
{kind=link}
弱筋小麦宁麦9号及其衍生系的蛋白质含量遗传多样性及关联分析
[姜朋, 张平平, 张旭, 陈小霖, 姚金保, 马鸿翔*  ]
]
]
|
|


第一作者联系方式: E-mail: hmjp2005@163.com
优质弱筋专用小麦品种宁麦9号是江苏淮南麦区小麦育种的重要亲本, 以其为亲本已育成15个新品种。研究宁麦9号遗传信息的传递特点及蛋白质含量的数量性状位点, 对进一步利用其进行优质弱筋小麦育种具有重要意义。以宁麦9号及其117个衍生品种(系)为材料, 利用覆盖小麦全基因组的185对SSR引物对其进行基因组扫描, 解析宁麦9号遗传信息在其衍生品种(系)中的分布特点, 同时于2009—2010、2010—2011连续2个生长季测定宁麦9号及其衍生品种(系)的籽粒及面粉蛋白质含量, 应用全基因组关联作图发掘与其相关联的分子标记位点。结果显示, 宁麦9号与其衍生品种(系)的遗传相似系数为0.55~0.88; 在Neighbor-Joining聚类图中, 大部分衍生一代品种首先与宁麦9号聚类, 其次是衍生二代, 衍生一代品种扬辐麦4号最后聚类。宁麦9号与其衍生一代和衍生二代相同等位变异频率分别为75.60%和67.81%。籽粒及面粉蛋白含量在宁麦9号衍生品种(系)中均呈现较大变异, 变异系数为5.07%~7.28%。共检测到6个与籽粒蛋白质含量关联的标记位点, 其中3个连续两年均检测到; 面粉蛋白质含量共有5个关联位点, 其中2个连续两年检测到, 包括1个同时与籽粒和面粉蛋白质含量关联。在这4个稳定的关联位点中, Xgwm539、 Xwmc397和 Xwmc468对籽粒及面粉蛋白质含量起负向调控作用, 可以降低籽粒或面粉蛋白质含量。
Ningmai 9 is an elite weak gluten wheat cultivar and an important breeding parent in the southern area of Huai River Valley. To date, 15 new cultivars have been developed and released from Ningmai 9. This study aimed at dissecting the genetic mechanism and inheritance of protein content in Ningmai 9 and its derivatives. The kernel protein content (KPC) and flour protein content (FPC) of Ningmai 9 and its 117 derivatives were evaluated in the 2009-2010 and 2010-2011 growing seasons and the QTLs associated with KPC and FPC were identified by whole genomes screening with 185 SSR markers. The genetic similarity index ranged from 0.55 to 0.88 among Ningmai 9 and its derivatives. In the Neighbor-Joining cluster tree, Ningmai 9 was clustered with most first-generation lines first, then with the second-generation lines, and Yangfumai 4 of first-generation derivative was the last one to incorporate in. The first- and second-generation derivatives inherited 75.60% and 67.81% of Ningmai 9 alleles, respectively. Great variations of KPC and FPC were observed in the Ningmai 9 derived lines with the variation coefficients ranging from 5.07% to 7.28%. Six and five QTLs were identified to be associated with KPC and FPC, of which three and two were stably detected in both years, respectively. One QTL was associated with both KPC and FPC. Three stable QTLs, Xgwm539, Xwmc397, and Xwmc468, had negative effects on KPC or FPC in Ningmai 9 and might be used for quality improvement in weak-gluten wheat breeding.
