




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
水稻抗稻瘟病基因 Pi35功能性分子标记的开发及其应用
[马建*  , 马小定
, 马小定* , 赵志超, 王帅, 王久林, 王洁, 程治军雷财林* , 雷财林* ]
, 马小定, 赵志超, 王帅, 王久林, 王洁, 程治军雷财林]
|
|


第一作者联系方式: 马建, E-mail: jian.ma2018@aliyun.com; 马小定, E-mail: maxiaoding@caas.cn
稻瘟病是水稻生产上的严重病害, 利用抗病基因培育抗病品种是控制稻瘟病最经济而有效的措施。在日本, 稻瘟病部分抗性基因 Pi35作为广谱持久抗性基因已广泛应用于水稻育种和稻瘟病防治实践。但是, Pi35基因在我国的资源和品种中的分布情况不清, 制约了这一重要基因在我国育种实践中的应用, 急需开发实用的分子标记, 并系统研究该基因在我国的品种及其亲本中的分布情况, 为稻瘟病抗性育种服务。本研究通过比对抗、感品种中 Pi35等位基因序列, 发现一个能检测抗、感病性差异的特异SNP (3780 T), 并据此开发了 Pi35基因的功能性分子标记Pi35- dCAPS。利用该标记检测了抗源藤系138的衍生品种10份、微核心种质204份和主栽品种67份, 结合测序鉴定, 确认5份藤系138衍生品种(垦鉴稻3号、垦鉴稻6号、垦稻8号、绥粳3号和龙粳34)及2份微核心种质(粳稻品种抚宁紫皮粳子和籼稻品种细麻线)携带 Pi35基因。本研究结果为通过分子育种手段高效利用 Pi35基因改良我国水稻(特别是籼稻)品种的稻瘟病抗性提供了手段。
, MA Xiao-Ding, ZHAO Zhi-Chao, WANG Shuai, WANG Jiu-Lin, WANG Jie, CHENG Zhi-Jun, LEI Cai-Lin
Rice blast is one of the most destructive diseases, and breeding resistant cultivars is considered to be the most economical and effective strategy to control this disease. The Pi35gene shows partial resistance to leaf blast and has been used as a broad-spectrum and durable resistance source in rice breeding programs in Japan. However, its distribution is not clear in Chinese rice germplasm and cultivars. For the purpose to facilitate the application of Pi35 in rice breeding programs in China, we compared the coding sequences of Pi35 alleles in multiple resistant and susceptible rice cultivars, found a specific nucleotide 3780T which was only present in the functional resistance allele of Pi35, and further developed a Pi35functional marker (Pi35-dCAPS). Among 281 rice accessions including 10 Fukei 138-derived japonicacultivars, 67 leading cultivars, and 204 accessions of rice mini-core collection of Chinese germplasm, five Fukei 138-derived cultivars (Kenjiandao 3, Kenjiandao 6, Kendao 8, Suijing 3, and Longjing 34) and two mini-core accessions ( japonicacv. Funingzipijingzi and indica cv. Ximaxian) were detected to possess the intact Pi35 gene by using the Pi35-dCAPS marker in combination with the genomic sequencing of Pi35. These results will greatly facilitate the utilization of Pi35 in rice breeding programs by marker-assisted selection.
