
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
四倍体小麦地方品种矮蓝麦矮秆性状的遗传分析
[周强1, 2, 3 , 袁中伟1 , 张连全1 , 甯顺腙1 , 任勇1, 2, 3 , 陶军1, 2, 3 , 李生荣2, 3 , 刘登才1, *  ]
]
]
|
|


第一作者联系方式: E-mail: zqmy0000@163.com
四倍体圆锥小麦( Triticum turgidum L. ssp. turgidum)地方品种矮蓝麦是我国重要的小麦矮秆基因资源, 经鉴定其矮秆特性对外源赤霉酸敏感。2012年配制矮蓝麦与2个高秆圆锥小麦的正反交组合, 2012—2013年在四川绵阳分别种植F1、F2代和F2:3家系, 对株高的遗传分析表明, 矮蓝麦的矮秆性状受1对隐性基因控制。利用BSA法构建高秆和矮秆池筛选多态性SSR标记, 并对矮蓝麦/青稞麦F2分离群体进行连锁分析, 将目标基因定位于7AS染色体上, 与标记 GWM471的遗传距离为2.5 cM。矮蓝麦与矮秆番麦正反交的F1和F2群体表现非常相似的株高变异特征, 初步推测矮蓝麦的矮秆基因是 Rht22; 进一步用高通量SNP和DArT标记对两品种进行全基因组扫描, 发现二者的遗传相似性高达98.7%~99.3%。因此认为, 历史上矮蓝麦和矮秆番麦可能是同一品种, 是通过人为交流而传播到不同地方。矮蓝麦携带的矮秆基因在人工合成六倍体小麦遗传背景中降低株高能力中等或较弱, 在育种中需要聚合其他矮秆基因而被利用。
Ailanmai is an important Triticum turgidum ssp. turgidum landrace carrying dwarf gene in China. Its dwarfing trait was found to be sensitive to gibberellic acid. In 2012, we crossed Ailanmai with two high plant landraces, Qinkemai and Ganmai, and obtained their reciprocal F1 hybrids. The genetic analysis was carried out in Mianyang, Sichuan Province using the F1, F2, and F2:3 populations during the 2012-2013 crop seasons. One recessive gene was proved to control the dwarfing trait in Ailanmai. Polymorphic simple sequence repeat (SSR) primers associated with plant height were selected through bulked segregant analysis (BSA) and used to identify the F2 individuals. The results indicated that the dwarf gene was located on the short arm of chromosome 7A with a genetic distance of 2.5 cM from marker GWM471. We speculated Rht22to be the dwarf gene in Ailanmai because the reciprocal F1 and F2 hybrids between Ailanmai and Aiganfanmai (carrying Rht22) exhibited similar distributions in plant height. This speculation was validated with high-through molecular marker analysis. The percentages of identical SNP and DArT markers between Ailanmai and Aiganfanmai were as high as 98.7% and 99.3%, respectively. We conclude that the two landraces might be the same variety a long time ago and became synonymic during their spread accompanying with humanity activities. The dwarf gene in Ailanmai had a moderate or weak effect to reduce plant height in synthetic hexaploid wheat. Thus, it should be utilized by pyramiding other dwarfing genes in wheat dwarfing breeding.
