

{kind=link}
{kind=link}
{kind=link}
{kind=link}
小麦粒重基因 TaGW2-6A等位变异的组成分析及育种选择
[寇程 , 高欣, 李立群, 李扬, 王中华, 李学军
, 高欣, 李立群, 李扬, 王中华, 李学军* ]
, 高欣, 李立群, 李扬, 王中华, 李学军]
|
|


第一作者联系方式: E-mail:koucheng7417@163.com
位于6A染色体的 TaGW2是控制小麦籽粒大小的关键基因, 已发现其第8外显子有一个T碱基插入等位变异, 其启动子区存在Hap-6A-A及Hap-6A-G等位变异。利用高分辨率熔解曲线分析技术(high resolution melting curve analysis, HRM)和 Hap-6A-P1/P2分子标记检测了316份小麦品种(系)的 TaGW2-6A基因在上述2个位点的等位变异, 分析了其不同等位变异与粒长、粒宽和千粒重的相关性, 并以大面积推广的大粒品种周麦22为例, 解析 TaGW2-6A基因优异等位变异在系谱选育中的遗传传递。共检测到61份T碱基插入等位变异(命名为977T基因型)和255份无T碱基插入的等位变异(977-基因型)材料; 在977T基因型中, Hap-6A-A (TA)和Hap-6A-G (TG)单倍型材料分别为29份和32份, 在977-基因型中, Hap-6A-A (-A)和Hap-6A-G (-G)单倍型材料分别为160份和95份。关联分析表明, 977T基因型与977-基因型的粒长( P<0.05)、粒宽( P<0.001)和千粒重( P<0.001)均有显著差异, Hap-6A-A单倍型与Hap-6A-G单倍型的粒长( P<0.05)、粒宽( P<0.05)和千粒重( P<0.001)也有显著差异。 TaGW2-6A基因编码区和启动子区等位变异之间存在相互作用, 共同调控小麦籽粒的大小, 其中TA单倍型比TG、-A、-G单倍型更能增加小麦的粒宽和粒重, 是优异的等位变异组合。周麦22为TA单倍型, 系谱分析表明, 该等位变异并非来源于亲本周8425B, 而是来源于亲本辉县红, 且TA单倍型能够稳定遗传, 但是在常规育种选择过程中可能会丢失。本研究筛选出的 TaGW2-6A优异等位变异TA单倍型材料及高通量分子检测方法为分子标记辅助育种提供材料和方法依据。
TaGW2 on chromosome 6A is a key gene governing kernel size of wheat ( Triticum aestivumL.). There are a single T-base insertion in the eighth exon of TaGW2 (977 bp) and two SNPs (Hap-6A-A and Hap-6A-G) in the promoter region. In this study, 316 wheat varieties (lines) were detected by high resolution melting curve (HRM) analysis and Hap-6A-P1/P2 molecular marker for TaGW2-6A allelic variations and their correlations with kernel length, kernel width, and thousand-kernel weight were analyzed. Furthermore, the TaGW2-6Aallelic variations were traced in the pedigree of the famous large-kernel variety Zhoumai 22. In the 977 bp position, 61 and 255 lines were detected with and without the T-base insertion, which were designated 977T and 977-genotypes, respectively. In the 977T genotypes, 29 lines were Hap-6A-A (TA) haplotype and 32 lines were Hap-6A-G (TG) haplotype. In the 977-genotypes, 160 lines were Hap-6A-A (-A) haplotype and 95 lines were Hap-6A-G (-G) haplotype. Significant difference was found in kernel length ( P < 0.05), kernel width ( P < 0.001) and thousand-kernel weight ( P < 0.001) between 977T and 977-genotypes. Similarly, significant difference was also found in kernel length ( P < 0.05), kernel width ( P < 0.05) and thousand-kernel weight ( P < 0.001) between Hap-6A-A and Hap-6A-G haplotypes. The allelic variation in TaGW2-6A encoding region and the promoter region jointly contributed to kernel size, and the TA haplotype was superior to -A, TG, and -G haplotypes in increasing kernel width and weight. According to pedigree analysis, the Zhoumai 22 inherited the TA haplotype from the parent Huixianhong, not from the popular parent Zhou 8425B. This haplotype is inheritable stably but tends to be lost in the process of wheat breeding. The results of this study provide not only a high-throughput molecular technique to detect wheat TA haplotype but also breeding materials in marker-assisted selection of wheat.
