
{kind=link}
{kind=link}
{kind=link}
{kind=link}
甘蓝型油菜 BnTT3基因的表达与eQTL定位分析
[卢坤1, 2, *;*  , 曲存民
, 曲存民1, 2, *;* , 李莎1, 2 , 赵会彦1, 2 , 王瑞1, 2 , 徐新福1, 2 , 梁颖1, 2 , 李加纳1, 2, * ]
, 曲存民, 李莎]
|
|


第一作者联系方式: E-mail:drlukun@swu.edu.cn (卢坤); lion4302@163.com (曲存民)
**同等贡献(Contributed equally to this work)
类黄酮途径中, TT3编码的4-二氢黄铜醇还原酶是参与原花色素和花青素合成的关键酶。为了明确该基因可能的上游调控网络, 利用黄籽母本GH06和黑籽父本ZY821构建的遗传图谱, 以 BnTT3基因在高世代重组自交系群体中随机选取的94个株系花后40 d种子的表达量作为性状, 采用复合区间作图法进行eQTL分析。结果共检测到5个表达量相关的eQTL, 分别位于A03、A08、A09和C01染色体, 单个eQTL解释表型变异的5.22%~24.05%。A09染色体上存在2个主效eQTL, 单个eQTL分别解释24.05%和16.55%的表型变异, 分别位于标记KS10260~KBrB019I24.15和B055B21-5~KS30880之间, 微效eQTL分布于A03、A08和C01染色体上。A09染色体上的2个主效eQTL区间(包含200 kb侧翼序列)与拟南芥、白菜、甘蓝和芸薹族近缘物种基因组同源区段具有很好的共线性关系。基因注释结果表明检测到的eQTL均为 trans-QTL, 2个主效eQTL区段共包含78个基因, 包括 MYB51、 MYB52和 bZIP5转录因子, 可能为 BnTT3基因的上游直接调控因子, 对这些基因功能的深入分析将有助于阐明甘蓝型油菜黄籽性状形成的分子调控机制, 为黄籽候选基因的克隆筛选奠定基础。
, QU Cun-Min, LI Sha
In flavonoid biosynthesis pathway, the key enzyme dihydroflavonol 4-reductase (DFR) involved in the proanthocyanidin and anthocyanin biosynthesis pathway is encoded by TRANSPARENT TESTA 3 ( TT3) gene. The objective of this research was to identify the upstream regulatory networks of BnTT3 using the composite interval mapping method (CIM). Hence, we performed an eQTL analysis for the transcript-level variation of BnTT3 gene in seeds at 40 days after flower (DAF) in 94 recombinant inbred lines (RILs) derived from a cross between the yellow-seeded female parent GH06 and black-seeded male parent ZY821. Five eQTLs for expression levels of BnTT3 were detected on four different chromosomes (A03, A08, A09, and C01) in B. napus, accounting for 5.22% to 24.05% of phenotypic variation. Two major eQTLs were found and located among the markers KS10260-KBrB019I24.15 and B055B21-5-KS30880 of chromosome A09, with explained 24.05% and 16.55% of phenotypic variation, respectively. Three minor eQTLs were also detected to be located on chromosomes A03, A08, and C01. The flanking sequences with 200 kb of two major eQTLs on chromosome A09 of B. napus showed well synteny to those of A. thaliana, Brassica rapa, Brassica oleracea and other Brassiceae relatives. Furthermore, the annotation results showed that they belong to the trans-QTL, containing 78 genes in the two trans-QTL regions. Some transcription factors ( MYB51, MYB52, and bZIP5) might be upstream regulatory factors associated with transcriptional regulation of BnTT3. Therefore, further study about these genes function will be helpful to elucidate the molecular mechanism of the seed coat colour formation, as well as lay the foundation for selecting candidate genes of seed coat colour in B. napus.