稻瘟病是水稻生产上最主要的病害之一, 严重影响稻谷的产量和质量[1]。利用寄主抗病基因培育并种植抗病品种是控制稻瘟病最安全、经济和有效的措施[2]。水稻对稻瘟病的抗性可被划分为完全抗性(或称质量抗性、主效基因抗性、垂直抗性等)和部分抗性(或称数量抗性、微效基因抗性、田间抗性等)[1, 3]。完全抗性能完全抵抗稻瘟病菌的侵染和繁殖而表现高度抗性, 一般受单个或几个主效基因控制, 这类抗性具有很强的小种专化性, 容易随着小种的变化而丧失[1, 4, 5]。部分抗性一般被视为数量性状, 它不能阻止稻瘟病菌的侵染, 但可以减少病原菌在寄主体内的增殖, 起到降低病斑数目和病斑大小而减轻病害的作用。大多数部分抗性受多个微效基因或数量性状位点(quantitative trait loci, QTL)控制, 无明显的小种专化性, 具有这类抗性的水稻品种往往因其降低了对病原菌的选择压而保持稳定和持久的抗性[4, 6, 7]。因此, 提高部分抗性水平一直是水稻抗稻瘟病育种的一个重要目标[4, 8]。
随着分子生物学和QTL定位技术的发展, 水稻稻瘟病抗性基因研究取得很大进展, 迄今已鉴定和定位90多个完全抗性基因及300多个抗性QTL或部分抗性基因, 克隆了Pia、Pib、Pita、Pi1、Pi2、Pi5、Pi9、Pi36、Pi37、Pi54、Pi64、Pi-d2、Pi-d3、Pik、Pik-m、Pik-p、Pik-h、Pish、Pit、Piz-t等20个完全抗性基因, 以及pi21、Pb1、Pi63和Pi35等4个部分抗性基因(http://www.ricedata.cn/gene/gene_ pi.htm)[7, 9, 10, 11, 12]; 并开发了针对Pita[13]、Pib[14]、Pik[15]、Pik-m[15]、Pik-p[16]、Pi1[17]、Pit[18]、Pi5[19]、Pi25[20]、Pi54[21]和Pi64[22]等11个完全抗性基因的功能性标记。Pi35是最近被克隆的一个稻瘟病部分抗性基因, 是Pish的等位变异基因[7]。携带Pi35基因的粳稻品种北海188 (Hokkai 188)和藤系138 (Fukei 138)在日本自育成30多年来一直保持稳定的高水平的叶瘟抗性[6, 7, 23]; 其中, 藤系138在上世纪80年代中期被引入黑龙江、吉林等省的有关育种单位, 并以之作为抗性亲本培育了一批推广品种[24]。但是, Pi35基因在我国的资源和品种中的分布情况不清, 制约了这一重要基因在我国育种实践中的应用, 急需开发实用的分子标记, 并系统研究该基因在我国的品种及其亲本中的分布情况, 为稻瘟病抗性育种实践服务。本研究拟通过测序和比对多个抗病和感病水稻品种的Pi35等位基因序列, 在开发出Pi35基因特异功能性分子标记的基础上, 对我国水稻核心种质资源和部分育成品种进行鉴定, 以期发掘携带Pi35基因的新种质, 推进我国水稻(特别是籼稻)品种的稻瘟病持久抗性育种工作。
