
{kind=link}
{kind=link}
{kind=link}
膜下滴灌水分亏缺下棉花开花后非叶绿色器官光合特性及其对产量的贡献
[占东霞, 张超, 张亚黎, 罗宏海, 勾玲, 张旺锋*  ]
]
]
|
|


第一作者联系方式: E-mail: zhandongxia@sina.cn
选用北疆棉区主栽品种新陆早33号和新陆早45号, 设置常规滴灌量(CI)、轻度水分亏缺滴灌量(SDI)、中度水分亏缺滴灌量(MDI) 3种处理, 在田间条件下研究了不同滴灌量对棉花叶片和非叶绿色器官叶绿素(Chl)含量、净光合速率( Pn)、气孔导度( gs)和光合物质累积的影响, 明确了水分亏缺下棉株非叶绿色器官光合作用对产量的贡献。结果表明, 各滴灌量条件下棉花非叶绿色器官单位面积的 Pn、Chl含量下降幅度较叶片小, 且随棉花生育进程的变化较小。在棉铃生长发育后期, 随滴灌量减少, 棉花非叶绿色器官光合物质生产能力对产量起着更为重要的作用。中度水分亏缺条件下, 棉铃(铃壳和苞叶)和茎秆光合作用对铃重的相对贡献率分别增加至16.8%~34.9%和7.6%~17.5%。因此, 采用膜下滴灌植棉技术时适当控制滴水量, 在保证叶片具有较高光合速率的同时, 发挥棉花非叶绿色器官的光合抗逆能力, 对挖掘滴灌棉花节水增产潜力具有重要意义。
Leaf is one of the main photosynthetic organs, while other green parts of plant also retain or develop chlorophyll and have photosynthesis. To better understand the whole plant photosynthesis production potential and contribution to cotton yield, we selected Xinluzao 33 and Xinluzao 45 (two common cultivars in Northern Xinjiang) with three irrigation treatments (CI, conventional irrigation; SDI, slight deficit irrigation; MDI, moderate deficit irrigation) to measure the chlorophyll content (Chl), net photosynthetic rate ( Pn), stomatal conductance ( gs) and photosynthate accumulation in leaf and non-leaf green organs during different growth stages and the contribution of non-leaf green organs to yield. The results showed that the Pn and Chl in non-leaf organs were relatively insensitive to soil moisture stress, decreasing by only a small amount between 25 and 45 days after anthesis. With reduction of water supply, the dry matter production in non-leaf green organs played more important roles in cotton yield formation. Cotton boll weight in the moderate deficit irrigation treatment decreased by 16.8% to 34.9% when the bolls (capsule walls plus bracts) were shaded and by 7.6% to 17.5% when the stalks were shaded. Hence, limiting-irrigation treatment is important to maintain high leaf photosynthetic rates. It is also important to develop the potential photosynthetic capacity of non-leaf green organs. This is especially important when leaf photosynthesis capacity declines due to leaf aging or water stress.