小麦是全球主要的粮食作物, 占世界总耕地面积的四分之一[1]。小麦产量由单位面积穗数、穗粒数和千粒重三因素构成, 其中千粒重在产量构成因素中受遗传特性的影响最大, 广义遗传力高达59%~80%[2]。研究表明, 千粒重和穗粒数的增加对陕西关中地区小麦品种产量的提高有较大贡献, 其中千粒重的作用更为突出[3]。胡延吉等[4]提出, 随着品种产量水平的提高, 单株穗数或单位面积穗数一般没有相应增加或略有减少, 穗粒重成为进一步提高品种产量水平的主导因素。Wu等[5]研究表明, 小麦粒重的显著增加对1945年到2010年间中国多个小麦主产区产量的持续增长有较大贡献。可见, 在品种选育过程中注重对千粒重的选择是提高小麦产量的有效方法。
GW2基因是对作物粒重有较大贡献的一个功能基因。在水稻中, OsGW2基因编码一个新型E3泛素连接酶, 通过26S蛋白酶体系统负调控种子细胞的分裂[6]。在拟南芥中, 转AtGW2基因拟南芥籽粒千粒重明显高于野生型[7]。在玉米中, 连锁不平衡、关联分析及基因表达分析均表明ZmGW2基因控制玉米粒重[8]。在小麦中, Hong等[9]通过构建TaGW2RNAi表达载体并转化到小麦品种石4185中, 发现转化后石4185籽粒的粒宽、粒厚和粒重都显著增加, 证明TaGW2基因负调控小麦籽粒大小; Yang等[10]通过同源克隆得到小麦的粒重基因TaGW2, 发现兰考大粒中TaGW2-6A基因第8外显子977 bp处有一“ T” 碱基的插入, 关联分析表明该等位变异对小麦粒重、粒宽有显著影响; Su等[11]克隆了TaGW2-6A基因的启动子区, 序列比对分析发现存在Hap-6A-A (-593A和-739G)和Hap-6A-G (-593G和-739A)两种等位变异型, 相关性分析表明Hap-6A-A是小麦大粒的优异等位变异。
综上, TaGW2-6A基因的编码区“ T” 插入等位变异与启动子区Hap-6A-A和Hap-6A-G等位变异均对小麦种子大小有重要影响。目前, 关于该基因编码区和启动子区等位变异的相互作用及两个区域优异等位变异基因型系谱分析的相关研究还未见报道。
本研究以316份小麦品种(系)构成的自然群体为材料, 利用HRM分析技术和Hap-6A-P1/P2分子标记检测TaGW2-6A基因编码区和启动子区的等位变异, 研究4种等位变异与小麦粒长、粒宽和粒重的关系, 同时以大面积推广的大粒品种周麦22为例分析了TaGW2-6A基因优异等位变异在品种系谱选育中的遗传传递。旨在为粒重基因TaGW2-6A优异等位变异的分子标记辅助选择育种提供材料和方法依据。