矮秆和半矮秆基因(Rht)的发掘和利用引发了著名的“ 绿色革命” [1, 2], 一系列矮秆和半矮秆小麦品种的育成和推广对小麦产量的提高起了关键作用, 使得在20世纪60-80年代全世界小麦产量每年递增3.4%[3, 4]。目前, 已正式命名的小麦矮秆基因有35个[5], 利用最广泛的矮源是来自日本的农林10号(Norin 10, 含Rht1和Rht2)和赤小麦(Akakomugi, 含Rht8和Rht9) [4, 6, 7, 8]。根据对赤霉酸(GA3)反应敏感程度, 将小麦矮秆基因分为赤霉酸反应不敏感型(Rht-B1、Rht-D1等)和赤霉酸反应敏感型(Rht4、Rht5、Rht6、Rht22等) [5, 9, 10]。多数矮秆基因来源于六倍体普通小麦(Triticum aestivum L., 2n=6x=42), 而Rht14、Rht15、Rht16、Rht18、Rht19、Rht22等来源于四倍体小麦(T. turgidumL., 2n=28), 且多数是通过突变获得的。目前, 四倍体小麦矮秆基因还很少应用于普通小麦育种。四倍体小麦中的优良基因可以用四倍体小麦与普通小麦杂交而被直接利用, 另外还可以通过四倍体小麦与节节麦(Aegilops tauschiiCosson, 2n=14)杂交并经染色体加倍, 形成人工合成六倍体小麦[11], 再以此为“ 桥梁” 和普通小麦杂交间接加以利用[11, 12, 13, 14, 15, 16]。
四倍体圆锥小麦(T. turgidumssp. turgidum)地方品种矮蓝麦在抽穗前后茎叶上蜡质多, 呈蓝青色, 并且植株较矮, 因而得名[17]。矮蓝麦原产于我国四川遂宁一带, 是四川古老的一个裸粒栽培四倍体小麦品种, 具有矮秆、多花多实和适应性强等特点。一般圆锥小麦的株高为120~180 cm, 而矮蓝麦的株高可低到80 cm左右, 是圆锥小麦中稀有的类型[18]。但是矮蓝麦在不同地区种植, 株高存在差异。例如, 在四川麦区成都平原种植, 其株高常年在100~115 cm之间, 但仍是所种植的圆锥小麦里最矮的。据田良才[18]报道, 矮蓝麦具有2对隐性矮秆基因, 与农林10号、大拇指矮等普通小麦的矮秆基因不同, 而且对赤霉酸反应敏感, 是小麦属中一个新的矮秆基因资源。为加速矮蓝麦矮秆基因的育种利用, 本研究对矮蓝麦的矮秆性状进行了遗传分析, 为进一步定位和利用该基因提供依据, 同时分析该基因在人工合成小麦中的遗传效应, 以助于将矮蓝麦矮源用于普通小麦矮化育种。