蛋白质含量是小麦籽粒的重要组分, 不仅影响小麦的营养品质, 同时也是小麦加工品质的基础。蛋白质含量为典型的数量性状, 受多基因控制, 已有很多关于其分子标记和QTL定位的研究报道。Prasad等[1]利用近等基因系在7条染色体上定位到13个籽粒蛋白质含量QTL, 其中7A染色体上的一个QTL的遗传贡献率高达32.44%; 吴云鹏等[2]利用RIL群体, 在3A和3B染色体上检测到控制籽粒蛋白质含量的QTL; 沈玮囡等[3]在1B等7条染色体上定位到控制籽粒和面粉蛋白质含量的QTL, 发现多个遗传贡献率超过10%的QTL; Bordes等[4]利用372份来自世界各地的小麦材料及803个分子标记在15条染色体上关联到14个籽粒蛋白质含量相关的分子标记; Jochen等[5]以207份欧洲冬小麦品系为材料, 利用115个分子标记在4条染色体上关联到4个籽粒蛋白质含量相关的分子标记。
我国长江中下游麦区具有不利于小麦籽粒蛋白质形成和积累的生态条件, 适宜发展弱筋小麦, 已经成为我国弱筋小麦生产的优势产业带[6]。低蛋白质含量是弱筋小麦品质的重要指标, 国家优质小麦标准GB/T 17893-1999规定优质弱筋小麦蛋白质含量应在11.5%以下。宁麦9号是江苏省农业科学院选育的优质弱筋专用小麦品种, 具有高产、稳产和广泛的适应性及抗小麦黄花叶病、赤霉病等特点, 近年来已成为长江中下游麦区小麦育种的重要亲本, 以其为亲本已经育成15个小麦新品种。该品种弱筋品质稳定, 在饼干和蛋糕加工品质上可与美国、澳大利亚进口的优质软质小麦媲美[7]。研究宁麦9号遗传信息的传递特点, 特别是蛋白质含量的遗传机制, 发掘与其低蛋白含量相关的分子标记, 对弱筋小麦育种具有重要意义。姚金保等[8]利用7个蛋白质含量有差异的软质小麦品种为亲本, 以双列杂交法配制组合, 研究小麦籽粒蛋白质含量的遗传, 发现宁麦9号籽粒蛋白质含量的一般配合力最高, 可显著降低后代蛋白质含量。后来的试验进一步证实, 籽粒硬度、蛋白质含量、面筋强度、碱水保持力等弱筋小麦品质相关指标仍以宁麦9号一般配合力最好, 在优质弱筋小麦育种中具有较高利用价值[9, 10]。利用9个宁麦9号衍生品种与宁麦9号进行遗传相似性分析, 结果显示这些衍生品种遗传背景一半以上来自宁麦9号[11]。本研究拟通过较大样本的宁麦9号衍生品系群体的基因型分析, 揭示其对后代的遗传贡献, 并利用关联分析发掘控制蛋白质含量的标记位点, 阐明其遗传特点, 为更好地应用宁麦9号优质弱筋性状提供依据。
以宁麦9号及其117个衍生品种(系)为试验材料, 其中衍生一代39份, 衍生二代78份(见附表)。2009— 2010、2010— 2011连续2个生长季将供试材料种植于江苏省农业科学院试验基地, 3行小区, 常规田间管理。2010年和2011年分别收获种子晾干, 利用Perton DA7200近红外品质测定仪按照GB/T 24899-2010方法测定籽粒蛋白含量(kernel protein content, KPC); 用Buhler 202磨粉机将籽粒磨成面粉后, 按照GB/T 24871-2010方法测定面粉蛋白含量(flour protein content, FPC)。