作物产量的90%~95%来自光合作用所形成的有机物。叶片是作物主要的光合器官, 但许多作物非叶器官或组织, 如茎秆、叶柄、叶鞘、花器和果实等, 也能形成或含有叶绿素, 并具有实际或潜在的光合能力, 对产量具有一定的贡献[1, 2, 3, 4]。小麦(Triticum aestivumL.)穗的外稃、内稃、颖片、花梗、果皮和芒等都有固定CO2的能力, 对作物产量的贡献达10%~81%[1, 5, 6, 7]; 玉米(Zea mays L.)开花后15 d, 去除苞叶, 产量下降了17.7%[8]; 水稻(Oriza sativa L.)圆锥花序的单位面积光合同化效率较对应旗叶高达30%, 但其单位叶绿素的同化速率与旗叶相差不大[9], 其光合产物对谷粒的贡献率为20%~30%[10]; 大豆(Phaseolus vulgaris L.)豆荚对外源CO2的固定速率是叶片的5%~9%[11]。研究表明, 禾谷类作物非叶绿色器官具有较强的光合抗逆性[12, 13, 14]; 在水分胁迫条件下小麦叶片光合能力的衰退速率显著高于穗、叶鞘和茎秆[15]; 小麦灌浆期土壤水分亏缺时非叶绿色器官光合能力较叶片更为稳定[16], 麦穗成为籽粒灌浆的主要光合器官[17, 18, 19], 且随灌水量的减少, 小麦旗叶部位以上非叶绿色器官对粒重的贡献率增加[20]。
棉花(Gossypium hirsutum L.)叶片是光合作用的主要器官, 但苞叶和铃壳也具有光合作用[21], 对棉铃发育的贡献一直为人们所关注[21, 22], 并对其形态特征、解剖结构[23]、生理特性[21], 以及棉铃的碳同化和碳利用等方面均有研究报道。然而有关水分亏缺条件下棉花非叶绿色器官的光合生理特性及对产量的贡献研究较少。新疆是我国最重要的优质商品棉生产基地, 棉花生产完全依赖于灌溉, 目前水资源的短缺制约了棉花综合生产力的提升, 发展节水灌溉是新疆农业经济可持续发展的必然选择。开展棉花非叶绿色器官光合抗逆性的研究, 充分了解滴灌节水条件下非叶绿色器官的光合耐逆特性, 对增强棉花整体光合能力具有重要意义。因此, 本试验在前期研究基础上, 设置不同滴灌量处理, 研究棉花各绿色器官光合能力与物质生产对水分变化的响应, 明确水分亏缺下棉花非叶绿色器官光合作用对产量的贡献, 为充分挖掘棉花整株光合生产潜力, 实现节水高产高效栽培提供理论依据。
