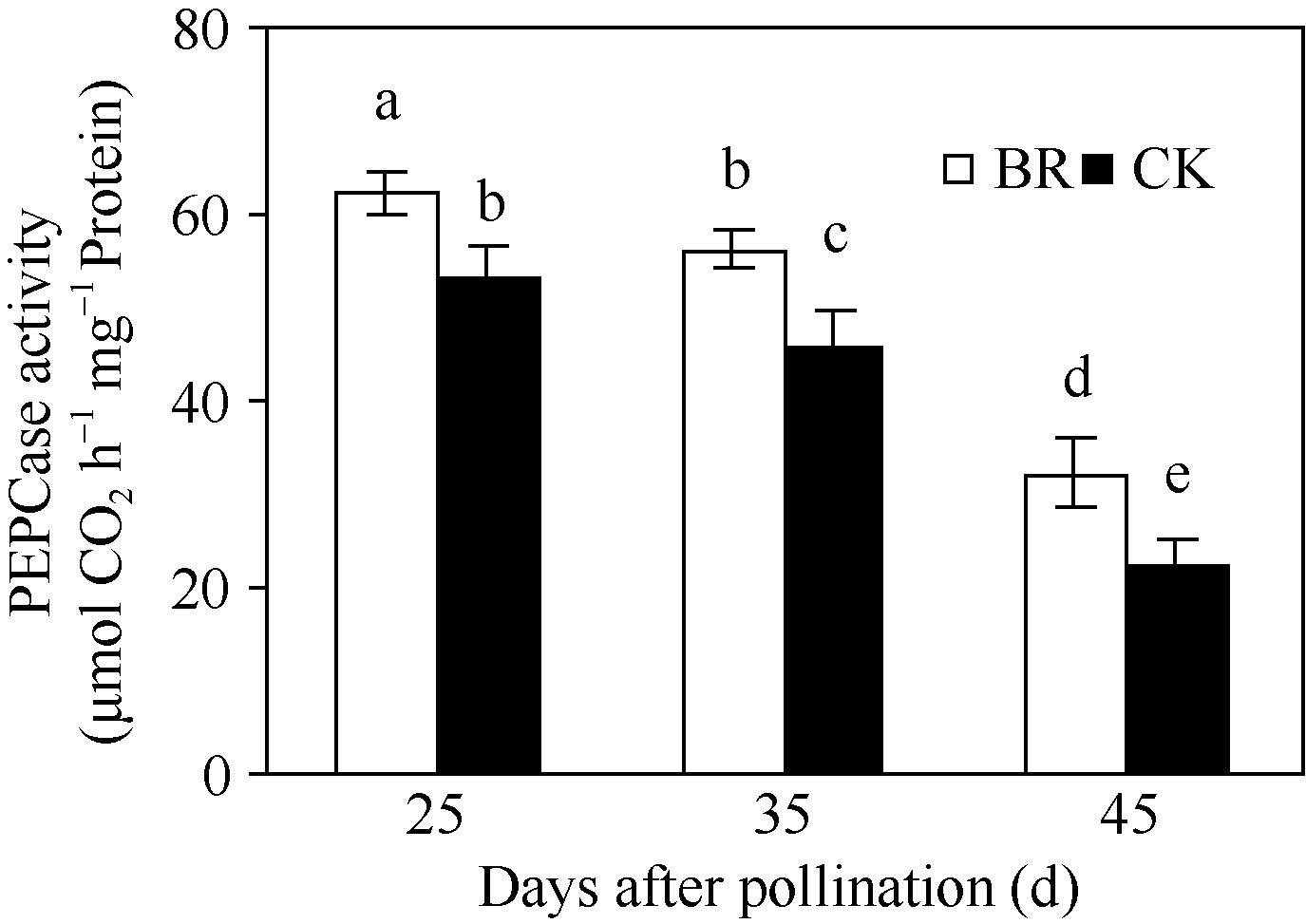 | 图4 BR处理对玉米穗位叶PEPCase活性的影响BR: 0.1 mg L-1油菜素内酯; CK: 对照。误差线为标准差; 不同字母表示0.05水平差异显著。Fig. 4 Effect of BR on activity of PEPCase in maize ear leafBR: 0.1 mg L-1 brassinolide; CK: control. Error bars represent standard deviations; different letters are significantly different at the 0.05 probability level. |
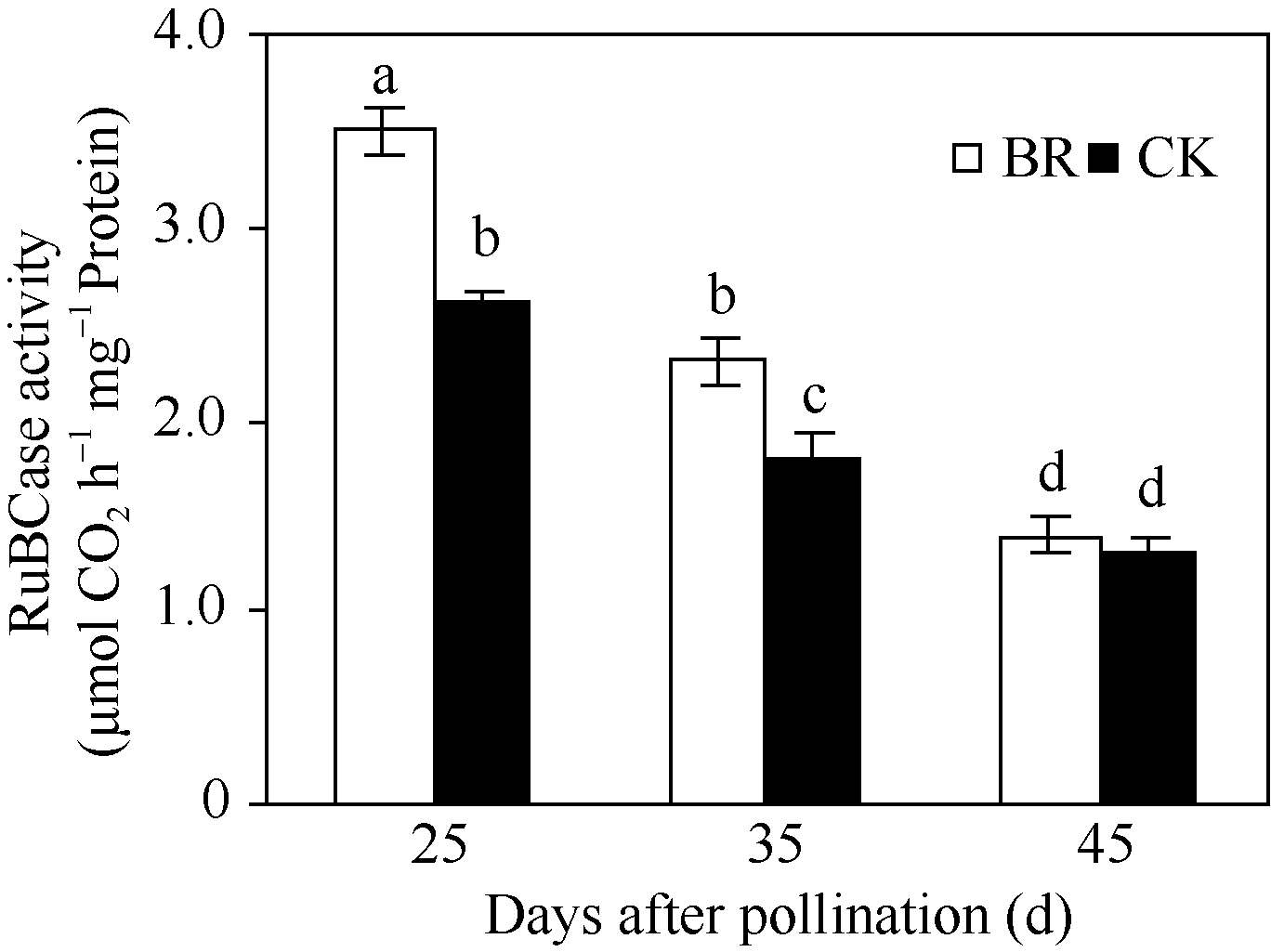 | 图5 BR处理对玉米穗位叶RuBPCase活性的影响BR: 0.1 mg L-1油菜素内酯; CK: 对照。误差线为标准差; 不同字母表示0.05水平差异显著。