圆锥小麦(T. turgidumL. ssp. turgidum, 2n=4x=28, AABB)矮秆地方品种矮蓝麦(保存号为AS313)、高秆地方品种甘麦和青稞麦、圆锥小麦AS2255, 硬粒小麦Langdon (T. turgidumL. ssp. durum, 2n=4x=28, AABB), 节节麦(Ae. tauschii, 2n=2x=14, DD)材料AS60和AS77, 以及四倍体小麦和节节麦杂交并染色体自然加倍的人工合成六倍体小麦(2n=6x=42, AABBDD)材料Syn-SAU-1 (AS2255/ AS60)、Syn-SAU-2 (AS313/AS60)、Syn-SAU-5 (Langdon/ AS60)、Syn-SAU-16 (AS313/AS77)[15], 均由四川农业大学小麦研究所提供。圆锥小麦地方品种矮秆番麦(T. turgidumL., 2n=4x=28, AABB)由西华师范大学生命科学学院提供。
参考郭保宏[19]的描述进行赤霉酸处理, 方法略有改动。随机选取300~400粒矮蓝麦种子, 用0.1%氯化汞消毒3~6 min, 蒸馏水冲洗, 放在直径9 cm的培养皿中, 加入适量蒸馏水, 室温浸种; 将萌动的种子在4℃冰箱内放置24 h, 以确保发芽整齐; 然后将发芽的种子放入培养皿, 每皿50粒, 3次重复, 加入50 mg L-1赤霉酸溶液(处理)或蒸馏水(对照), 在20~22℃下暗箱培养, 每天补充10 mL赤霉酸溶液或蒸馏水。出苗7~10 d, 待第一叶长基本稳定之后, 测定第一叶长和胚芽鞘长度。用t测验法检验处理与对照的差异。