供试水稻材料包括抗性品种藤系138, 感病对照品种丽江新团黑谷(LTH)和日本晴; 藤系138的衍生品种10份(垦鉴稻3号、垦鉴稻6号、垦稻8号、垦稻12、垦稻16、绥粳3号、龙花00-835、龙粳10号、龙粳13和龙粳34); 水稻微核心种质204份及主栽培品种67份(见附表)。其中, 藤系138和微核心种质由中国农业科学院作物科学研究所国家农作物种质资源平台水稻中期库提供, 藤系138的衍生品种由黑龙江农垦科学院水稻研究所和黑龙江省农业科学绥化分院提供, 67份水稻主栽培品种由中国水稻研究所国家水稻种质资源中期库提供, 品种LTH和日本晴为本实验室保存的材料。
供试稻瘟病菌株包括日本致病性稳定菌株Ken 54-20(小种003)和Ken 53-33(小种137.1), 用于藤系138及Pi35基因的部分抗性评价[6, 7, 23, 25]。
| 附表 Table Supplementary table Screening of the Pi35 gene in the mini-core collection of rice germplasm and contemporary leading cultivars in Chinay |
将供试水稻材料浸种至露白后, 穴播于装有草炭土的塑料育苗盘(54 cm × 28 cm × 4 cm, 60穴), 每穴10~15粒种子, 重复3次。待稻苗长至二叶一心叶时, 用木村营养液B[26]定期浇灌。待稻苗生长至四叶一心时喷雾接种稻瘟病菌[27]。7 d后, 参考Mackill和Bonman[5]的标准调查病情。0级为无任何病斑; 1级为直径小于0.5 mm的褐色斑点; 2级为直径0.5~1.0 mm的褐色病斑; 3级为直径1~3 mm的圆形或椭圆形病斑, 中央灰白色, 边缘褐色; 4级为典型的纺锤形病斑, 直径3 mm或更长, 病斑无融合或稍有融合; 5级为同4级病斑, 但病斑融合, 叶片的上半部枯死。0~1级视为抗病(R), 2级视为中抗(MR), 3级视为中感(MS), 4~5级视为感病(S)。
利用CTAB法[28]提取叶片基因组总DNA。PCR体系为20 μ L, 包括10 μ L 2× PCR buffer for KOD FX、4 μ L dNTP (2 mmol L-1 each)、1 μ L Primers (10 mmol L-1, F+R)、0.4 μ L KOD FX (1 U μ L-1, TOYOBO)、2 μ L基因组DNA (100 ng μ L-1)和2.6 μ L ddH2O。