类黄酮是植物中一类重要的次生代谢产物, 在花、叶、果实和种子颜色变化过程中起着至关重要的作用, 也有些类黄酮物质可作为抗氧化剂, 并对人类健康有益而备受关注[1, 2, 3, 4]。目前的研究表明模式植物拟南芥(Arabidopsis thaliana)种子颜色不同程度变异的主要原因是类黄酮途径中一些关键酶基因的突变, 引起拟南芥体内类黄酮物质的积累或终产物的合成异常[5]。同为十字花科植物的甘蓝型油菜(Brassica napus)与拟南芥亲缘关系很近, 其基因组编码序列保守性达到86%, 因此, 两种植物的种皮色泽变化和色素合成机理有较多相似之处。尽管在芸薹属作物中, 研究证实种皮色泽与控制类黄酮合成相关基因存在一定关联性[6, 7, 8, 9, 10], 本课题组也克隆了甘蓝型油菜苯丙烷-类黄酮代谢途径中多个透明种皮(TRANSPARENT TESTA, TT)基因家族的基因序列, 其中一些基因在黄籽油菜种子发育过程中下调表达(F3'H、PAL1、TTG1和TT2)[11, 12, 13], 但目前的研究还主要集中于对单个基因功能的解析, 尚缺乏对这些基因上下游调控网络和调控机制的研究。
随着分子标记技术的发展, 构建的高密度分子标记连锁遗传图谱已有效地应用于数量性状基因定位, 图位克隆和分子标记辅助育种中。为了研究控制基因表达量变异的上游调控位点, Jansen和Nap[14]提出将全基因组中每个基因的表达量作为数量性状, 对其进行QTL定位分析, 即基因表达的数量性状定位分析技术(expression QTL, eQTL), 这种方法不仅有助于寻找控制这些基因转录表达模式的上游调控位点, 发掘受该基因调节的下游基因及与该基因协同作用的基因, 而且还有可能建立较为完整基因表达调控网络, 以阐明基因转录表达的调控机制, 从而在表达及调控水平上分析控制复杂性状的遗传基础[15]。目前, eQTL方法不仅成功地应用到酵母(Saccharomyces cerevisiae)、小鼠(Mus musculus)和人(Homo sapiens)的相关研究中, 而且在拟南芥、玉米(Zea mays)、小麦(Triticum aestivum)和大麦(Hordeum vulgare)等植物的研究中也取得了重要成果[16, 17, 18]。
在最近的研究中, 我们发现编码4-二氢黄酮醇还原酶(dihydroflavonol 4-reductase, DFR)的TT3基因在不同来源的黄黑籽甘蓝型油菜中存在着稳定性的序列变异[19]。在类黄酮途径中, TT3基因是参与原花色素和花青素合成的重要基因, 其拟南芥突变体种皮为黄色透明[1, 20]。因此, 本研究以典型黄籽GH06与黑籽中油821为亲本构建重组自交系群体, 采用qRT-PCR检测BnTT3基因在群体株系中的相对表达量, 以复合区间作图(composite interval mapping, CIM)法对BnTT3基因表达量进行eQTL分析, 期望通过紧密连锁分子标记确定参与BnTT3基因表达调节的上游调控区段, 明确BnTT3基因的上游直接调控因子, 研究结果将有助于理解甘蓝型油菜黄籽性状形成的分子调控机制, 为黄籽主效候选基因的筛选克隆提供依据。