2012年春在四川农业大学温江校区试验地, 配制矮蓝麦/青稞麦、青稞麦/矮蓝麦、矮蓝麦/甘麦、甘麦/矮蓝麦的正反交, 并于2012年夏在四川省阿坝州马尔康县夏繁加代(单粒播种)。2012年秋季, 将亲本、F1、F2种于四川省绵阳市农业科学研究院试验地, 行长2 m、行距26.5 cm、株距10 cm, 亲本、F1代各播3行、F2代播30~60行, 于2013年春小麦抽穗后逐株调查单株株高(从主茎基部量至穗顶, 不包括芒长)。2013年秋在四川省绵阳市农业科学研究院种植杂交组合矮蓝麦/甘麦的661个F2:3家系, 2014年春逐株调查株高及其分离情况。
2013年春季, 选择矮蓝麦/青稞麦F2群体, 采用2× CTAB法分单株提取基因组DNA [20]。根据已公布的小麦全基因组上的SSR引物, 随机选取均匀分布于A、B染色体上的SSR引物202对, 由生工生物工程(上海)有限公司合成。参照Michelmore等[21]提出的集群分离分析法(BSA), 从F2群体中随机选取高秆和矮秆单株各10个, 等量混合基因组DNA, 构建高秆池和矮秆池。用SSR引物对矮蓝麦和青稞麦进行多态性分析, 筛选在双亲间存在差异的引物; 用差异引物在高、矮秆池中进行PCR扩增, 找到差异引物对F2群体单株进行PCR扩增, 参照Gill等[22]报道的PCR反应体系和程序。PCR产物经8%变性聚丙烯酰胺凝胶电泳分离, 银染显影后按单株记录带型。应用JoinMap4.0软件绘制基因连锁图。
采用2× CTAB法提取圆锥小麦(矮蓝麦、矮秆番麦、AS2255)、硬粒小麦Langdon的叶片基因组DNA, 纯化后送Triticarte公司(Canberra, Australia; http://www.Triticarte. com.au/)进行DArT和SNP标记全基因组扫描, 选取P≥ 80%的可靠标记进行数据分析。利用NTSYSpc version 2.10e软件进行遗传相似系数分析和聚类分析。
2014年春在四川农业大学温江试验地, 配制矮蓝麦与矮秆番麦的正反交组合, 2014年夏将杂交后代在四川省阿坝州马尔康县夏繁加代。2014年秋季, 将亲本、F1、F2同时种植于四川省绵阳市农业科学研究院试验地, 单株单粒播种, 亲本、F1代各播3行, F2代各播16~60行, 2015年抽穗后分单株调查株高。
2012年和2013年在绵阳, 2014年在温江分别种植人工合成六倍体小麦材料及矮蓝麦、圆锥小麦AS2255、硬粒小麦Langdon, 及节节麦亲本AS60和AS77, 亲本及各材料分别播3行, 3次重复, 抽穗后调查株高。