PCR程序为94℃ 2 min; 98℃ 10 s, 59℃ 15 s, 68℃ 0.5~8.0 min (依片段大小而定, 1 kb min-1), 35个循环; 68℃延伸5 min。酶切反应体系为25 μ L, 包括20 μ L PCR产物、2 μ L的10× FastDigest green buffer、1 μ L的Tai I内切酶(Thermo Scientific, USA)和2 μ L的ddH2O, 65℃酶切3 h。PCR产物由北京博迈德生物有限公司测序, 用DNAMAN软件(http://www.lynnon. com/)拼接。酶切产物经2.5%琼脂糖凝胶电泳检测。
从Gramene网站(http://www.gramene.org/)获得Pish基因序列(Os01g0782100, 登录号为NM_001185663.1), 从NCBI数据库(http://www.ncbi.nlm.nih.gov/)获得Pi35基因序列(登录号为FW369319.1)。从感病品种丽江新团黑谷(LTH)中扩增出这2个基因的等位变异序列。利用软件dCAPS Finder 2.0 (http://helix.wustl. edu/dcaps/dcaps.html)和NCBI网站程序Primer- BLAST (http://www.ncbi.nlm.nih.gov/tools/primer-blast/ index.cgi?LINK_LOC=BlastHome)设计dCAPS标记和PCR扩增用引物(表2), 引物由上海英潍捷基贸易有限公司合成。
| 表2 本研究所用的PCR引物序列 Table 2 PCR primer sequences used in this study |
利用ZR Plant RNA MiniPrep Kit (ZYMO, USA)提取水稻幼苗总RNA, 用DNase I (TaKaRa, 大连)消化基因组DNA 30 min。取5 μ g总RNA合成第1链cDNA (TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix试剂盒, TransGen Biotech, 北京)。采用RT-PCR方法检测基因的表达情况, PCR体系含10 μ L 2× PCR buffer for KOD FX, 4 μ L dNTP (2 mmol L-1 each), 1 μ L Primers (10 μ mol L-1, F+R), 0.4 μ L KOD FX (1 U μ L -1, TOYOBO), 1 μ L反转录产物, 3.6 μ L ddH2O; 反应程序为94℃ 2 min; 98℃ 10 s, 57℃ 15 s, 68℃ 45 s, 28~33个循环; 68℃ 5 min, 4℃保存; 用1.5%的琼脂糖凝胶电泳检测, 紫外光下照相。