以典型黄籽与黑籽遗传背景的GH06和中油821为亲本, 通过“ 一粒传法” 连续自交10代建成高世代重组自交系群体。2011— 2012年在重庆市北碚歇马镇试验基地种植该群体, 9月20日播种育苗, 10月30日移栽至大田, 无重复, 随机排列, 每小区3行, 每行15株, 田间按常规生产方式管理。任选94个株系, 在油菜盛花期选6株正常生长的植株标记主花序开花时间, 在花后40 d, 取主花序天然授粉结实的种子, 以家系为混合单位, 用液氮冷冻后, 于-80℃超低温冰箱保存备用。
以植物总RNA提取试剂盒(天根生化科技有限公司, 北京)提取各株系总RNA, 用DNase I (宝生生物工程有限公司, 大连)消化RNA样品中痕量基因组DNA, 然后各取1 μ g总RNA, 以Oligo dT-Adaptor Primer进行反转录[TaKaRa RNA PCR Kit (AMV) Ver.3.0], 合成总cDNA第1链, 并参照Qu等[21]的方法, 用Bn26S检测反转录效率。
采用qRT-PCR方法检测BnTT3基因在亲本GH06和ZY821及后代株系花后40 d种子中的相对表达量。扩增引物为BnTT3F (5′ -TTCCACATAGCAACTCCCATG-3′ )和BnTT3R (5′ -TTATCCCCAACACTCCATTCAC-3′ ), 扩增反应体系含0.5 μ L (约100 ng)模板cDNA、10 μ L SYBR Premix Ex TaqII (2× )、10 μ mol· L-1上下游引物各0.8 μ L, 加ddH2O至20 μ L。用实时荧光定量PCR仪Bio-Rad CFX96进行扩增反应, 程序为95℃预变性2 min, 40个循环(95℃变性10 s, 60℃退火20 s), 扩增结束后, 进行熔解曲线分析。用Bio-Rad CFX Manager记录分析qRT-PCR结果, 由软件自动设定阈值线。根据参照基因ACT7 (5′ -T GGGTTTGCTGGTGACGAT/TGCCTAGGACGACCAACAATACT-3′ ); UBC21 (5′ -CCTCTGCAGCCTCCTCAAGT/CA TATCTCCCCTGTCTTGAAATGC-3′ )的2-Δ Δ CT法计算目的基因相对表达量[22], 3次重复。通过SPSS 13.0软件对数据进行t检验。
由重庆市油菜工程技术研究中心构建的遗传图谱含19个连锁群, 包括1089个标记位点(451个SRAP标记, 456个SSR标记, 97个RAPD标记和75个IBP标记), 覆盖基因组长度约为2775 cM, 标记间的平均距离为2.54 cM。