Fig. 5 Effect of BR on activity of RuBPCase in maize ear leafBR: 0.1 mg L-1 brassinolide; CK: control. Error bars represent standard deviations; different letters are significantly different at the 0.05 probability level. |
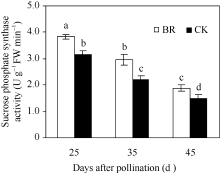
2.2.4 BR对玉米穗位叶蔗糖合酶和蔗糖磷酸合酶活性的调控 蔗糖是植物碳同化产物的主要运输形式, 而蔗糖合酶和蔗糖磷酸合酶是蔗糖合成的关键酶, 其活性是反映籽粒对同化物需求的重要指标。在授粉后25~45 d, 玉米穗位叶蔗糖合酶(图6)和蔗糖磷酸合酶(图7)活性均呈下降趋势, 但BR处理显著高于CK, 授粉后25 d、35 d和45 d分别比CK提高了25.0%、31.6%、15.2%和21.4%、34.2%和27.1%。
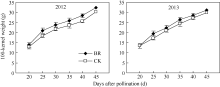
2.3.1 BR对玉米籽粒干物质积累的调控 2012年和2013年玉米灌浆过程中籽粒百粒干重的变化趋势如图8所示, 两季BR处理均显著提高了玉米籽粒干物质积累速率。吐丝后20~45 d, BR处理和CK百粒干重均呈上升趋势, 其中授粉后20~25 d BR处理籽粒百粒干重增加速率显著高于CK, 25~45 d两者干物质积累速率差异减小, 但BR处理百粒干重在各时期均显著高于CK。其中2012年和2013年授粉后45 d BR百粒干重分别比CK增加6.8%和3.6%。
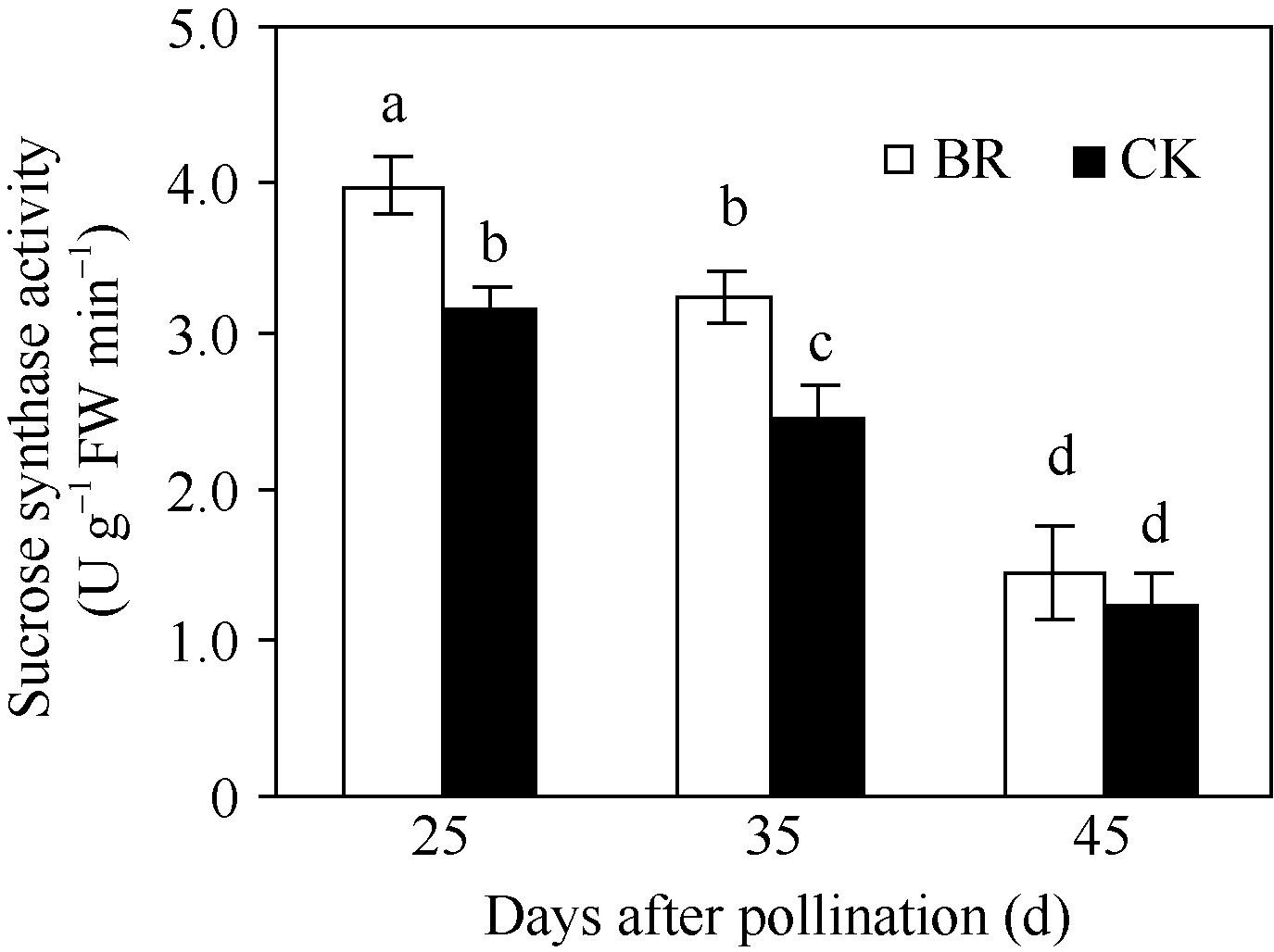 | 图6 BR处理对玉米籽粒蔗糖合成酶活性的调控BR: 0.1 mg L-1油菜素内酯; CK: 对照。误差线为标准差; 不同字母表示0.05水平差异显著。Fig. 6 Effect of BR on activity of sucrose synthase in maize kernelBR: 0.1 mg L-1 brassinolide; CK: control. Error bars represent standard deviations; different letters are significantly different at the 0.05 probability level. |