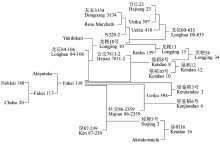
Pi35基因克隆自日本品种北海188[7]。藤系138是北海188的衍生品种(图1), 它继承了北海188的高水平部分抗性(图2)。之前的研究认为, 藤系138的抗性由一个主效基因控制, 可能为Pi35基因, 但缺乏更进一步的证据[6, 23]。为确认藤系138携带的就是Pi35基因, 我们利用引物Pi35-F/R (表2)对藤系138的基因组DNA进行PCR扩增、测序、拼接和序列比对, 发现扩增出的DNA序列及氨基酸与数据库中公布的Pi35的序列完全一致(图3)。综合前人[6, 23]及本研究的结果, 我们确信藤系138携带部分抗性基因Pi35, 并将其命名为Pi35-Fukei138。
 | 图1 藤系138及其衍生品种系谱图Fig. 1 Pedigree of the rice cultivar Fukei 138 and its derived cultivars |
 | 图2 藤系138、日本晴和LTH对2个稻瘟病鉴别菌株的反应型Fig. 2 Phenotypes of Fukei 138, Nipponbare, and LTH to two Magnaporthe oryzae isolates |
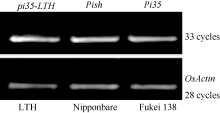
Pi35是Pish的等位变异基因[7], 在日本晴中Pish/Pi35基因簇包含4个串联的NBS-LRR基因Os01g0781100、Os01g0781200、Os01g0781700和Pish[29]。为了设计和开发特异性鉴定Pi35基因的功能性标记, 我们首先从数据库中获得日本晴中上述4个基因的编码区DNA序列, 经序列比对发现第一个基因Os01g0781100与后面3个基因相似性较低(氨基酸序列一致性分别为65.7%、65.6%和65.5%), 存在着较多的碱基差异, 因此, 未将该基因用于后续序列比较。利用引物Pi35-F/R对普感品种LTH[25, 29, 31]的基因编码区进行扩增, 所得产物被命名为pi35-LTH (图3)。利用ClustalW软件对5个基因(Pi35、pi35-LTH、Pish、Os01g0781200和Os01g0781700)编码的蛋白质序列分析发现, Pi35与pi35-LTH之间仅在LRR区(位置589-1289)存在一个氨基酸的差异(C1260W); Pi35与Pish以及Os01g0781200和Os01g0782100之间在该位置也存在差异, 第1260位的半胱氨酸为Pi35蛋白所特有(图3)。