经赤霉酸溶液处理的幼苗纤细披垂、叶色发白, 而用蒸馏水处理的幼苗直立, 叶色为正常绿色。胚芽鞘长度与幼苗第1叶长在赤霉酸和蒸馏水处理间差异显著(表1), 可见矮蓝麦为赤霉酸敏感型。
| 表1 赤霉酸对矮蓝麦幼苗叶长及胚芽鞘长度的影响 Table 1 Effect of gibberellic acid (GA3) on seedling leaves and coleoptile lengths of Ailanmai |
2.2.1 不同亲本及杂交F1代的株高构成
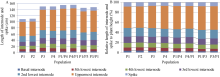
株高由穗长与各节间长度共同构成, 矮蓝麦株高显著低于高秆品种甘麦和青稞麦; 除了基部节间长度外, 矮秆品种矮蓝麦与矮秆番麦的穗长、穗下节间长、倒2节间长、倒3节间长、倒4节间长、倒5节间长均比甘麦和青稞麦短; 2个矮秆品种的株高、穗长及各节间长差异不显著(图1-A)。比较株高构成比例, 矮秆品种、高秆品种及其杂交F1代十分接近, 穗下节是株高最主要的构成因素(图1-B)。
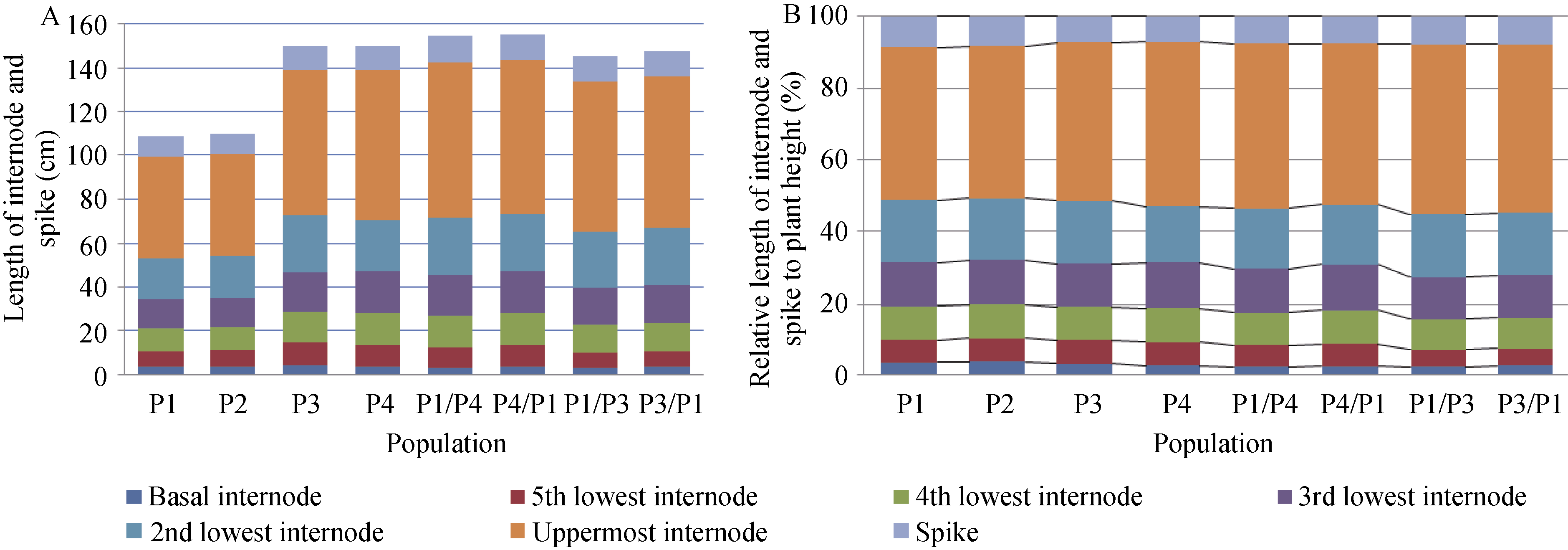 | 图1 高、矮秆亲本及其杂种F1的株高组成Fig. 1 Plant height compositions |
2.2.2 矮蓝麦矮秆性状的遗传分析
矮蓝麦与青稞麦正反交F1代的株高都偏向于高秆亲本(图1-A), 暗示矮蓝麦的矮秆性状受隐性核基因控制。矮蓝麦与青稞麦正反交F2群体中, 超过和低于127.5 cm的单株数分别为803株和277株(图2-A), 经χ 2测验, 符合3∶ 1的孟德尔分离比(χ 2=0.21, χ 20.05, 1=3.84); 同样, 矮蓝麦与甘麦杂交的F2群体中, 超过和低于122.5 cm的单株分别为839株和327株(图2-B), 也符合3∶ 1 (χ 2=5.60, χ 20.01, 1=6.63)。进一步调查矮蓝麦与甘麦杂交后代661个F2:3家系的株高, 结果F2为高株、F3发生高矮分离的有322个家系, 而F3代未发生分离的家系中高秆和矮秆的分别有155个和184个, 经χ 2测验, 该分离符合1∶ 2∶ 1 (χ 2=2.982, χ 20.05, 2=5.99)。以上证据表明, 矮蓝麦的矮秆性状由1对隐性基因控制。