由108份我国小麦微核心种质、174份黄淮麦区育成的小麦品种(系)及34个具有代表性的国外品种, 共计316个小麦品种(系)构成的自然群体为研究材料。在2012— 2014连续2年在西北农林科技大学北校区小麦育种试验田种植, 田间顺序排列, 人工播种; 每个品种种植1行, 行长2 m, 行距0.23 m, 田间管理同大田。
在小麦收获晾干后, 从每份材料随机取30粒完整饱满的籽粒, 按其长度方向紧密排成一行测定其长度, 再按籽粒宽度方向放置测定其宽度, 然后换算成单个籽粒的粒长和粒宽(mm), 每份材料重复测量3次, 取平均值。随机取200粒种子称重, 每份材料2次重复, 换算为千粒重(g)。
用SPSS 19.0和Microsoft Excel 2007计算和统计分析调查数据, 用LSR (least significant ranges)方法比较基因型间的差异显著性。
每个品种取0.2~0.3 g新鲜叶片, 液氮预冷后粉碎放入1.5 mL离心管, 按照Gawel等[12]的方法提取小麦基因组DNA。
1.2.1 TaGW2-6A基因编码区“ T” 碱基插入等位变异
根据兰考大粒TaGW2-6A基因T插入位点附近序列设计引物HRM-P (F: 5′ -ATGGGTGCTGCGGA AAGT-3′ ; R: 5′ -CGCTCCAGCTATCTGGTGAA-3′ ), 扩增片段长度为38 bp。引物由宝生物工程(大连)有限公司合成, 纯化级别为PAGE。用Light Cycler 480 (Roche)进行PCR扩增及HRM分析, 反应体系为20 μ L, 含80 ng模板DNA, 上下游引物各0.4 µ mol L-1, 10 μ L Light Cycler HRM Master Mix (Roche), 1.5 mol L-1 Mg2+ (Roche); 反应程序, 首先进行普通PCR (95℃ 5 min; 95℃ 30 s, 58℃ 30 s, 72℃ 2 s, 40个循环; 72℃ 10 min), 再对其产物进行HRM分析, 即以95℃ 30 s使荧光染料结合到碱基上, 再降温至59℃ 30 s使解开的DNA单链重新结合成双链结构, 随后逐渐升温到95℃, 每升高1℃收集25次荧光信号, 最后迅速降到37℃, 保持10 s, 完成整个HRM分析反应。
1.2.2 HRM-P引物扩增产物序列分析 中国春和兰考大粒的HRM反应产物经3%琼脂糖凝胶电泳分离, 将目标片段挖胶回收, 用pUCm-T载体克隆, 由生工生物工程(上海)股份有限公司测序。每个品种至少测定10个克隆。采用DNAMAN软件分析引物的扩增产物序列等位变异, 并在NCBI数据库(http://www.ncbi.nlm.nih.gov/)上进行比对。
1.2.3 TaGW2-6A基因启动子区等位变异 利用Su等[12]设计的Hap-6A-P1/P2分子标记检测316份供试材料中TaGW2-6A基因启动子区Hap-6A-A和Hap-6A-G等位变异。