进一步利用引物Pi35-C-F/R对7796 bp (ATG前面2041 bp、3870 bp Pi35编码区和TGA下游1885 bp)目的基因组片段进行扩增、测序, 结果发现, 抗病品种藤系138和感病品种LTH在包括启动子的周边序列上没有其他差异。表达分析结果表明, 基因Pi35、Pish及pi35-LTH在各自的背景品种中均能正常表达(图4), 不存在其他影响基因表达量的因素。因此, 第1260位的氨基酸的差异是Pi35与pi35-LTH基因抗、感病性差异的决定因素, 可以被用于开发特异性的功能性标记。
 | 图3 Pi35/Pish位点基因氨基酸序列比对箭头表示Pi35蛋白中第1260位特异的半胱氨酸。Fig. 3 Multiple alignment of deduced amino acid sequences of Pi35/Pishlocus Arrow indicates the 1260th specific cysteine of Pi35 protein. |
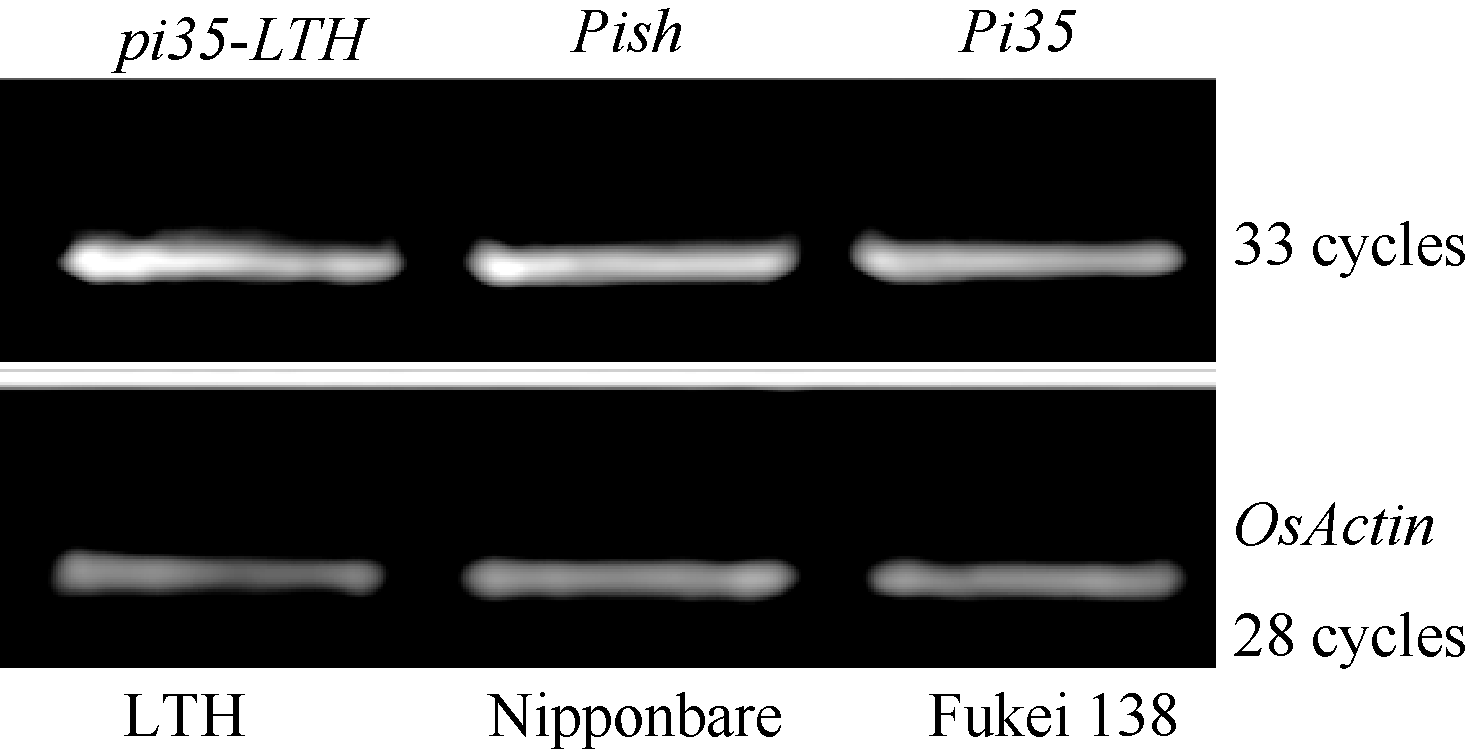 | 图4 3个等位基因pi35-LTH、Pish和Pi35的RT-PCR分析Fig. 4 RT-PCR analysis of three allelic genes of pi35-LTH, Pish, and Pi35 |
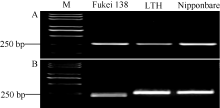
基于藤系138和LTH在该位点上的序列差异, 开发了一个特异性识别Pi35等位基因的标记Pi35-dCAPS (表2和图5)。该标记在藤系138 (Pi35供体)、LTH和日本晴(Pish供体)中均能扩增出一条长度为272 bp的目标片段(图6-A), 藤系138的产物可以被Tai I酶切成241 bp和31 bp两段, 由于缺乏酶切位点, LTH和日本晴的产物均不能被切开, 在琼脂糖电泳胶上清楚地显示出Pi35的特异条带(图6-B)。
 | 图5 DNA序列多态性和标记Pi35-dCAPS引物的位置Fig. 5 DNA polymorphism and positions of primers for Pi35-dCAPS marker |
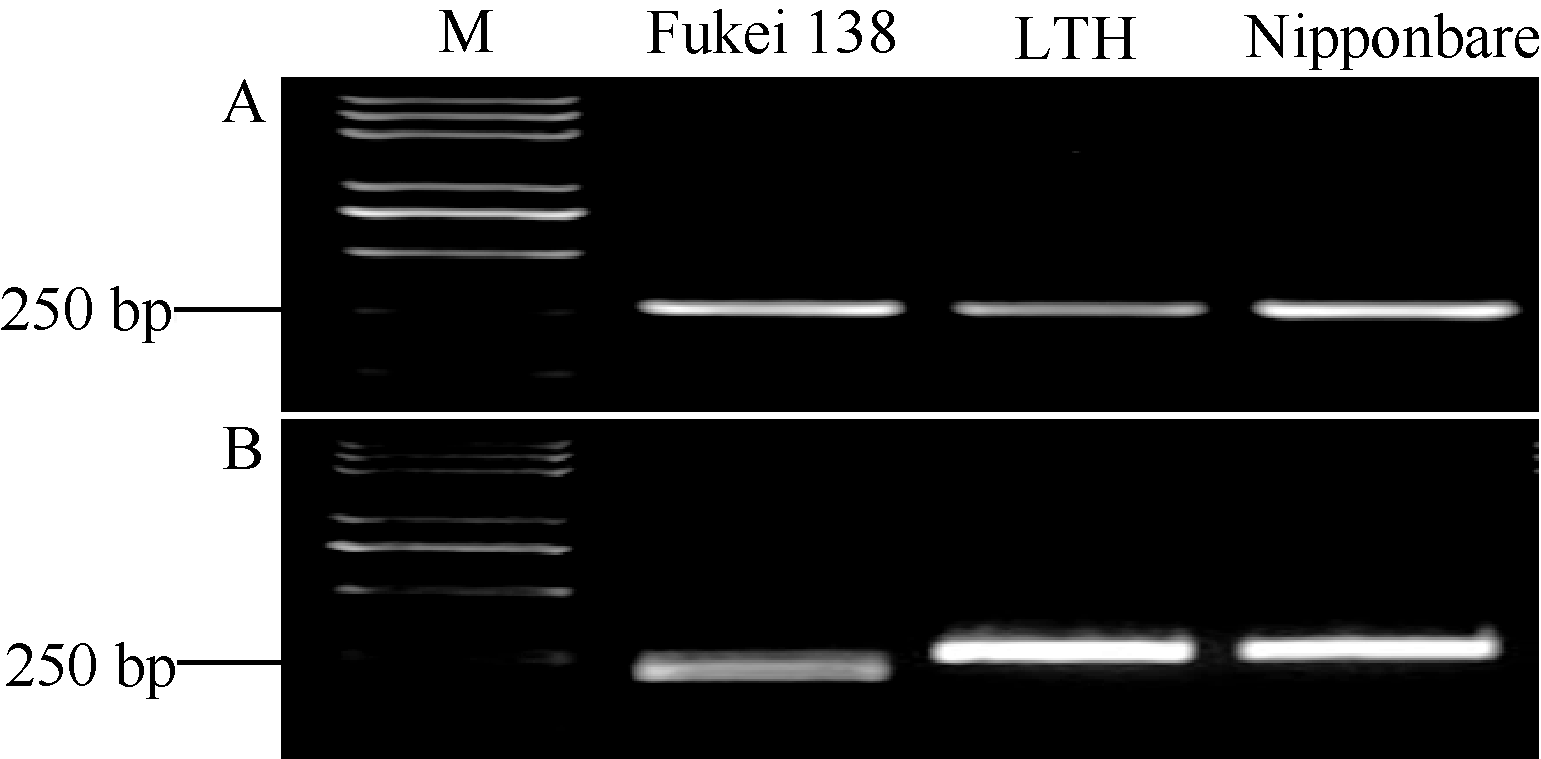 | 图6 Pi35-dCAPS标记在3个水稻品种中的多态性 Fig. 6 Genotypic analysis of Pi35-dCAPS marker in three rice cultivars |