试验于2012— 2013年在石河子大学农学试验站(45° 19° N, 86° 03° E)进行, 供试品种为北疆棉区主栽品种新陆早33号(Xinluzao 33)和新陆早45号(Xinluzao 45)。两年分别于4月18日和4月22日播种。播前深施有机肥1500 kg hm-2、氮肥240 kg hm-2、磷肥170 kg hm-2作基肥, 全生育期随水滴施氮肥45 kg hm-2。两年分别于7月4日和7月10日打顶, 化学调控6~8次, 缩节胺用量为300 g hm-2; 田间种植方式及管道铺设方法同大田膜下滴灌棉花。
设3个水分处理, 即(i)常规滴灌量(CI, conventional irrigation), 控制0~60 cm土壤含水量保持在大田正常田间持水量; (ii)轻度水分亏缺滴灌量(SDI, slight deficit irrigation), 每次灌水量为常规滴灌量的60%~70%; (iii)中度水分亏缺滴灌量(MDI, moderate deficit irrigation), 每次灌水量为常规滴灌量的30%~ 40%。试验小区面积68 m2, 裂区试验设计, 主区为水分处理, 副区为品种, 重复3次。在棉花盛蕾期进行不同滴灌量处理, 8月底停止灌溉, 滴水量用水表和球阀严格控制。2012年3个滴灌量处理总的滴水量(滴灌量+降雨量)分别为451、317和183 mm; 2013年分别为474、385和297 mm。为使土壤水分达到试验设计要求, 在棉花整个生长季节定期采用Watermark土壤水分传感器(model 200SS; Irrometer Co., Riverside, USA)监测0~60 cm土层土壤相对含水量。
取打顶后棉株主茎倒二叶及对应的主茎秆、苞叶和铃壳用于棉花气体交换参数和叶绿素含量的测定。为减少测定叶片的叶龄不同所造成的误差, 用标签纸标定每处理约1000株左右棉株倒二果枝第1果节花蕾。每次测量均在滴水后第4天进行。
1.2.1 各绿色器官气体交换参数