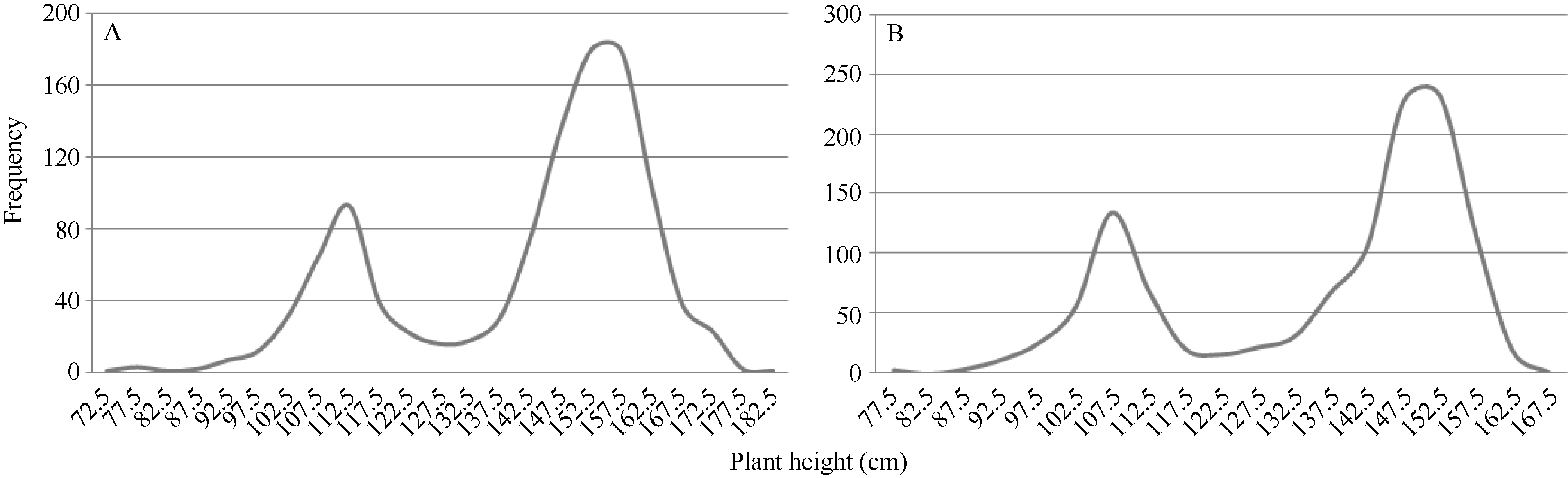 | 图2 矮蓝麦与青稞麦、甘麦杂交F2代的株高分布频率 Fig. 2 Frequency distributions of the F2 populations derived from the crosses between Ailanmai and Qingkemai or Ganmai |
2.2.3 遗传连锁图
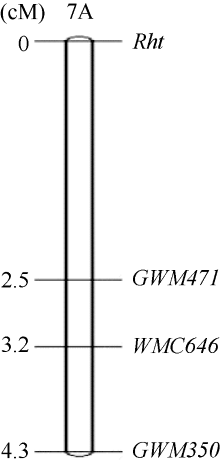
从202对SSR引物中筛选出44个在双亲间表现多态性的标记, 其中只有GWM350、GWM471和WMC646在高秆、矮秆池间呈多态性。这3对引物都位于7AS, 所以判定矮蓝麦的矮秆基因位于7A染色体短臂上。用这3对SSR引物对矮蓝麦× 青稞麦F2群体160个单株进行PCR扩增, 绘制了遗传连锁图(图3)。在该连锁图中, 距目标矮秆基因最近的标记是GWM471, 遗传距离为2.5 cM。
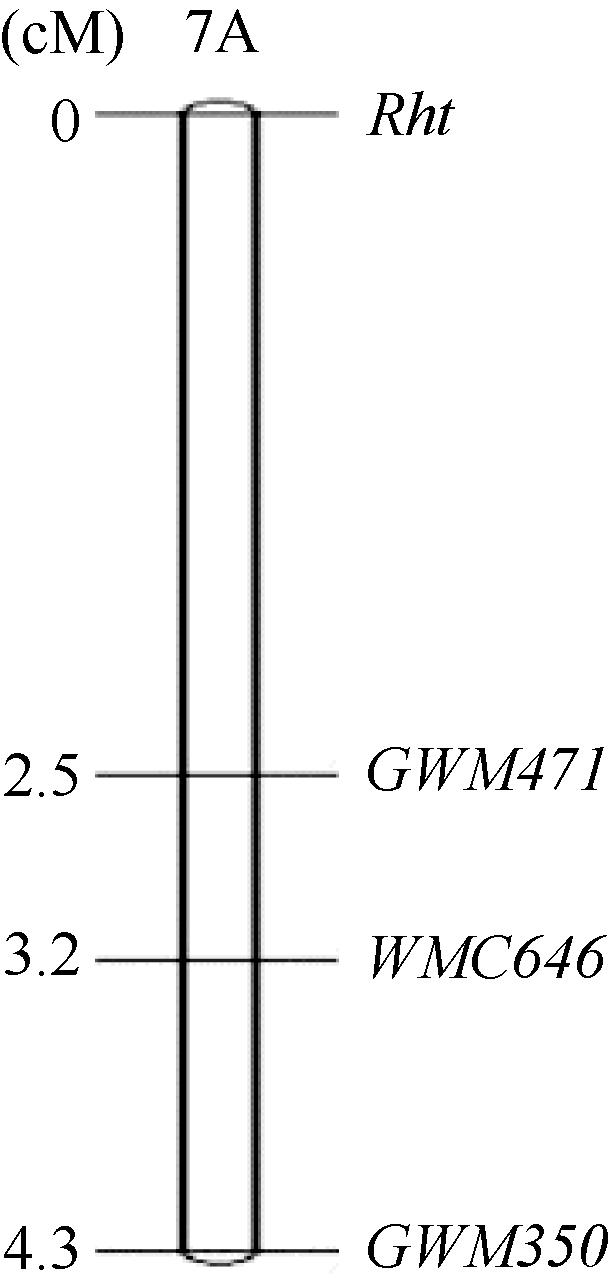 | 图3 矮蓝麦矮秆基因的遗传连锁图Fig. 3 Genetic linkage map of the dwarfing gene in Ailanmai |
SNP和DArT标记全基因组扫描结果表明, 矮蓝麦与矮秆番麦具有相同SNP标记的比例为98.9%~99.8%, 在11 260个SNP标记中, 两品种具有11 179个相同标记, 遗传相似性高达99.3%; 矮蓝麦与矮秆番麦具有相同DArT标记的比例为97.5%~99.3%, 在13 591个DArT标记中有13 410个为相同标记, 遗传相似性达到98.7%。
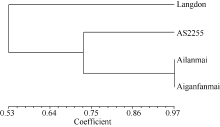