选取每个品种幼嫩的叶片5~6 cm, 采用SDS-酚法提取DNA[12]。依据Somers等[13]的遗传连锁图选择SSR标记, 选择覆盖全基因组的185对SSR引物, 每条染色体5~14对, 平均8.8对。PCR反应体系为10 μ L, 包含10× buffer 1 μ L、15 mmol L-1 MgCl2 0.6 μ L、2 mmol L-1 dNTP 0.8 μ L、20 ng μ L-1引物1 μ L、0.1 μ L Taq酶、20 ng μ L-1模板DNA 3 μ L、去离子水3.5 μ L。PCR扩增程序为94℃预变性5 min; 94℃变性45 s, 50~60℃ (依引物而定)退火45 s, 72℃延伸90 s, 36个循环; 72℃延伸10 min。扩增产物经聚丙烯酰胺凝胶电泳检测。在相同位点上, 有带记为1, 无带记为0, 缺失记为9。
以Neighbor-Joining聚类基本将宁麦9号及其衍生品种(系)按世代分为两类(图1), 衍生一代品种(系)多先与宁麦9号聚类, 其次是衍生二代, 扬辐麦4号因经辐射诱变产生新变异而最后聚类。宁麦9号与其衍生品种(系)的遗传相似系数为0.55~0.88, 衍生一代的遗传相似系数多高于0.70, 衍生二代则较衍生一代低0.10左右, 该结果与聚类次序基本一致。
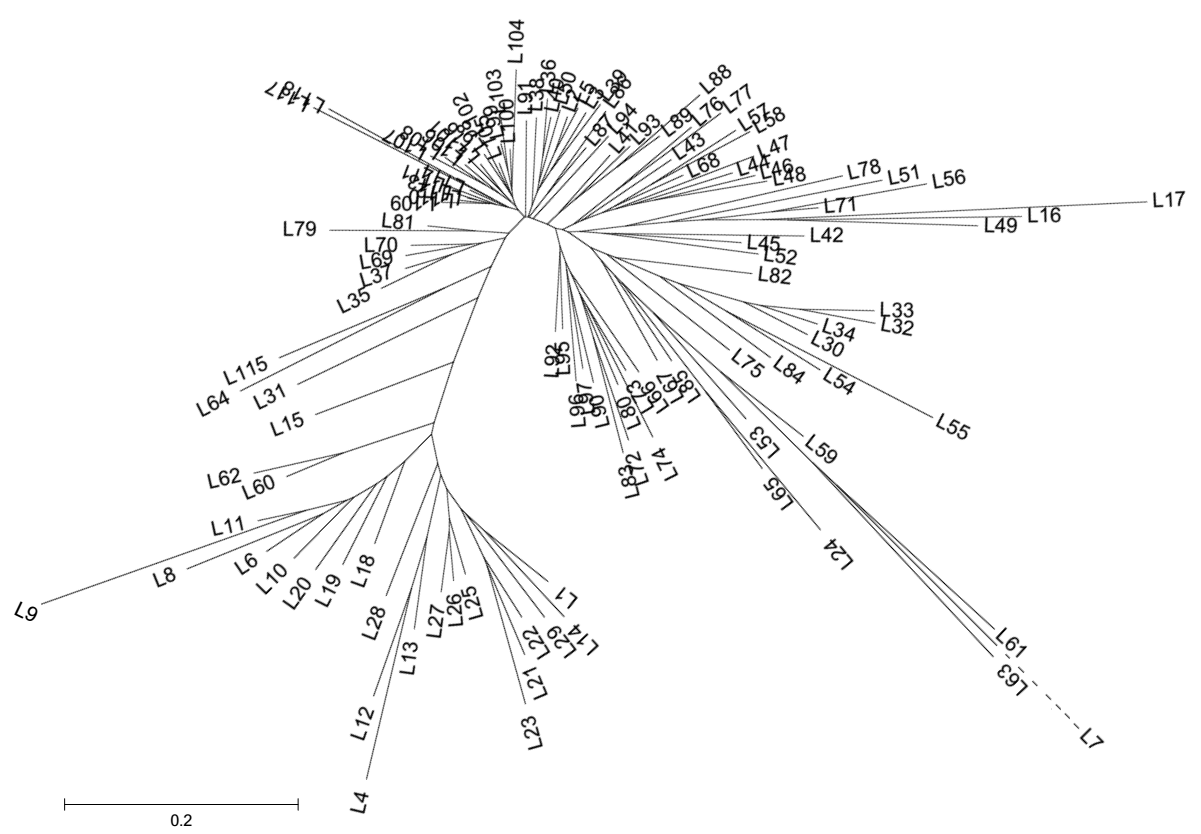 | 图1 宁麦9号及其衍生品种(系)的Neighbor-Joining遗传聚类分析品种名称详见附表。L7遗传距离较大, 以虚线表示。Fig. 1 Dendrogram of Ningmai 9 and its derivatives based on Neighbor-Joining analysis Variety names are listed in supplementary table. The genetic distance of L7 is very large and broken down with dashed line. |
185对SSR引物共扩增出490个等位变异位点, 每对引物1~7个, 平均2.6个。衍生一代各染色体相同等位变异比例为55.71%~88.29%, 衍生二代则为56.33%~83.50%, 均以4D变异比例最高。除6D外, 衍生一代各染色体相同等位变异比例均高于衍生二代, 3个基因组间等位变异比例相近(表1)。在全基因组中, 衍生一代的相同等位变异比例为75.60%, 衍生二代为67.81%。
| 表1 不同染色体上宁麦9号与其衍生品种(系)的相同等位变异比例 Table 1 Percentage of same alleles shared between Ningmai 9 and its derivatives on thechromosomes (%) |
衍生一代的KPC及FPC平均略高于衍生二代; 二者KPC都高于亲本宁麦9号, 而FPC则无明显差异; 衍生世代的KPC、FPC出现较大变异, 变异指数均超过5% (表2)。方差分析结果表明, KPC和FPC来自基因型的变异, 其F值分别为7.92 (P< 0.01)和5.31 (P< 0.01), 两性状来自年份的变异, 其F值分别为6.48 (P< 0.05)和2.33 (P> 0.05); 世代间比较, 虽然两性状都呈现衍生一代> 衍生二代> 宁麦9号的趋势(宁麦9号及其衍生一代和二代的KPC依次为10.71、11.90和11.41; FPC依次为9.64、9.96和9.93), 但KPC在不同世代间存在差异, 而FPC无显著差异。