利用Pi35-dCAPS标记鉴定10份藤系138衍生的粳稻品种(垦鉴稻3号、垦鉴稻6号、垦稻8号、垦稻12、垦稻16、绥粳3号、龙花00-835、龙粳10号、龙粳13和龙粳34) (http://www.ricedata.cn/ variety/, 图1)的Pi35基因型, 发现其中的5个(垦鉴稻3号、垦鉴稻6号、垦稻8号、绥粳3号和龙粳34)可以被Pi35-dCAPS标记识别。进一步利用引物Pi35-F/R对这5份品种的Pi35等位基因编码区扩增、测序表明, 上述5个品种确实含有Pi35基因(图7)。上述结果证实, Pi35-dCAPS和Tai I组合可以特异地鉴定不同水稻品种中Pi35基因的存在。
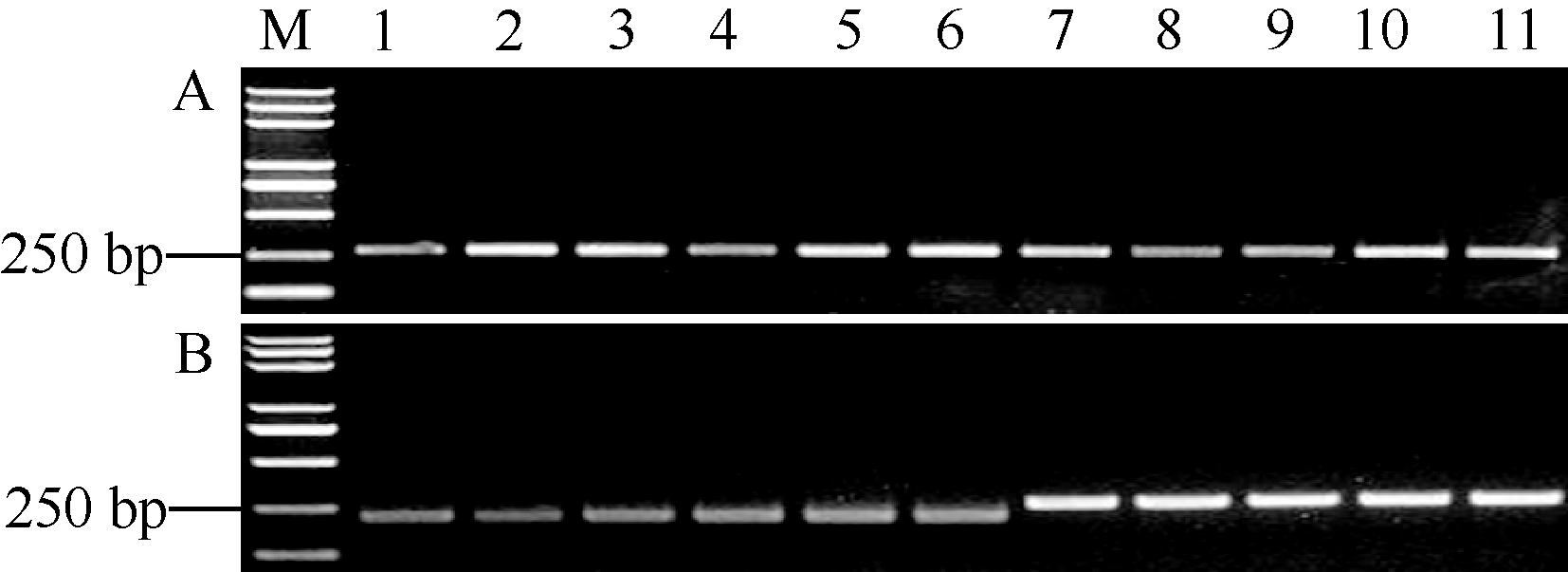 | 图7 Pi35-dCAPS标记对10份藤系138衍生品种检测的结果 Fig. 7 Genotypic analysis in ten Fukei 138-derived japonicacultivars with Pi35-dCAPS marker |
利用Pi35-dCAPS标记对204份水稻微核心种质及67份水稻主栽品种(附表)进行PCR检测, 所有供试材料均可特异地扩增出一条272 bp的目标片段, 经限制性内切酶Tai I酶切这些扩增产物, 仅有2份微核心种质材料(抚宁紫皮粳子和细麻线)的PCR片段可以被Tai I酶切而呈现与藤系138一致的带型(图8)。因此, 推断在供试的271份材料中只有1份粳稻材料抚宁紫皮粳子和1份籼稻材料细麻线携带Pi35基因(见附表)。进一步利用引物Pi35-F/R对这2份品种的Pi35等位基因编码区进行扩增, 经测序、拼接和比对发现这2份品种的基因编码区序列与Pi35基因完全一致, 证明这2份品种确实携带Pi35基因。
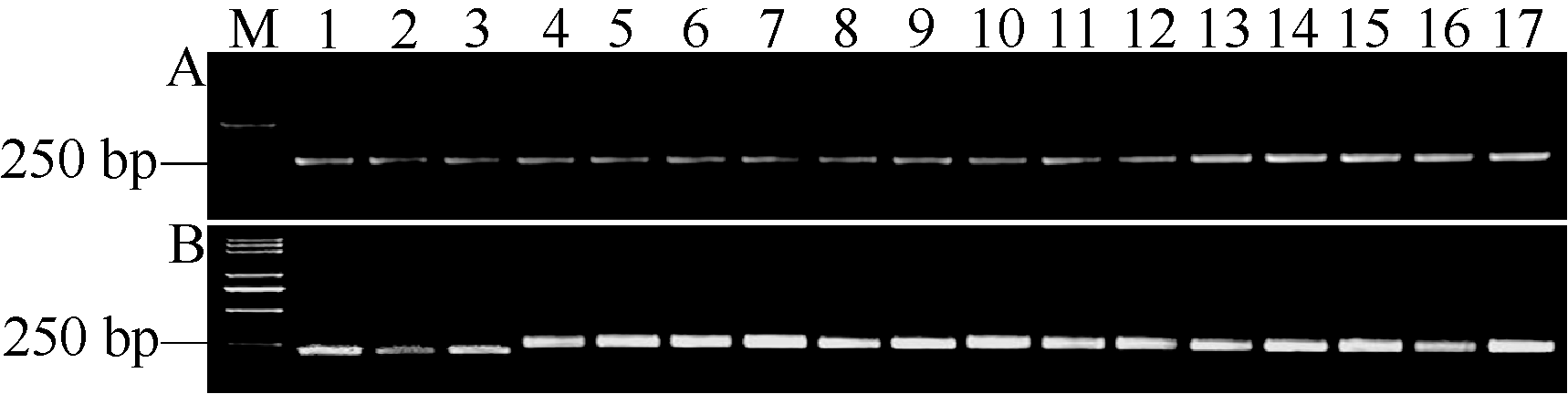 | 图8 Pi35-dCAPS标记在部分水稻种质资源中检测的结果 Fig. 8 Genotypic analysis in partial rice accessions with Pi35-dCAPS marker |
与完全抗性基因相比, 部分抗性基因通常无明 显的小种专化性, 其抗性表现广谱而持久, 因此, 利用部分抗性基因被视为最有希望能持续控制稻瘟病的措施[4, 6, 7, 32, 33, 34, 35, 36]。目前已报道的300多个稻瘟病抗性QTL中, 绝大部分在染色体上的定位区间较大, 缺乏紧密连锁的分子标记[37], 尽管已有4个部分抗性基因(pi21、Pb1、Pi63和Pi35)被克隆, 但迄今尚未见其功能性标记开发的报道。这在很大程度上限制了这些QTL或基因在水稻抗稻瘟病育种中的广泛应用。有针对性地选择一些重要的主效QTL或部分抗性基因, 开发与其紧密连锁的分子标记, 尤其是基因位点特异性标记和基因功能性标记, 对于精准鉴定水稻品种的部分抗性基因型, 以及通过基因聚合手段培育持久抗病品种具有重要意义。