 | 图7 BR处理对玉米籽粒蔗糖磷酸合成酶活性的调控BR: 0.1 mg L-1油菜素内酯; CK: 对照。误差线为标准差; 不同字母表示0.05水平差异显著。Fig. 7 Effect of BR on activity of sucrose phosphate synthase in maize kernelBR: 0.1 mg L-1 brassinolide; CK: control. Error bars represent standard deviations; different letters are significantly different at the 0.05 probability level. |
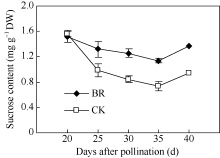
2.3.2 BR对玉米籽粒蔗糖含量的调控 授粉后20~35 d, 玉米籽粒蔗糖含量随时间呈下降趋势。在授粉后20 d, BR与CK间籽粒蔗糖含量无显著差异, 但在授粉后25 d、30 d、35 d和40 d, BR处理籽粒蔗
糖含量分别比CK高34%、48%、55%和45% (图9)。
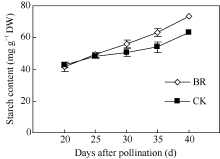
2.3.3 BR对玉米籽粒淀粉含量的调控 淀粉是玉米籽粒的主要组成成分, 占成熟籽粒干重的70%左右, 籽粒灌浆过程主要是淀粉合成和积累的过程。授粉后20~40 d玉米籽粒淀粉含量随时间呈上升趋势(图10)。授粉后20~25 d, BR和CK处理间玉米籽粒淀粉含量无显著差异; 但在授粉后30 d、35 d和40 d BR处理下玉米籽粒淀粉含量分别比CK高10.7%、17.0%和16.8%。
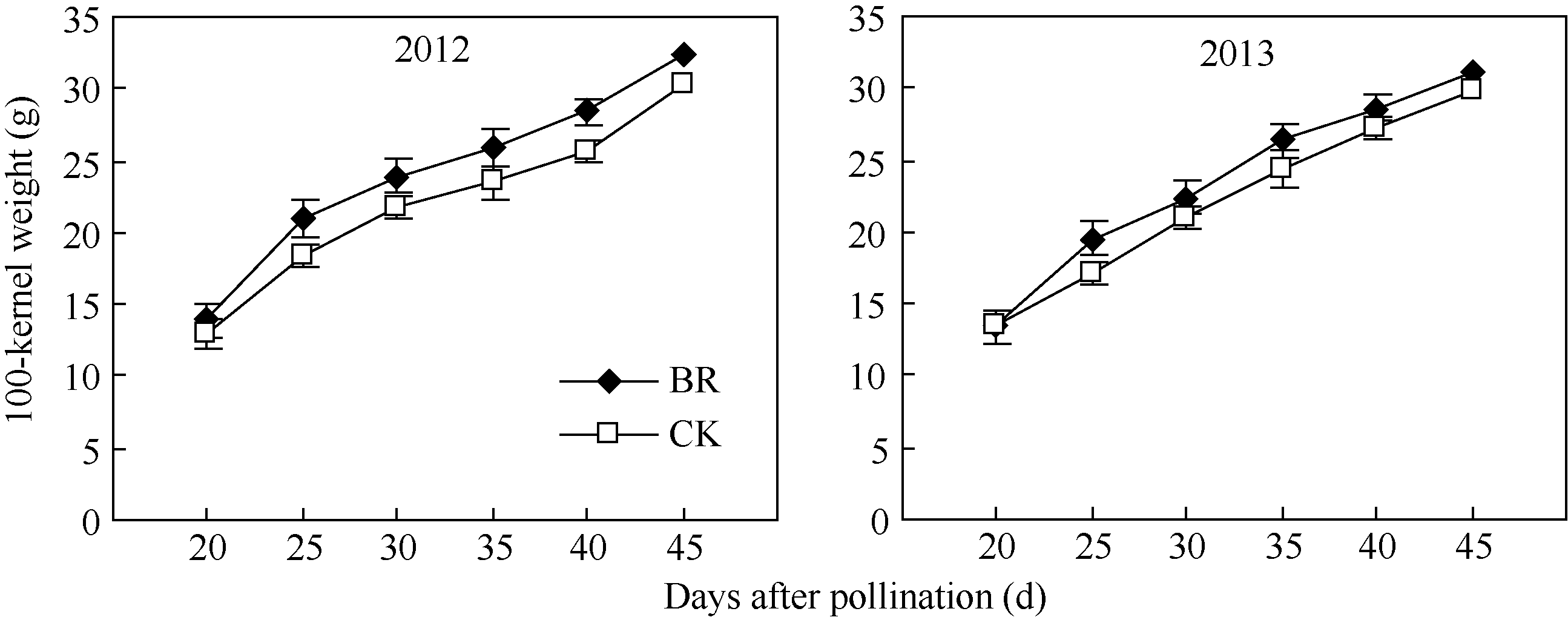 | 图8 BR处理对玉米百粒干重的调控BR: 0.1 mg L-1油菜素内酯; CK: 对照。误差线为标准差。Fig. 8 Effect of BR on dry weight of 100-kernel in maizeBR: 0.1 mg L-1 brassinolide; CK: control. Error bars represent standard deviations. |