以同时含有两种优异等位变异(编码区为T插入型; 启动子区为Hap-6A-A单倍型)的大粒品种周麦22为例, 通过HRM分析技术及Hap-6A-P1/P2分子标记检测周麦22系谱中各亲本TaGW2-6A基因编码区及启动子区等位变异的组成, 进而分析TaGW2- 6A基因T碱基插入等位变异以及启动子区Hap-6A- A和Hap-6A-G等位变异的遗传传递规律。周麦22于2007年通过国家审定, 截至2014年夏收, 在黄淮麦区累计推广面积460多万公顷, 是大粒高产品种的代表。
HRM分析结果显示HRM-P引物能够明显的区分出兰考大粒与中国春高分辨率熔解曲线的差异。目的片段扩增产物测序显示, 中国春扩增产物只有一种序列, 而兰考大粒扩增产物存在两种序列, 一种序列与中国春的序列完全相同, 推测其为6B、6D基因组TaGW2扩增的产物; 另一种序列比中国春多一个T碱基插入(图1), 将其在NCBI数据库进行比对后发现与TaGW2-6A-LK目的序列完全相同, 由此判断HRM分析技术显示出的熔解曲线的差异是由TaGW2-6A基因T碱基插入引起的。
 | 图1 HRM-P引物在中国春和兰考大粒中扩增产物的序列比对Fig. 1 Sequence alignment of PCR products in Chinese Spring and Lankaodali amplified with HRM-P primer |
316份材料中有61份材料的熔解曲线呈现与兰考大粒的熔解曲线相近, 255份材料的熔解曲线呈现与中国春的熔解曲线相近(图2), 由此得出供试材料中T碱基插入材料为61份(977T基因型), 无T碱基插入的材料为255份(977-基因型)。
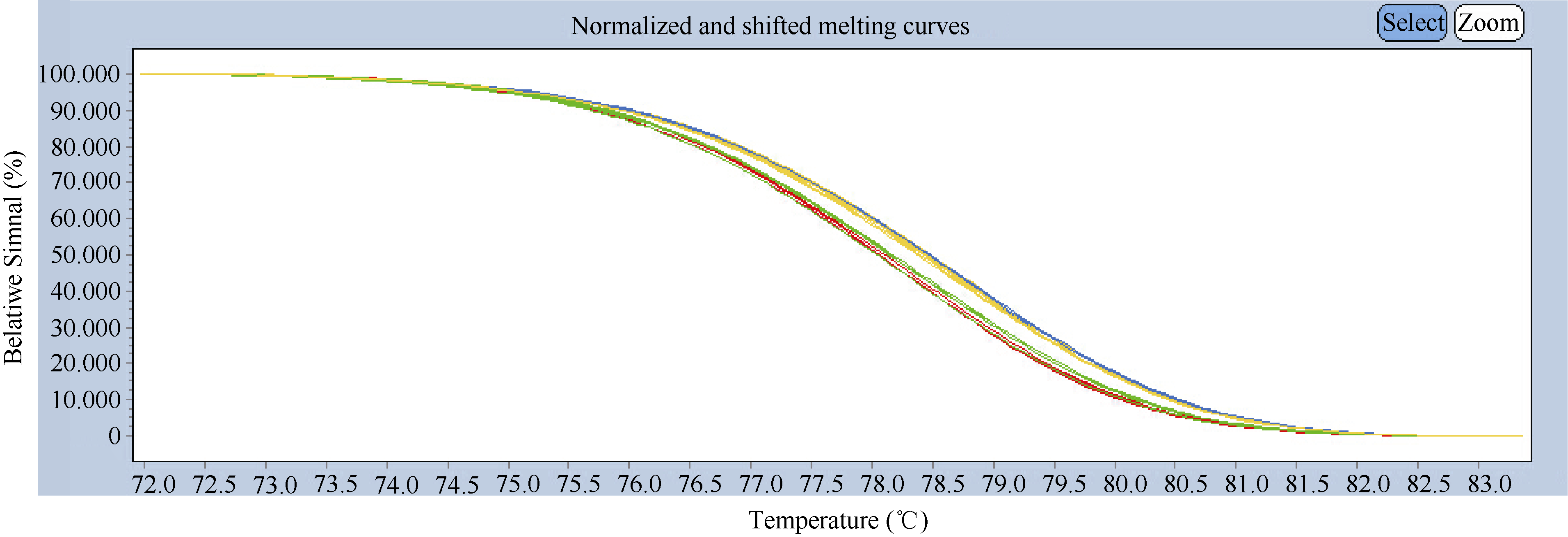 | 图2 977T和977-基因型的高分辨率熔解曲线分析红色、蓝色、绿色、黄色曲线分别表示兰考大粒、中国春、977T基因型和977-基因型熔解曲线。Fig. 2 High-resolution melting curve analysis of 977T and 977-genotypesThe melting curves of Lankaodali, Chinese Spring, 977T genotype, and 977-genotype are in red, blue, green, and yellow, respectively. |
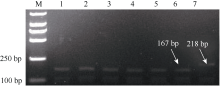
用Hap-6A-P1/P2引物进行PCR扩增, 产物在1%琼脂糖凝胶上电泳检测, 区分启动子区Hap-6A-A (167 bp)和Hap-6A-G (218 bp)两种等位变异材料(图3), 结果Hap-6A-A等位变异材料189份, Hap-6A-G等位变异材料127份。