采用QTL分析软件Windows QTL Cartographer 2.5及复合区间作图(composite interval mapping, CIM)法, 将甘蓝型油菜BnTT3基因相对表达量作为考察性状, 参照曲存民等[23, 24]的方法, 根据BnTT3在各株系中的表达量进行eQTL定位和效应检测[25]。
通过克隆获得的BnTT3基因序列进行BlastN本地比对, 根据比对结果确定基因在染色体上的位置, 并提取eQTL连锁标记上下游各200 kb的侧翼序列进行基因注释, 如果在此区间内存在BnTT3基因则该eQTL为cis-QTL, 表明基因自身定位于此; 如果此区段未发现该基因, 则该eQTL为trans-QTL, 表明该eQTL可能为该基因上游调控基因的所在位点。在Brassica database (BRAD, http:// brassicadb.org/brad/)和Genoscope (http://www.genoscope. cns.fr/brassicanapus/)网站进行甘蓝型油菜中eQTL区段与近缘或亲本物种的共线性分析。
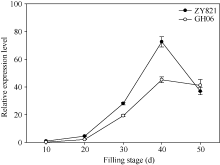
qRT-PCR检测结果表明, BnTT3基因在甘蓝型油菜亲本中的表达模式类似, 其表达量均在花后40 d达到峰值, 且在黑籽亲本ZY821中的表达量显著高于黄籽亲本GH06 (图1)。
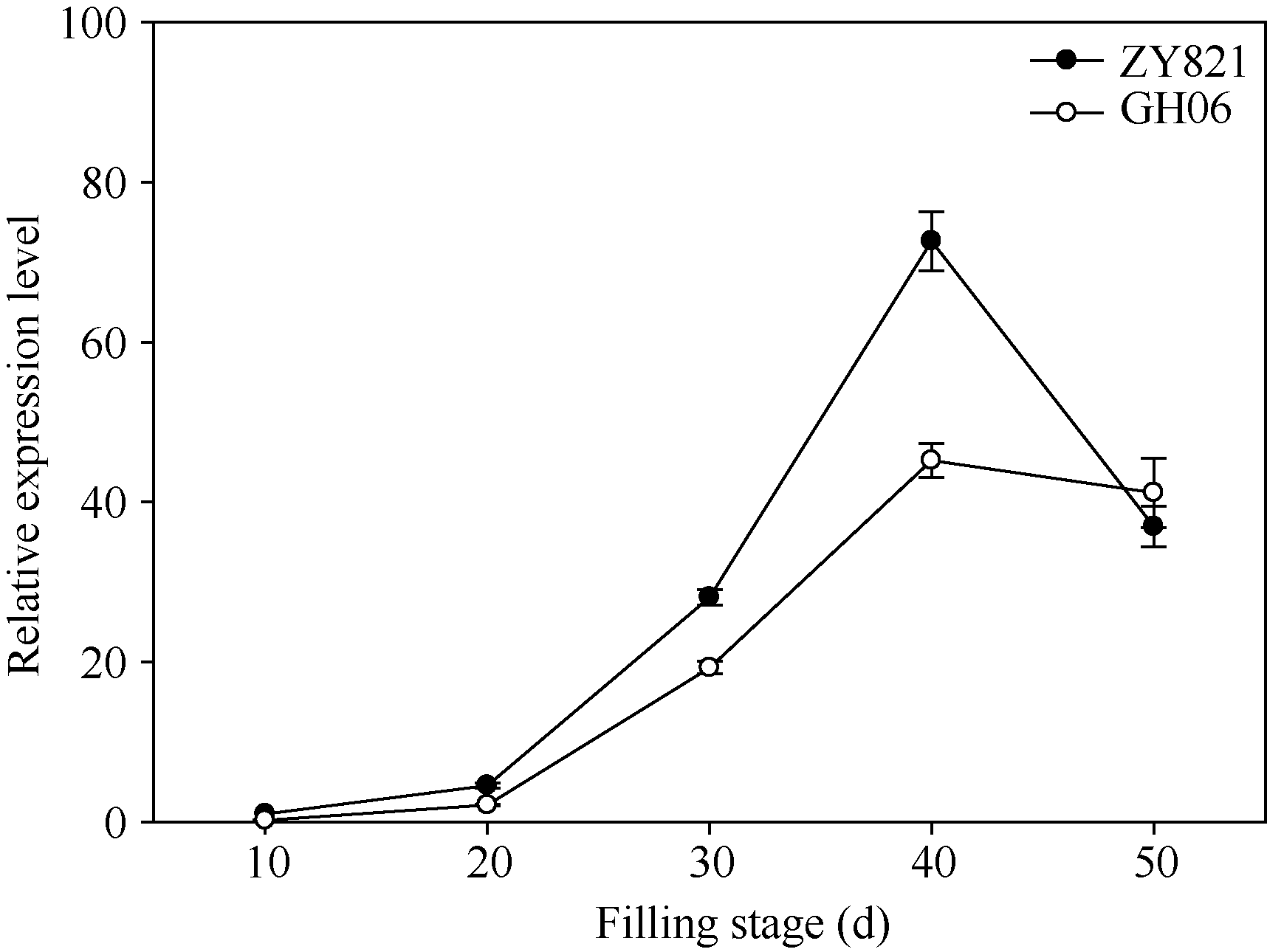 | 图1 BnTT3在黄籽GH06、黑籽ZY821甘蓝型油菜亲本和不同灌浆时种子中表达差异以黑籽亲本ZY821花后10 d种子的表达量为参照计算基因的相对表达量。Fig. 1 Expression patterns of BnTT3 genes from the yellow- (GH06) and black-seeded (ZY821) rapeseed at seed-filling stageRelative gene expression levels were normalized according to the expression values of 10 days old seed in ZY821. |
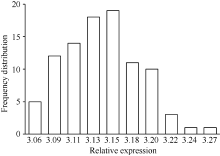
统计分析结果表明, BnTT3基因在后代株系花后40 d种子中表达量呈现典型的数量遗传模式— — 近似正态分布(图2), 其表达不是由单个上游主效基因控制, 而是由多个基因综合调控的, 可以将BnTT3基因表达量作为数量性状进行eQTL分析。