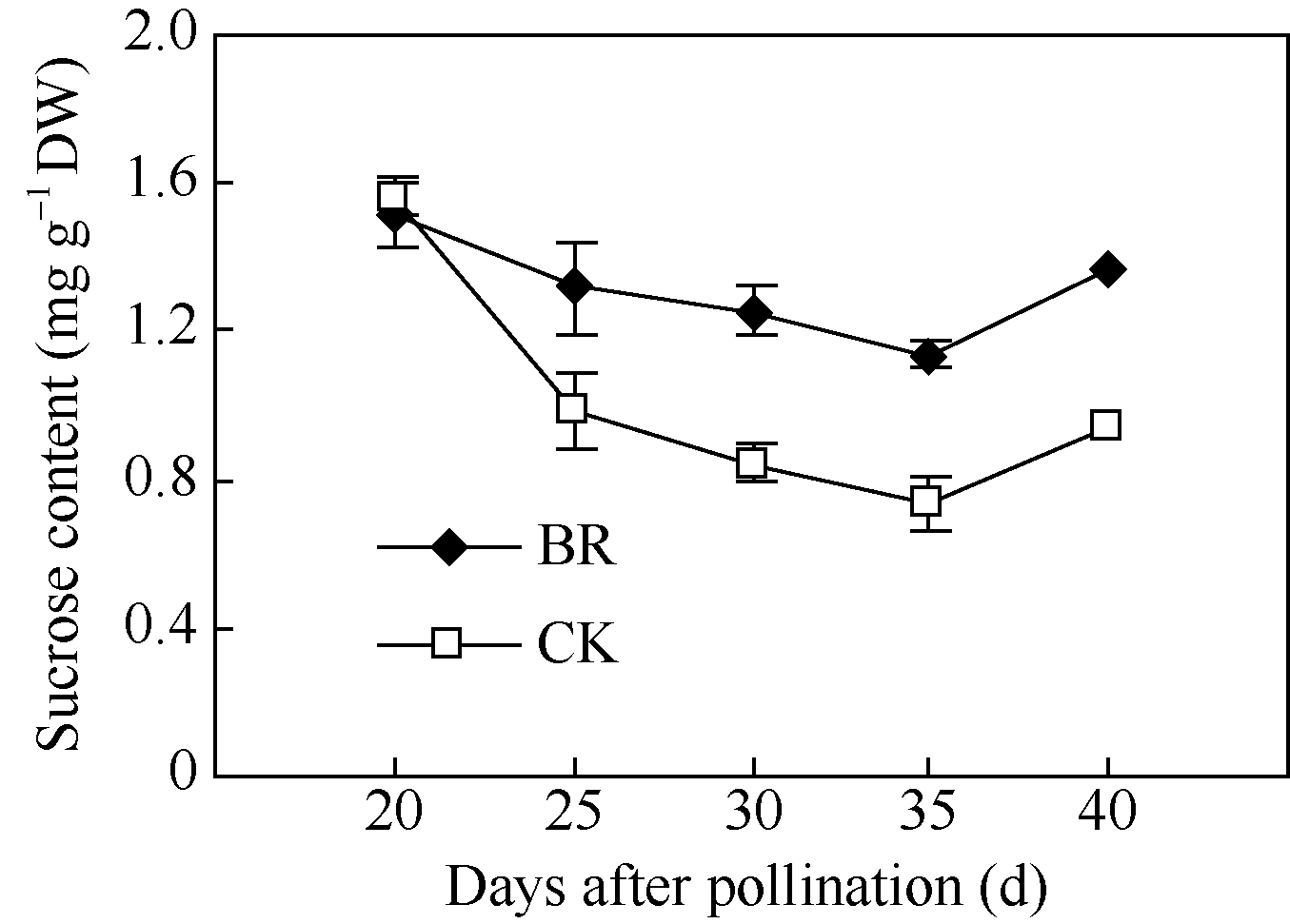 | 图9 BR处理对玉米籽粒蔗糖含量的影响BR: 0.1 mg L-1油菜素内酯; CK: 对照。误差线为标准差。Fig. 9 Effect of BR on sucrose content in maize kernelBR: 0.1 mg L-1 brassinolide; CK: control. Error bars represent standard deviations. |
籽粒灌浆依靠叶片等源器官制造的光合产物以蔗糖形式转运到籽粒, 在一系列酶的催化下形成淀粉[19]。穗位叶是玉米籽粒灌浆过程中最重要的源器官, 因此提高穗位叶光合效率, 增强碳同化作用, 促进光合产物向籽粒转运对玉米增产具有重要意义。本研究表明BR可以提高玉米穗位叶叶绿素含量和光合速率, 增强叶片PEPCase和RuBPCase的活性, 促进碳同化作用, 同时, BR显著提高了蔗糖磷酸合酶和蔗糖合酶活性, 促进了蔗糖合成, 有效提高了叶片源活性。