用Li-6400便携式光合测定系统(Li-Cor, USA)于棉株开花后5、15、25、35和45 d测定叶片、苞叶和铃壳的气体交换参数。用6400-02 LED红蓝光源叶室(2 cm × 3 cm)测定叶片和苞叶光合, 控制叶室内光强为1800 μ mol m-2s-1; 采用簇状叶室(6400-05 Conifer Chamber, Li-Cor, USA)和白光LED光源(Luxeon LEDs; Electus Distribution, NSW, Australia)测定铃壳光合速率; 各绿色器官重复测定3次。测定结束后, 精确测量苞叶和铃壳表面积并重新计算光合速率。
1.2.2 各绿色器官叶绿素(Chl)含量
采用Lichtenthaler[24]的方法, 测定棉株开花后5、15、25、35和45 d叶片、苞叶、铃壳和茎秆的Chl含量。将叶片、苞叶、铃壳用打孔器(直径为4 mm)打成小圆片, 茎秆切成长方形, 量其长宽, 放入到25 mL具塞试管中, 用80%的丙酮溶液浸提后测定。各绿色器官重复3次。
1.2.3 各绿色器官干物质积累
在棉花吐絮期, 从各水分处理分别取长势一致的棉株3株, 分别剪下叶片、苞叶、茎秆和铃壳, 将其装袋置干燥箱, 于105℃杀青30 min, 80℃烘干至恒重后称重。
1.2.4 非叶绿色器官对铃重的贡献
采用遮阴法[1]测定棉铃(铃壳和苞叶)与茎秆光合作用对铃重的相对贡献。从3个水分处理取72株棉花, 每处理分成3组, 即对照、棉铃遮阴、茎秆遮阴, 每组为8株。棉铃遮阴, 即用铝箔纸(铝箔纸上打微孔便于内外气体交换)将棉株上每个棉铃从开花后15 d后开始包裹; 茎秆遮阴即用铝箔纸将主茎秆和果枝遮阴(从打顶后开始)。于收获期将对照、棉铃遮阴、茎秆遮阴处理的每个铃的籽棉分别称干重。
相对贡献率(%)=(对照产量-遮阴产量)/对照产量× 100%。
采用SPSS 17.0 统计软件(SPSS Inc., Chicago, IL, USA)分析试验数据, 用最小显著差异法做方差分析(P< 0.05), 图表中数据均为平均值± 标准误差。
3种滴灌量处理下棉花单位面积叶片Chl含量显著高于非叶绿色器官, 两年表现一致。随着开花后时间延长, 叶片Chl含量下降最为明显。与开花后5 d相比, 开花后45 d滴灌量处理下叶片Chl含量, 2013年分别下降了60.4%~79.0%、43.7%~49.0%、49.3%~62.4% (图1-A)。