根据四倍体圆锥小麦(矮蓝麦、矮秆番麦、AS2255)和硬粒小麦Langdon的全基因组DArT标记扫描结果, 选取位于A、B基因组14条染色体上P值大于80%的4588个标记进行遗传相似系数分析和聚类分析。结果, 矮蓝麦与矮秆番麦的遗传相似系数为0.97, 大于它们与同为四倍体圆锥小麦地方品种AS2255的遗传相似系数(约0.72), 远大于它们与四倍体硬粒小麦Langdon的遗传相似系数(约0.53)。矮蓝麦与矮秆番麦的遗传相似性极高(图4)。
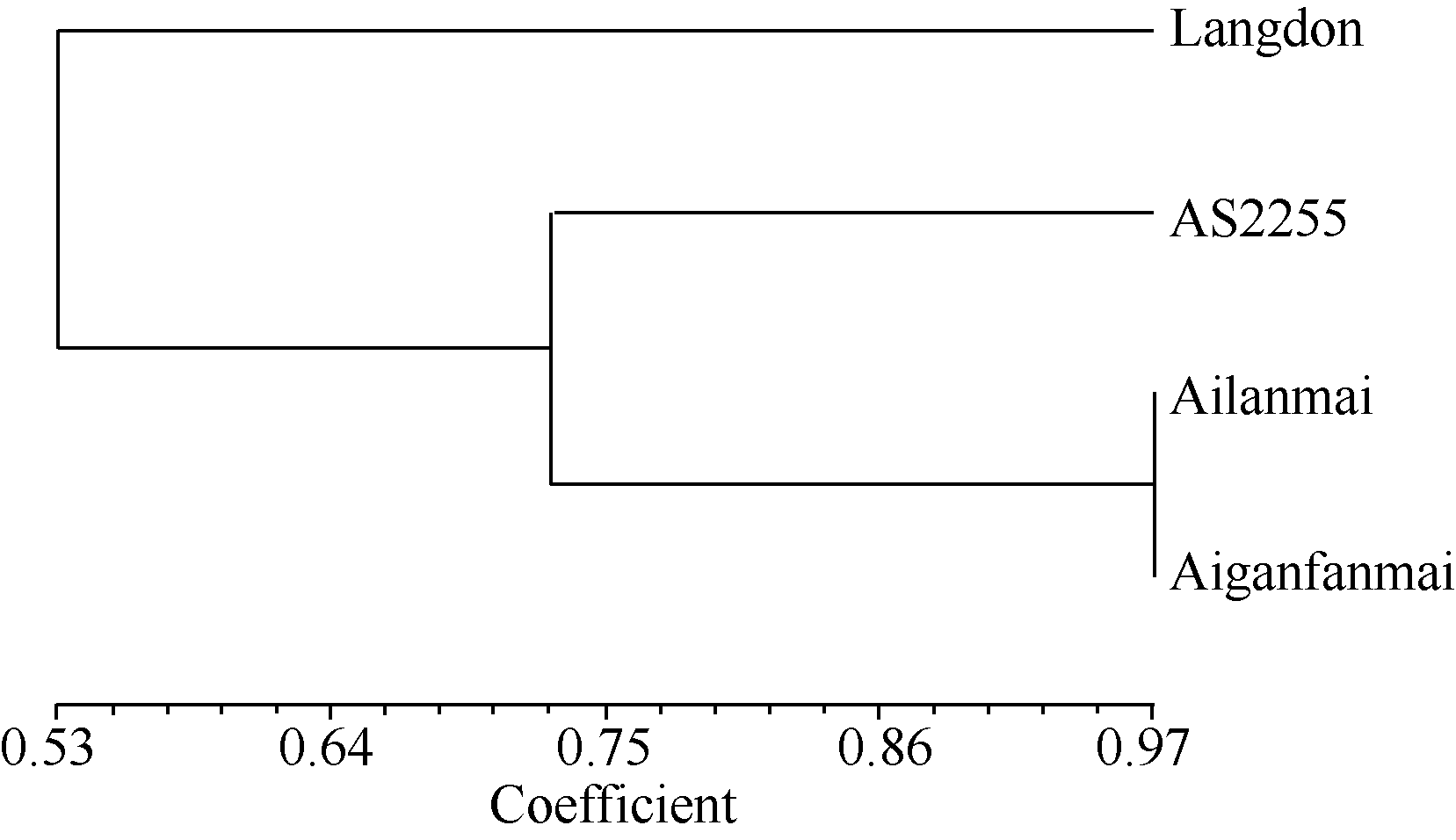 | 图4 四倍体小麦材料的DArT分析聚类图Fig. 4 Clustering of tetraploid wheat lines based on DArT analysis |
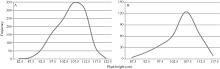
矮蓝麦平均株高为103.7 cm (88~115 cm, n = 35), 矮秆番麦平均株高为106.8 cm (85~115 cm, n = 33); 正交(矮蓝麦/矮秆番麦) F1代杂种株高为105.1 cm (95~112 cm, n = 19), 反交(矮秆番麦/矮蓝麦)F1代杂种株高106.7 cm (98~111 cm, n = 11), 都与两亲本接近; 正交F2群体1194个单株, 其株高变异为86~120 cm, 平均106.9 cm (图5-A), 反交F2群体317个单株的株高变异幅度为87~118 cm, 平均106.7 cm (图5-B)。说明正、反交F2群体的株高分布特征非常相似, 并与亲本和F1接近; 都没有出现高于120.0 cm的单株, 表明控制矮蓝麦与矮秆番麦株高的基因可能相同。
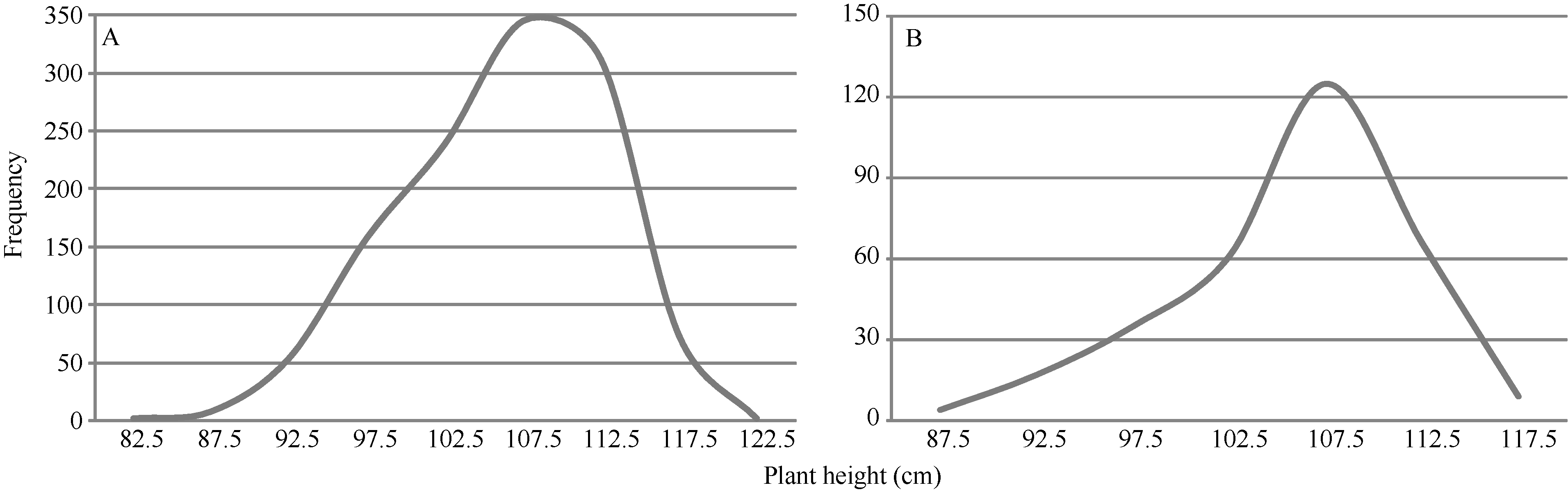 | 图5 矮蓝麦/矮秆番麦(A)和矮秆番麦/矮蓝麦(B)杂交F2代的株高频次分布Fig. 5 Frequency distributions of the F2 hybrids derived from Ailanmai/Aiganfanmai (A) and Aiganfanmai/Ailanmai (B) |