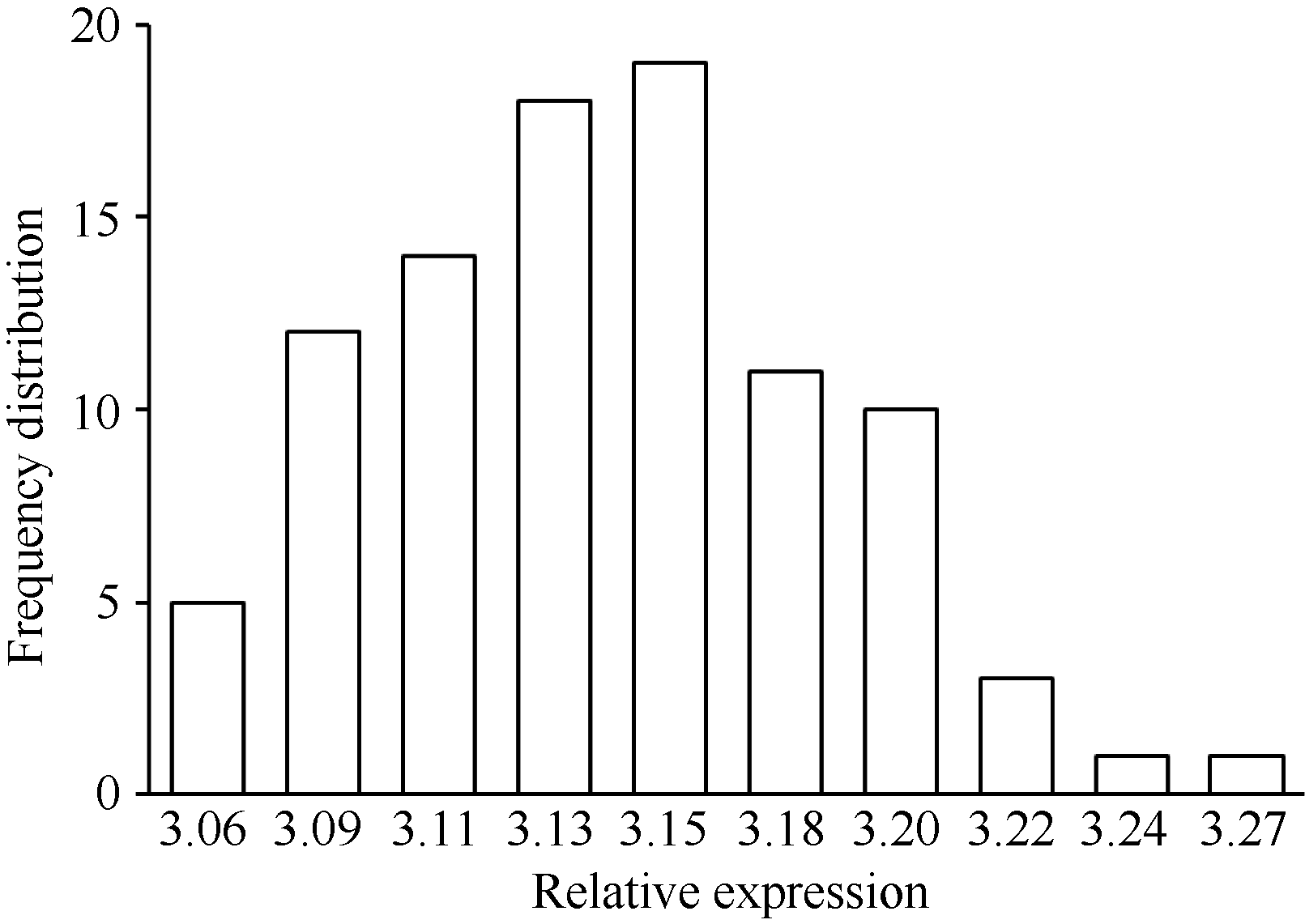 | 图2 重组自交系群体BnTT3基因表达量的频率分布Fig. 2 Frequency distribution of BnTT3 expression levels in B. napus RILs |
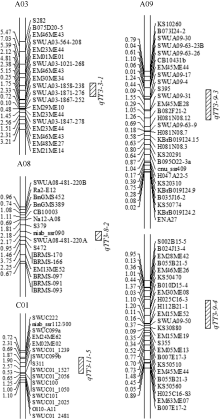
利用复合区间作图法共检测到5个BnTT3基因表达量相关的eQTL, 分别位于A03、A08、A09和C01染色体上(图3), 共解释表型总变异的59.62%, 单个eQTL可解释性状表型变异的变幅介于5.22%~24.05%之间(表1)。其中在A09染色体上存在2个主效eQTL, 单个eQTL分别解释24.05%和16.55%的表型变异, 介于标记KS10260~ KBrB019I24.15和B055B21-5~KS30880之间, 而在A03、A08与C01染色体上仅检测到BnTT3基因的微效eQTL (图3)。为了确定A09染色体上2个主效eQTL的类型, 从甘蓝型油菜基因组序列中提取与eQTL连锁标记SWUA09-17和B010D15-4上下游各200 kb序列, 并以BlastN分析BnTT3基因是否位于该区段。根据eQTL分类标准, 在上述连锁标记相关的eQTL区段未发现BnTT3基因, 因此这些eQTL可能均属于trans-QTL, 即在该标记附近存在BnTT3基因的上游调控基因, 对BnTT3基因的表达起着关键调控作用。从加性效应值还可以看出, BnTT3基因eQTL的增效作用均来自父本ZY821的等位基因。
| 表1 甘蓝型油菜重组自交系中BnTT3基因的eQTL分析 Table 1 eQTLs for BnTT3genes detected from the RILs in B. napus |
 | 图3 以复合区间作图法检测BnTT3基因表达量的eQTLFig. 3 eQTL detection for the expression of BnTT3 with CIM |
为了分析A09染色体上的trans-QTL区段是否存在对TT3基因起调控作用的上游基因, 我们对主效eQTL连锁标记SWUA09-17和B010D15-4上下游各200 kb侧翼序列进行了本地BlastN比对, 结果发现该区间在白菜、甘蓝和油菜染色体上对应的具体位置存在一定的差异(表2), 但它们与芸薹族其他物种间均具有较好的共线性(图4-A, B), 油菜A/C亚基因组间也具有很好的共线性(图4-C, D), 说明该区段序列在进化过程中相对比较保守, 在芸薹属物种中同源性较高, 适宜于基因注释比较分析。
| 表2 连锁标记在芸薹属测序物种染色体上的物理位置 Table 2 Physical locations of linked markers in chromosome of Brassica species sequenced |