不同滴灌量处理下非叶绿色器官Chl含量表现为: 苞叶和茎秆的Chl含量先增加随后轻微下降, 苞叶Chl含量在开花后15 d到达峰值, 随后下降, 且在各水分处理间差异不显著(图1-B); 铃壳的Chl含量仅次于叶片, 与开花后15 d相比, 3种水分处理下分别下降了26.5%~26.7%、24.7%~33.9%和39.2%~41.7% (图 1-C); 茎秆Chl含量随棉花生育进程变化幅度较小, 在开花后25 d达到最大值(图1-D)。品种间差异不大。
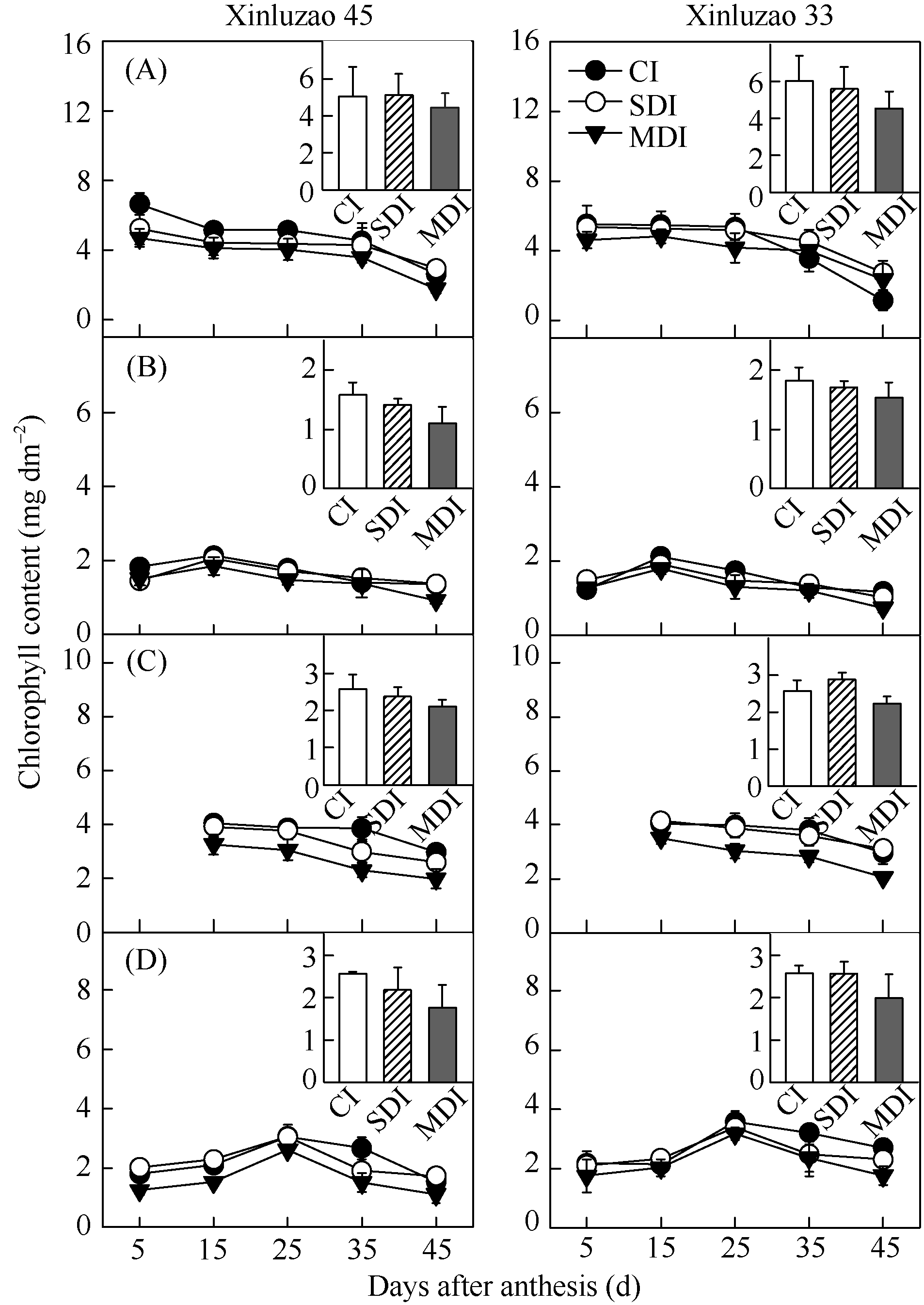 | 图1 不同滴灌量下棉花叶片Fig. 1 Change of chlorophyll contents in leaves |
2.2.1 净光合速率
随着棉铃发育, 单位面积叶片光合速率显著下降; 单位面积苞叶光合速率在开花后15 d达到最大值, 铃壳光合速率在开花后25 d达到最大值, 随后均下降(图2)。2个参试品种间光合速率的变化相差不大。各器官光合速率表现为叶片从最大值到最小值下降最快, 为54.3%~75.5%, 其次是铃壳为40.2%~68.9%, 苞叶下降幅度最小, 仅为12.1%~32.6%。至开花后45 d, 叶片光合速率为7.4~8.2 μ mol m-2 s-1, 苞叶为1.4~2.0 μ mol m-2 s-1, 铃壳为12.9~14.1 μ mol m-2 s-1。随着滴灌水量的减少, 2年试验期间叶片、苞叶和铃壳光合速率均表现为下降趋势, 但各生育时期不同绿色器官光合速率受水分影响的变化有差异。
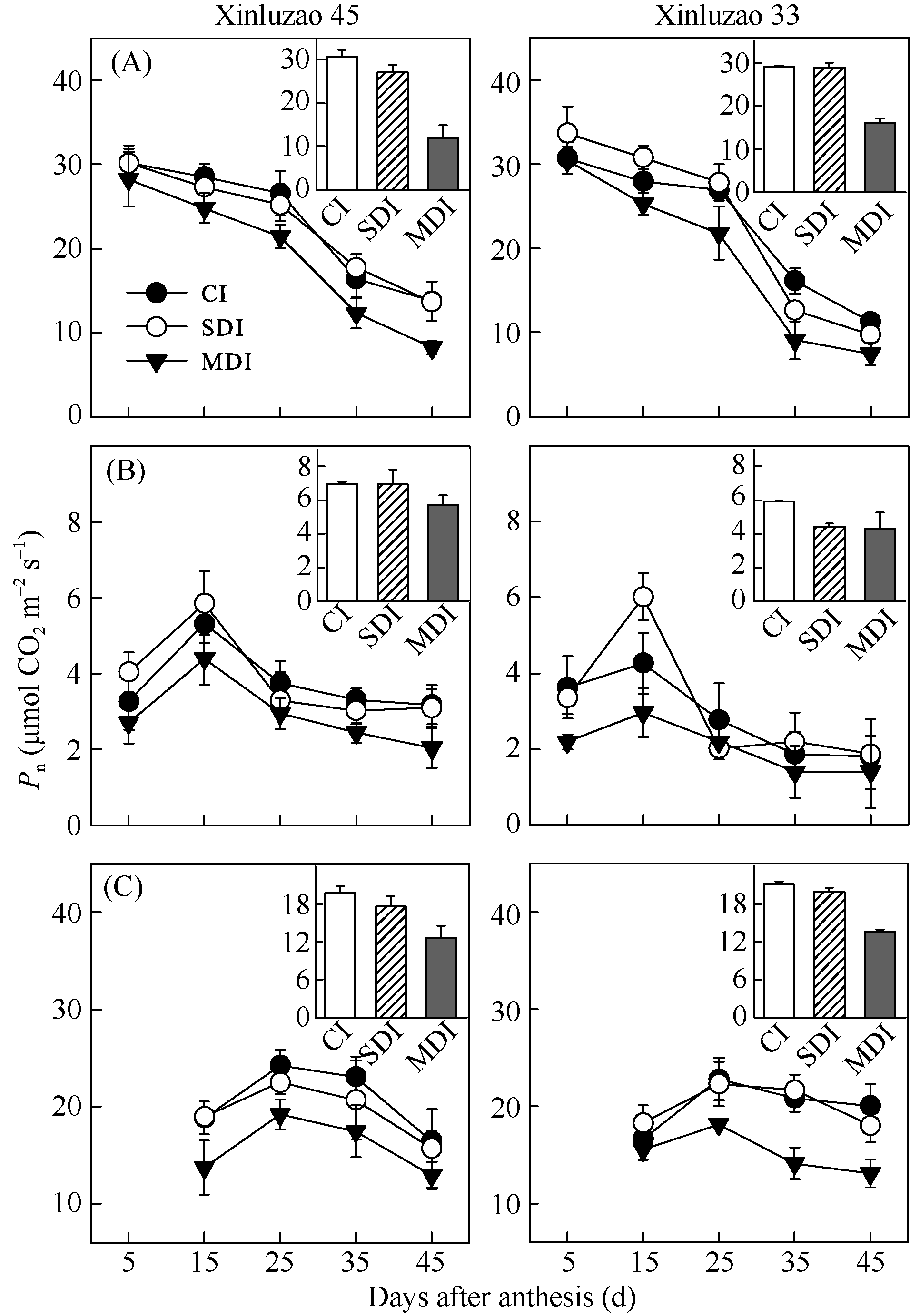 | 图2 不同滴灌量处理下棉花叶片Fig. 2 Change of photosynthetic rate in leaves |
为了便于比较, 假设各生育时期不同滴灌量处理叶片、苞叶和铃壳光合速率的和为总光合速率, 可分析盛花期和盛铃期棉花各绿色器官的净光合速率及对整株光合的贡献。常规滴灌量处理下, 盛花期苞叶和铃壳光合速率占总光合速率的42.7%~ 45.8%; 至盛铃期占总光合速率的58.5%~61.6%。轻度水分亏缺下盛花期和盛铃期苞叶与铃壳光合速率分别占总光合速率的44.1%~47.6%、57.2%~65.4%, 中度水分亏缺下分别占总光合速率的42.2%~42.3%、61.7%~63.0%, 这表明棉花苞叶和铃壳光合能力对整株的光合贡献随水分的减少和生育时期的推进而增加。