 | 图3 TaGW2-6A基因启动子区的多态性分析Hap-6A-A和Hap-6A-G等位变异的扩增片段长度分别为167 bp和218 bp。M: DL2000 marker; 1: 周麦22; 2: 西农2208; 3: 陕229; 4: 蚂蚱麦; 5: 中麦349; 6: 小偃22; 7: 藁优9618。Fig. 3 Polymorphism analysis on promoter region of TaGW2-6AThe sizes of amplified fragments were 167 bp for genotype Hap-6A-A and 218 bp for genotype Hap-6A-G. M: DL2000 marker; 1: Zhoumai 22; 2: Xinong 2208; 3: Shaan 229; 4: Mazhamai; 5: Zhongmai 349; 6: Xiaoyan 22; 7: Gaoyou 9618. |
根据HRM和Hap-6A-P1/P2分子标记分析结果, 将316份材料分成TA、TG、-A和-G四种单倍型, 分别为29、32、160和95份。表1列出了主要品种的相关信息。
| 表1 部分重要品种名称及基因型表型数据 Table 1 Part of important varieties and their genotype-phenotype data |
供试材料粒重表型数据与TaGW2-6A等位变异关联分析结果表明, 977T基因型与977-基因型材料的粒长(P< 0.05)、粒宽(P< 0.001)和千粒重(P< 0.001)差异显著, 而Hap-6A-A与Hap-6A-G基因型之间粒长(P< 0.05)、粒宽(P< 0.05)和千粒重(P< 0.001)也有显著差异。TA单倍型与TG单倍型材料的粒长、粒宽和千粒重差异不显著, 其中TA单倍型与-A单倍型(P< 0.05)和-G单倍型(P< 0.001)存在显著差异; TG单倍型与-A单倍型的3个籽粒性状无显著差异, 但二者都与-G单倍型(P< 0.05)呈显著差异(表2)。可见, TA单倍型比TG、-A、-G单倍型更能增加小麦的粒宽和粒重, 是优异的等位变异组合。
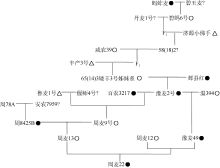