 | 图4 两个主要trans-QTL区段侧翼序列在芸薹属及其近缘物种中的共线性A和C: qTT3-9-3区段侧翼序列共线性分析; B和D: qTT3-9-4区段侧翼序列的共线性分析。Fig. 4 Collinearity of flanking sequences of two major trans-QTLs between Brassicaspecies and their relativesA and C: collinearity analysis of flanking sequence in trans-QTL qTT3-9-3; B and D: collinearity analysis of flanking sequence in trans-QTL qTT3-9-4. |
基因注释结果发现, qTT3-9-3区段(连锁标记SWUA09-17)中共有58个基因, qTT3-9-4区段(连锁标记B010D15-4)中存在20个基因, 但在区间范围内并未发现BnTT3和类黄酮途径其他相关基因, 进一步证实检测到的eQTL可能属于trans-QTL。此外, 这2个trans-QTL区段不仅包含重要的转录因子(MYB51、MYB52和bZIP5), 还存在一些与转运及抗病相关的基因(见附表1)。前人研究已证实MYB、MYC和bZIP等转录因子具有调节类黄酮次生代谢途径的功能[26, 27, 28, 29], 因此, 对2个主效eQTL区段这类转录因子的深入分析有助于明确甘蓝型油菜类黄酮途径的调控代谢机制。
| 附表1 两个主要eQTL区段基因列表 Supplementary table 1 List of genes in the two major eQTLs regions |
前人研究认为色素化合物及相关酶类是影响甘蓝型油菜种子发育过程中种皮色泽变化的重要因子, 包括原花色素、多酚和木质素及其衍生物等重要次生代谢物质, 而拟南芥种皮颜色发生不同程度变异的主要原因是一些类黄酮生物合成关键酶基因的突变[9, 30, 31]。迄今为止, 植物花青素和类黄酮物质代谢途径研究比较深入, 已鉴定出23个TT基因位点, 包括12个编码类黄酮生物合成相关酶的结构基因位点, 6个编码控制类黄酮生物合成的调控基因位点以及5个未知的基因位点[30]。其中, TT3基因编码4-二氢黄酮醇还原酶, 该酶在花青素苷积累与类黄酮的合成中起重要作用, 主要负责合成花青素苷的直接底物— 原花青素苷元(黄烷-3, 4二元醇), 其突变体导致种皮中花青素和原花色素合成受阻, 种皮黄色透明, 与野生型的黑色种皮区别明显[1, 20]。此外, TT3基因不仅在圆叶牵牛(Ipomoea purpurea)和水稻胚乳中起关键作用[32, 33], 在芸薹属埃塞俄比亚芥(Brassica carinata)和芥菜型油菜(Brassica juncea)中也是种皮色泽性状遗传调控网络中的关键基因[34, 35]。