2.2.2 气孔导度
随水分供应减少, 叶片、苞叶和铃壳的气孔导度随着开花后天数的增加逐渐降低。叶片的gs最高, 其次是铃壳, 苞叶最低(表1)。常规滴灌量下, 从花后5~45 d, 叶片、苞叶和铃壳的gs分别下降了69.7%~75.9%、33.3%~50.0%和30.1%~ 40.9%; 轻度水分亏缺下分别下降了75.3%~76.4%、44.4%~50.0%和21.0%~44.4%, 中度水分亏缺下分别下降了71.1%~78.3%、33.3%~42.9%和30.0%~33.3%。
| 表1 不同滴灌量处理下棉花各绿色器官的气孔导度(2013) Table 1 Effect of water deficit on stomatal conductance (gs) in cotton organs during various times after anthesis in 2013 (mol m-2s-1) |
综合两年的棉花生物量数据分析可知(图3), 不同滴灌量处理下, 吐絮期棉花叶片生物量占地上部单株总生物量的7.7%~16.7%, 非叶绿色器官生物量占地上部单株总生物量的83.3%~92.3%, 其中棉铃生物量占地上部单株总生物量的61.7%~75.7%, 茎秆生物量占地上部单株总生物量的15.9%~23.4%。中度水分亏缺下的叶片生物量占地上部单株总生物量的比例低于其他2个处理。
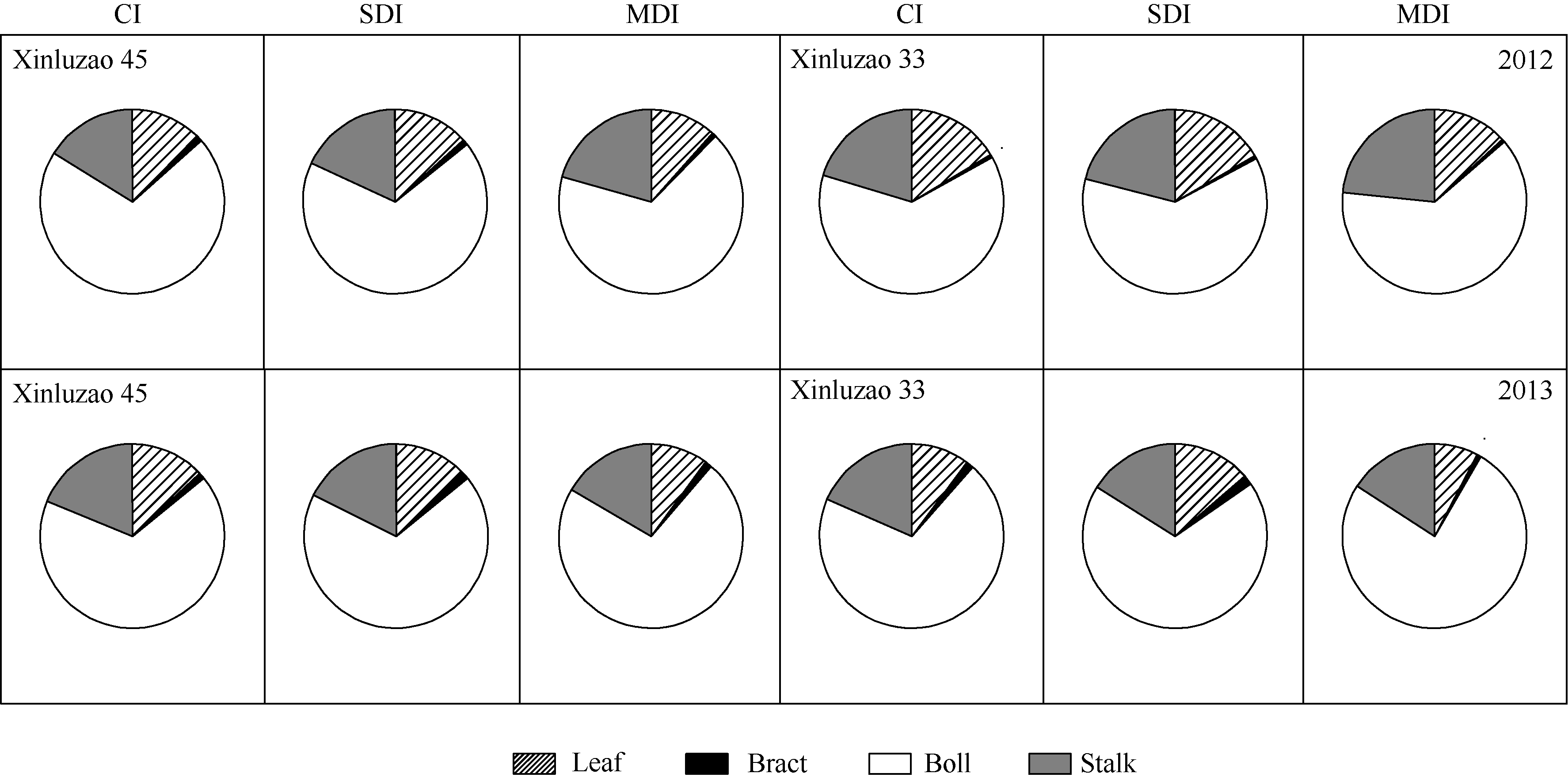 | 图3 吐絮期不同滴灌量处理棉花各绿色器官生物量Fig. 3 Biomass of each green organ in cotton as affected by different water supplies at boll open stage |
通过对棉花非叶绿色器官的遮阴处理, 测得棉铃(铃壳+苞叶)和茎秆对籽棉重的相对贡献(表2)。表明常规滴灌量处理下, 棉铃光合作用对籽棉重的相对贡献率为13.4%~29.7%, 茎秆为6.0%~9.9%。中度水分亏缺下, 棉铃光合作用对籽棉重的相对贡献率增加至16.8%~34.9%, 茎秆增加至7.6%~17.5%。中度水分亏缺下, 棉花非叶绿色器官光合作用对铃重总的贡献为31.1%~49.4%。