| 表2 宁麦9号及其衍生系籽粒蛋白质含量和面粉蛋白质含量变异 Table 2 Phenotype analysis for kernel protein content and flour protein content in Ningmai 9 and its derivatives (%) |
在8条染色体上有11个标记与KPC或FPC关联(P< 0.01), 单个标记表型贡献率为5.62%~10.59% (表3)。与KPC关联的6个位点中, Xgwm539、Xgwm508及Xwmc397连续两年均被检测到; 与FPC关联的个位点中, Xgwm539及Xwmc468也连续两年被检测到, 其中Xgwm539标记同时与KPC和FPC关联。上述4个标记在全部材料中的频率分别为43.48%、54.55%、91.45%和79.49%, 各位点在衍生一代中的频率均高于在衍生二代中(表4)。除Xgwm508外的3个标记均对KPC和FPC有负向调控作用。
| 表3 籽粒蛋白质含量和面粉蛋白质含量的关联分析结果 Table 3 Association analysis for kernel protein content and flour protein content |
| 表4 籽粒蛋白质含量和面粉蛋白质含量关联位点在宁麦9号衍生世代中的分布频率 Table 4 Frequencies of markers associated with kernel protein content and flour protein content in Ningmai 9 derived generations (%) |
| 附表 供试宁麦衍生品种(系)及其编号 Table Supplementary table Varieties (lines) derived from Ningmai 9 and their codes |
宁麦9号是以江苏淮南小麦品种扬麦6号为母本与日本品种西风为父本, 采用改良集团法选育而成的小麦品种, 1997年通过江苏省农作物品种审定委员会审定, 由于其丰产性好、弱筋品质稳定, 很快成为江苏淮南麦区主推品种。同时作为优良亲本被多家育种单位利用, 自2005年以来的10年间已经育成通过国家或省品种审定的小麦品种15个, 从而成为长江中下游麦区重要亲本。骨干亲本容易出品种的原因, 除了品种自身综合性状优良外, 这些优良性状通常具有较高的一般配合力, 即易与其他材料杂交育成优良品种[16]。宁麦9号在不孕小穗数(负向效应)、穗粒数、单穗粒重、收获指数、蛋白质含量(负向效应)、碱水保持力(负向效应)、赤霉病抗性等方面都具有较高的一般配合力[9, 10, 17, 18]。从基因组的水平分析, 其拥有的优势等位变异较其他品种多, 新品种的遗传组成与骨干亲本具有更高的相似性, 而不是双亲的平均值[19]。本研究发现宁麦9号与其衍生品种间的遗传相似性为0.55~0.88, 对衍生一代和二代的贡献率分别为75.60%和67.81%, 均显著高于理论遗传贡献率, 这与胜利麦/燕大1817[20]、碧蚂4号[21]、欧柔[22]和周8425B[23]等其他骨干亲本衍生群体的报道一致, 说明宁麦9号多数遗传位点与优良农艺性状或抗性等存在关联, 由于育种家按育种目标的选择偏好导致后代中保持较高频率宁麦9号的等位变异位点。