这与前人在水稻上的研究相似, 如肖琳等[20]发现初花期喷施BR可显著增强水稻叶片的生理代谢, 提高剑叶叶绿素、可溶性糖和淀粉含量, 增强光合速率。Wu等[21]研究发现BR促进水稻光合同化物从源向库的转运。另外, 王士银等[22]研究发现BR提高水稻比叶重。
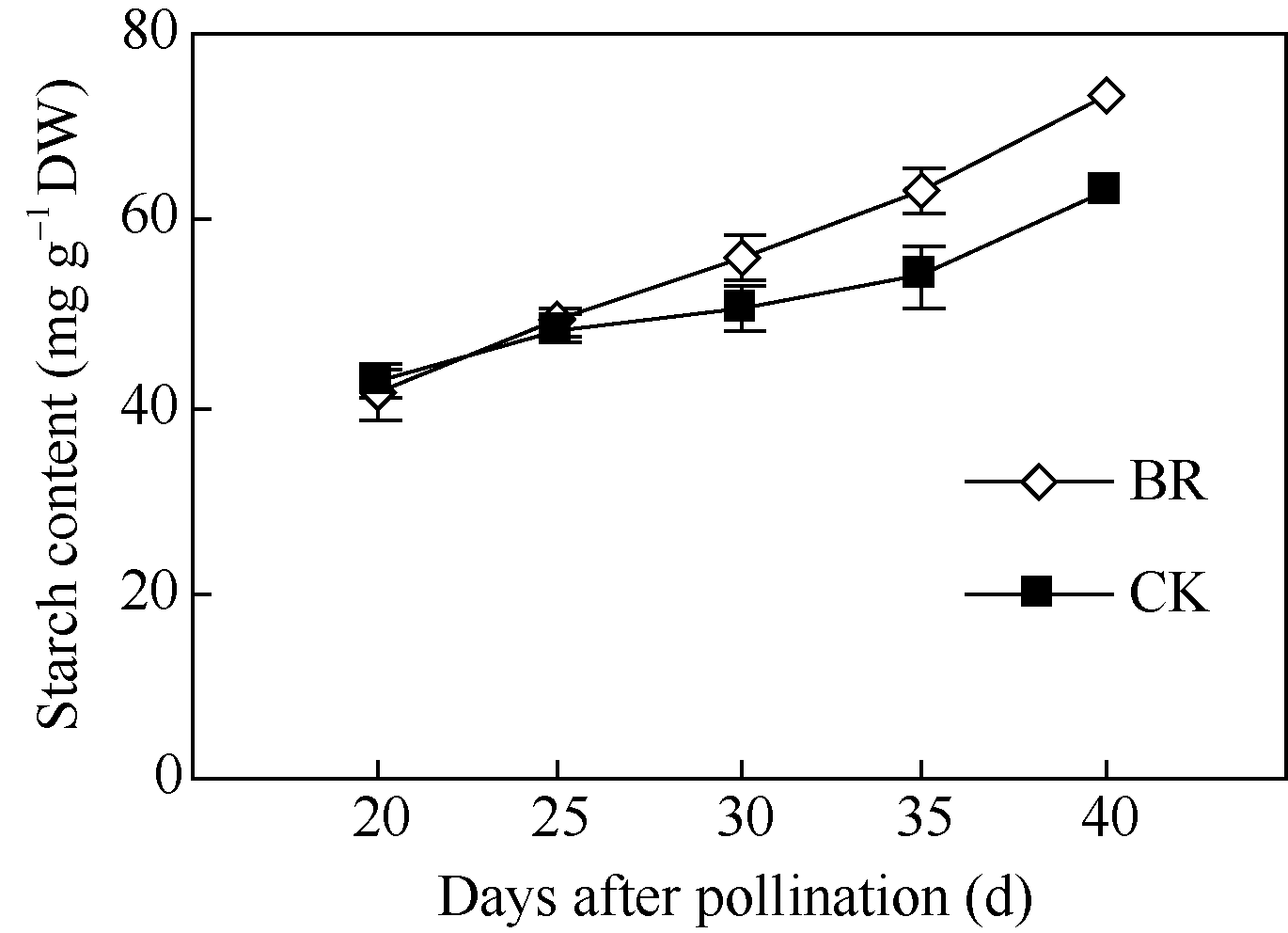 | 图10 BR处理对玉米籽粒淀粉含量的调控BR: 0.1 mg L-1油菜素内酯; CK: 对照。误差线为标准差。Fig. 10 Effect of BR on starch content in maize kernelBR: 0.1 mg L-1 brassinolide; CK: control. Error bars represent standard deviations. |
籽粒灌浆是与玉米产量及品质形成最密切的生理过程。本研究结果表明, 玉米籽粒灌浆过程中蔗糖含量逐渐下降, 淀粉含量逐渐上升。BR处理可显著提高授粉后25~40 d籽粒蔗糖含量, 加快淀粉积累, 有效提高淀粉含量和籽粒灌浆速率。