| 表2 不同滴灌量下棉花非叶绿色器官光合能力对铃重的相对贡献率 Table 2 Contribution of non-leaf organs to boll weight in cotton as affected by different water supplies (%) |
对产量有一定的贡献[21, 22, 25]。本研究结果表明, 在棉花生长发育后期, 非叶绿色器官单位面积的Chl含量棉花苞叶和铃壳含有叶绿素, 具有光合潜力, 下降幅度较叶片小, 苞叶和铃壳的叶绿素含量均保持相对稳定, 对其保持相对稳定的光合速率奠定了基础。棉花各非叶绿色器官单位鲜重Chl含量均显著低于主茎叶片, 且随生育进程, 下降幅度亦显著低于叶片[4]。叶片Chl含量下降幅度最大, 是其迅速衰老的体现(图1)。
3种滴灌量条件下, 开花后25 d单位面积铃壳光合速率明显高于叶片。本课题组前期研究发现, 在棉铃发育的后期, 铃壳单位面积光合放氧能力较叶片明显高, 开花后20 d左右铃壳单位面积氮含量显著高于叶片[4], 较高的光合速率可能与其氮含量有关[26]; 铃壳的蜡质层较叶片和苞叶厚[27], 具有较高的持水能力(数据未列出), 相对含水量高于叶片[28], 这可能是铃壳光合速率高的另一原因。3种滴灌量处理下, 单位面积苞叶的光合能力显著低于叶片。由于苞叶总的气孔密度是叶片的一半[29], 导致苞叶的gs较低(表1), 较低的光合能力可能是较低气孔密度所致[30]。另外, 苞叶的单位面积Chl含量也明显低于叶片(图1)。3种滴灌量处理下, 盛花期至盛铃期苞叶和铃壳光合速率占总光合速率的比例显著增加。另外随供水量的减少, 盛铃后期棉花苞叶、铃壳和茎秆的光合面积占整株总面积的比例增加至43%[31], 棉铃发育后期光合功能持续期延长。总之, 这些结果均表明苞叶和铃壳对棉花产量有重要的作用, 尤其是在生长发育后期和水分亏缺条件下表现更为突出。