部分抗性基因Pi35源自日本品种北海188, 该品种自1961年育成至今一直保持着稳定的高水平叶瘟抗性[6, 7, 23]。由于缺乏北海188的种子, 我们通过对北海188衍生品种藤系138中Pi35等位基因进行基因组全长测序, 确认了藤系138携带Pi35基因。通过分析高度感病的水稻品种LTH的Pi35等位基因pi35-LTH的DNA和蛋白质序列, 发现Pi35与pi35-LTH之间仅在LRR端存在一个氨基酸的差异(C1260W), 且第1260位的半胱氨酸为Pi35蛋白所特有(图2)。Fukuoka等[7]指出, Pi35基因LRR端的多个功能性多态位点(1053D、1056S、1073P和1083W)的累积强化了该基因的抗性, 且第1053位的天冬氨酸残基与该基因介导的数量抗性最相关。在基因组全长测序和基因表达分析的基础上, 我们通过多次重复测序验证了pi35-LTH与Pi35序列在这些位点上并无序列差异(图3), 我们认为Fukuoka等[7]关于“ 单个抗性基因中多个功能性多态性位点提高稻瘟病持久抗性” 的观点值得商榷, 推断第1260位的半胱氨酸是Pi35蛋白一个功能氨基酸。
基于Pi35和pi35-LTH基因DNA序列本身的一个差异位点T3780G, 我们开发了一个可特异鉴定Pi35基因的功能性分子标记Pi35-dCAPS, 利用此标记从281份水稻材料(10份藤系138衍生品种、204份微核心种质和67份主栽品种)中鉴定出7份含有Pi35基因的材料, 其中5份为藤系138衍生品种(垦鉴稻3号、垦鉴稻6号、垦稻8号、绥粳3号和龙粳34), 2份为微核心种质(抚宁紫皮粳子, 细麻线)。这一结果说明, Pi35基因在藤系138衍生品种以外的国内水稻种质中分布频率很低, 而在籼稻品种细麻线中鉴定到Pi35则为利用该基因改良籼稻品种的稻瘟病部分抗性提供了便利。
证实了水稻品种藤系138携带部分抗性基因Pi35, 鉴定出一个特异存在于Pi35基因编码区的碱基位点3780T, 开发了一个可特异鉴定Pi35基因的功能性分子标记Pi35-dCAPS。利用该标记从281份国内水稻种质资源中鉴定出7份携带Pi35基因的种质, 证实Pi35基因在除藤系138衍生品种以外的国内水稻种质中分布频率很低, 而籼稻种质细麻线携带Pi35基因则为利用该基因改良籼稻品种的稻瘟病部分抗性提供了便利。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|