前人研究表明, 甘蓝型油菜BnTT3是原花色素与花青素代谢途径关键调控基因[20], 其表达量在种皮发育中后期达到峰值, 且黄黑籽间差异极显著, 提示BnTT3在甘蓝型油菜种皮发育后期对黄黑籽差异的形成可能具有重要调控作用。研究还发现, 其他TT基因在黄黑籽甘蓝型油菜中表达模式与BnTT3类似, 仅存在表达量上的差异[21], 表明TT基因可能不是导致甘蓝型油菜黄黑籽差异的主效基因, 其表达差异很可能受上游调控因子调节。本研究也发现在黄黑籽甘蓝型油菜亲本材料中, BnTT3的表达量均随种子发育而不断上升, 并在花后40 d达到峰值, 且黄黑籽材料的表达量差异也在这个时期最明显, 表达模式与前人研究结果完全一致。由于BnTT3在黄黑籽甘蓝型油菜亲本花后40 d种子的表达量差异最大, 因此, 本研究选择了这个时期进行群体株系qRT-PCR检测和eQTL分析, 研究结果表明我们对BnTT3基因的表达量进行了准确的eQTL定位, 这说明基于前期研究结果, 准确选择eQTL定位分析的样品有助于获得更好的eQTL分析结果。
本研究共检测到5个BnTT3基因表达量相关的eQTL, 分别位于A03、A08、A09和C01染色体上。其中, 在A09染色体上检测到2个eQTL (qTT3-9-3和qTT3-9-4), 其置信区间分别位于种皮色泽主效QTL(BnSCA09)的上游与下游[36], 并与种皮色素相关的QTL区间相吻合, 表明这2个eQTL区间内的候选基因可能与种皮色泽主效QTL候选基因相同, 其不仅能够调节BnTT3基因的表达, 而且能控制甘蓝型油菜种皮色素合成和色泽差异的形成。通过连锁标记侧翼序列比对分析发现, 2个eQTL区间的序列在芸薹属内同源性较高, 相应区间基因注释后并未发现BnTT3及类黄酮途径其他相关基因, 说明本研究定位的eQTL很可能属于trans-QTL, 包含调节BnTT3基因的上游调控因子。基因功能分析发现, 2个R2R3-MYB转录因子MYB51和MYB52位于上游主效QTL置信区间内。拟南芥中的研究证实MYB51是调节吲哚硫苷生物合成的重要基因, 同时介导水杨酸和乙烯信号途径[37]; MYB52主要参与调节次生细胞壁的形成, 介导脱落酸信号途径[38]。目前, 2个基因在油菜中功能尚未被证实, 现有证据尚无法确认它们是否参与调控类黄酮途径。但大量的研究已经表明R2R3-MYB类转录因子不仅广泛参与植物苯丙烷-类黄酮代谢途径中的多种初生和次生代谢过程[39, 40], 同时还介导水杨酸、乙烯及脱落酸等多个信号途径[37, 38]。有研究发现, 生长素、乙烯及茉莉酸等途径可影响类黄酮的合成与积累[20, 41]。因此, 这2个MYB转录因子是否通过植物激素信号途径间接影响类黄酮代谢途径, 实现其对BnTT3基因的调控值得进一步深入研究。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|