根据分子标记检测结果, 周麦22为TA单倍型, 而其系谱中相关亲本在TaGW2-6A基因编码区和启动子区的等位变异呈多样性(图4)。周麦22是以周麦12/豫麦49为母本、周麦13为父本杂交选育而成的, 其中周麦12和周麦13是-A单倍型材料; 周麦13又是由周麦骨干亲本周8425B和周麦9号杂交选育而成, 其中周8425B是TA单倍型; 豫麦49遗传了周麦22的1/4血统, 而豫麦49是TA单倍型。可见, 周麦22的TA单倍型并非来源于周麦13的亲本周8425B, 而是来源于豫麦49, 并通过系谱向上追溯到豫麦2号。豫麦2号的一个亲本65(14) 3是矮丰3号(-A单倍型)的姊妹系, 另一个亲本是辉县红(TA单倍型), 因此推定, 豫麦2号的TA单倍型来源于小麦地方品种辉县红。可见, TaGW2-6A基因的TA单倍型能够在品种选育过程中稳定遗传, 但如果选择不当也可能丢失, 如蚂蚱麦的TA单倍型没有传递到碧蚂6号, 周8425B的TA单倍型没有传递到周麦13 (图4)。
| 表2 TaGW2-6A基因等位变异的表型关联分析 Table 2 Correlation analysis of TaGW2-6A polymorphism and phenotype |
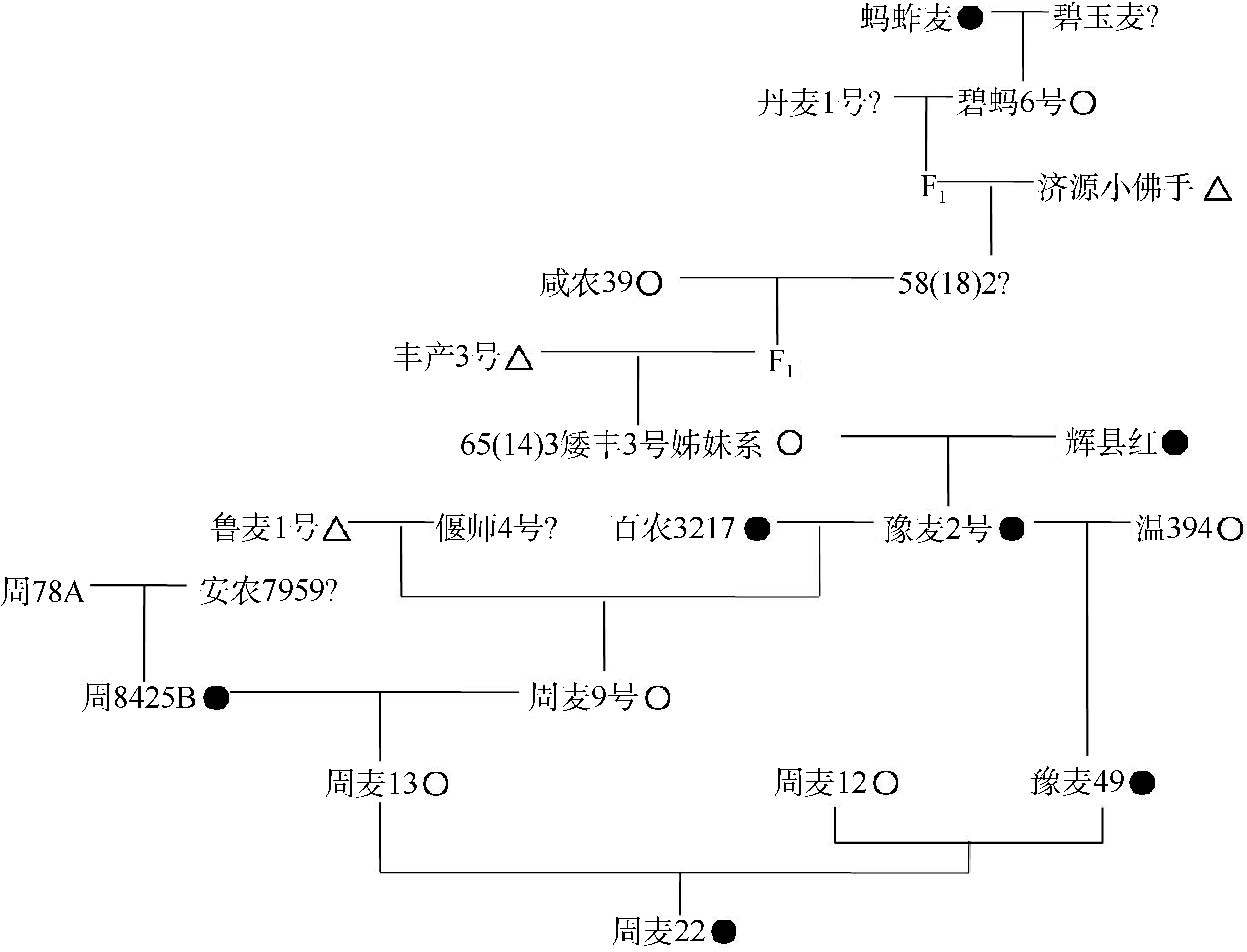 | 图4 周麦22系谱●: TA单倍型; ○: -A单倍型; △ : -G单倍型; ?: 没有进行分子检测。Fig. 4 Pedigree of Zhoumai 22●: TA genotype; ○: -A genotype; △ : -G genotype; ?: not detected. |
在TaGW2-6A基因T插入等位变异的品种筛选过程中, Yang等[10]利用AS-PCR技术来筛选T基因型材料, Liu等[13]针对特异等位位点设计了多对AS-PCR引物, 发现该类引物在扩增过程中无法达到稳定检测特异等位位点的目的。我们在利用AS-PCR技术检测的过程也发现其重复性差的问题。目前为止, 小麦粒重性状相关的分子标记被开发的较多, 但能够直接用于辅助育种的很少[14]。高分辨率熔解曲线分析(high resolution melting curve analysis, HRM)是Gundry等[15]于2003年提出并最初应用于SNP检测分析的一项新技术, 具有灵敏度高、特异性强、速度快、通量高、操作简便等优点, 近些年已经用于标记辅助选择、突变检测及种质鉴定等研究[16]。例如, 利用该技术, Ganopoulos等[17]分析菜豆SSR标记检测结果, 实现了菜豆品种的快速准确鉴定; Jeong等[18]利用COSII标记和Waxy基因的SNP标记数据, 对辣椒种质资源进行了鉴定; Han等[19]利用四倍体苜蓿的多个SNP位点数据, 对其亲本及后代群体进行SNP分型; Hofinger等[20]将HRM技术用于大麦eIF4E等位变异筛选。本研究将HRM技术引入小麦TaGW2-6A基因等位变异分析, 我们设计的HRM-P引物组合能有效区分两种基因型, 准确率达到100%且重复性良好, 为TaGW2-6A基因编码区优异等位变异的分子标记辅助选择提供了高通量的分子检测方法。