前人在水稻BR缺陷或不敏感突变体上研究表明, BR在籽粒的形成过程中起重要作用[10, 23]。花期喷施表油菜素内酯可以提高小麦籽粒中ADPG焦磷酸化酶、可溶性淀粉合酶和淀粉分支酶的活性, 调控小麦籽粒灌浆期淀粉积累速率, 改善小麦籽粒特性等[24]。
BR在玉米穗位叶源活性和籽粒灌浆过程的调控反应在最终产量及产量构成因素上表现为秃尖的缩短, 穗粒数的增加, 粒重的增大, 以及最终产量的增加, 其中对秃尖长的调控最为明显。众多研究表明, BR对模式植物拟南芥籽粒体积增大, 粒重增加及粒数提高均有促进作用[9, 25, 26], 类似结果在水稻和小麦等作物的研究上也得到验证[20, 21, 22, 24]。
BR处理可以显著提高穗位叶叶绿素含量和光合效率, 增强叶片PEPCase和RuBPCase以及蔗糖磷酸合酶和蔗糖合酶的活性, 从而提高叶片源活性。BR处理促进籽粒淀粉合成和籽粒灌浆, 延长籽粒灌浆期, 最终增加粒重。BR处理显著缩短玉米秃尖长度, 增加穗粒数, 从而提高产量。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|