水分亏缺下棉花苞叶和铃壳净光合速率下降很小, 而叶片光合速率下降幅度较大, 表明非叶绿色器官对水分逆境的适应能力较强。棉花苞叶的绿色表面积较小, 而且苞叶包裹着棉铃, 仅单面受光, 与叶片相比, 虽然光合速率较低, 但在水分亏缺下光合速率降低不明显。因此, 在水分亏缺逆境下非叶绿色器官的光合抗逆性强[29], 在后期能维持较稳定的光合速率, 有利于棉铃光合产物的累积。
本课题组前期研究表明, 棉花苞叶生长在高浓度CO2环境下[32], 尽管棉铃呼吸流失了大量的CO2, 但棉花非叶绿色器官具有循环利用内源CO2的能力, 对产量具有极其重要的作用[2], 棉铃的呼吸为铃壳的光合作用提供了一个重要的碳源[21]。前人研究表明, 油菜(Brassica napus L.)角果皮是发育后期光合作用的重要器官, 角果是库也是重要的源[33], 对籽粒的贡献达70%, 是产量的重要组成部分[34]; 角果皮具有较强的光合抗逆性, 表现在较低的气孔导度和维持较高的相对含水量上[35]。棉花铃壳直接相连并包裹着种子和纤维, 意味着由此光合器官产生的光合产物运输至果实距离短[4], 且铃壳有效光合作用时间也应是果实形成的最关键时期。随滴灌量减少, 棉花非叶绿色器官光合物质生产能力对产量起着更为重要的作用(图3)。棉花茎秆[36]、苞叶、铃的这种光合作用是对叶片光合能力不足的一种补充。
棉花各绿色器官光合作用对产量的相对贡献可以通过遮阴法来估算[1]。本研究表明, 水分亏缺下铃壳、苞叶和茎秆光合作用对铃重的贡献高达40%~ 50%; 当减少滴灌水量时, 棉铃和茎秆对籽棉重的相对贡献率增加。有研究表明, 谷类作物穗光合对粒重的贡献约为10%~76%[37]。因此, 在目前膜下滴灌生产条件下, 适当减少灌水量, 充分利用新疆丰富的光照资源, 发挥棉花各非叶绿色器官光合耐逆潜力, 增强生育后期非叶绿色器官的光合能力, 延长功能期, 对于补偿生育后期叶片光合能力不足, 增加产量和提高水分利用效率具有促进作用。
棉花生长后期, 随着水分亏缺程度的增加, 棉铃(铃壳和苞叶)和茎秆对铃重的相对贡献率增大。在生育后期叶片开始衰老时, 其光合能力迅速下降, 非叶绿色器官的光合作用对棉花产量的贡献增加, 尤其在水分亏缺条件下非叶绿色器官的光合功能期延长, 对棉铃的贡献增加。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|