对宁麦9号及其衍生品种(系)扩增带型进行比较分析, 在超过90%品种(系)中呈现相同带型的分子标记有33个, 对比前人研究发现这些标记与小麦籽粒产量、灌浆速率、开花期、小穗密度、穗发芽抗性、赤霉病抗性、锈病抗性等众多重要性状相关联[24, 25, 26, 27, 28, 29, 30, 31, 32], 宁麦9号可能在这些标记区段具有优良变异。本研究对宁麦9号及其衍生系的籽粒及面粉蛋白质含量连续两年测定发现, 衍生一代和衍生二代的平均蛋白质含量高于宁麦9号, 这是由于育种家在小麦育种后代选择中并未以弱筋专用小麦作为唯一品质目标, 因此, 弱筋专用小麦品种育种仍有待加强。
关联分析是以连锁不平衡为基础鉴定群体内性状与遗传标记关系的分析方法, 在小麦[33]、玉米[34]和水稻[35]上都得到了应用。在关联分析的基础上, 张学勇等[36]提出通过选择牵连效应和关联分析相结合寻找重要染色体区段的方法, 由于供试材料间存在着一定亲缘关系, 基于系谱的关联分析可能受群体遗传结构的影响, Yu等[37]发展了一种混合线性模型的关联分析方法以减少“ 伪关联” 和“ 假阴性” 的频率。本研究对小麦蛋白质含量进行关联分析, 共检测到4个相对稳定的与籽粒和面粉蛋白质含量相关联的分子标记, 除Xgwm508外, 其余标记均对籽粒和面粉蛋白质含量起负向调控作用, 即与宁麦9号低蛋白质含量相连锁, 因此在育种选择过程中, 选择带有Xgwm539、Xwmc397及Xwmc468的材料有利于获得低蛋白质含量的小麦品系。Prasad[38]在2D染色体上发现一个与蛋白质含量相关的分子标记Xwmc41, 与Xgwm539距离相近; Olmos等[39]发现Xgwm508可作为小麦高蛋白含量的选择标记, 与本研究结果一致; Xwmc397与Xwmc468则是首次发现与蛋白质含量相关。此外, 在这些标记附近区段检测到面粉吸水率[40]、面团形成时间、沉降值[41]等多个品质性状QTL, 这些性状同时关系到蛋白质质量, 可在应用中同时关注其表现。对比前人研究发现, 除Xwmc468外的其他3个标记附近区段还是产量、抗病等重要性状相关QTL的热点区域, 在Xgwm539附近区段检测到赤霉病抗性[42, 43, 44]、锈病抗性[45, 46]等性状相关QTL, 选育过程中注重这些性状的选择可能是导致其在衍生世代中频率较低的原因之一; Xgwm508与抗锈病基因Lr53、Yr35[47]紧密连锁; Xwmc397除与每穗粒重、每株穗数[48]相关联外, 还与赤霉病抗性基因Fhb2[49]连锁。因此, 为更好地在分子标记辅助选择育种中应用, 尚需采用更多标记对蛋白质含量进行精细定位以获得更加紧密的分子标记。
宁麦9号与其衍生品种间的遗传相似系数为0.55~0.88, 育种过程中受人为选择的影响, 宁麦9号对衍生一代和二代的贡献率分别为75.60%和67.81%。衍生品种(系)籽粒和面粉蛋白质含量存在较大变异, 4个稳定的籽粒和面粉蛋白质含量关联位点中, 3个与宁麦9号低蛋白质含量连锁, 有望进一步应用于优质弱筋小麦育种。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|