Yang等[10]以兰考大粒与中国春为亲本构建F2:3群体, 通过TaGW2-6A基因型与表型的关联分析, 证实该基因位点T插入突变与籽粒的表型相关, 能够显著增加粒宽和粒重。Su等[11]利用自然群体进行关联分析, 发现Hap-6A-A相对于Hap-6A-G是优异等位变异类型。本研究进一步表明, TaGW2-6A基因TA单倍型的粒宽和粒重大于TG、-A和-G单倍型, TG和-A单倍型的粒宽和粒重显著大于-G单倍型, 而TG单倍型与-A单倍型对粒宽、粒重的作用不显著, 说明TaGW2-6A基因TA单倍型是优异的等位变异组合。然而, 在977T基因型中也出现如藁优9618、中农28、定西24等粒重较小的材料, 其TaGW2-6A基因启动子区为Hap-6A-G等位变异; 同时, 在977-基因型中也存在周麦18、中麦895等籽粒较大的材料, 其TaGW2-6A基因启动子区为Hap-6A-A等位变异。根据本研究结果推测, TaGW2-6A基因编码区和启动子区等位变异之间存在相互作用, 共同调控TaGW2-6A基因表达, 从而控制小麦籽粒的大小和粒重。
本研究利用HRM分析技术以及Hap-6A-P1/P2分子标记, 在蚂蚱麦、西农6028、辉县红、周8425B、豫麦49、西农1376、豫麦2号、周麦22等黄淮麦区的骨干亲本中均检测到TA单倍型, 说明本研究采用的技术策略可用于小麦TaGW2-6A优异等位变异组合的检测和筛选。另一方面, 利用该方法我们进行了周麦22 TA单倍型的系谱追踪, 发现TA单倍型是从辉县红、豫麦2号、豫麦49传递到周麦22中, 而周麦22的其他亲本, 如蚂蚱麦、周8425B, 其TA单倍型没能传递给后代, 说明育种过程中如果选择不当, 将会导致优异基因丢失。本研究采用的分子标记检测技术和方法, 可有效监测小麦遗传改良中TaGW2-6A基因优异等位变异及其单倍型的传递过程, 提高传统育种的目标性和育种速度, 为加速培育突破性新品种提供技术支撑。
利用HRM分析技术和Hap-6A-P1/P2分子标记对316份小麦品种(系)进行了检测, 发现约一半品种(160份, 50.6%)为-A单倍型, 其次为-G单倍型(95份, 30.1%), TA和TG单倍型品种较少, 分别为29份(9.2%)和32份(10.1%), 尤其是优异等位变异组合TA单倍型的频率最低。TaGW2-6A基因编码区和启动子区等位变异之间存在相互作用, 共同调控TaGW2-6A基因从而控制小麦籽粒的大小。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|