人工合成六倍体小麦的株高比亲本高, 存在超高亲优势(表2), 对其中Syn-SAU-1和Syn-SAU-2进行了3年重复试验, 且它们具有相同的节节麦父本AS60。2012、2013和2014年表型鉴定发现, 矮蓝麦株高比圆锥小麦AS2255分别低13.7、9.5和3.4 cm, 相应的人工合成六倍体小麦Syn-SAU-2比Syn-SAU-1低9.4、11.4和3.3 cm (表2), 可能与矮蓝麦的矮秆基因有关。2014年, 硬粒小麦Langdon的株高比矮蓝麦高25.4 cm, 以Langdon产生的人工合成六倍体小麦Syn-SAU-5比以矮蓝麦产生的人工合成六倍体小麦Syn-SAU-2的植株高10.6 cm, 进一步表明矮蓝麦的矮秆基因对六倍体小麦有降低株高的作用(表2)。但是, Syn-SAU-2和Syn-SAU-16都是以矮蓝麦为母本人工合成的六倍体小麦, 而其株高有明显差异, Syn-SAU-16更矮一些, 表明父本节节麦的遗传背景对人工合成六倍体小麦株高有影响。
| 表2 人工合成六倍体小麦的株高 Table 2 Plant height for synthetic hexaploid wheat |
小麦矮秆品种在外源赤霉酸处理后, 有的株高无明显变化, 有的则表现株高增加, 根据对外源赤霉酸的反应, 将小麦矮秆基因分为赤霉酸不敏感型和赤霉酸敏感型[19, 23]。本研究通过测量赤霉酸处理前后的幼苗高度和胚芽鞘长度, 证实矮蓝麦为赤霉酸敏感型小麦。我们利用4个高秆与矮秆圆锥小麦的正、反交杂交组合进行遗传分析, 确定矮蓝麦的矮秆性状受1对隐性基因控制, 与田良才[18]的试验结果不同, 是否与杂交亲本的遗传背景有关需进一步验证, 同时分离群体大小也直接影响结论的可靠性。本研究在遗传分析的基础上, 又利用SSR标记构建连锁图谱, 将矮蓝麦矮秆基因定位于7AS染色体上, 与GWM471相距2.5 cM。由于本研究采用的分离群体偏小, 未能更加准确地定位该矮秆基因。今后, 应加大作图群体, 以便获得更近的分子标记, 来精细定位目标基因。
矮蓝麦的株高、穗长、各节间长等株高构成因素都比高秆品种的对应部位低, 但是各构成因素占整个株高的比例与高秆品种相似, 这与水稻上的dn型矮秆相似[24]。矮蓝麦的株高比高秆品种青稞麦和甘麦约低27%, 表明矮蓝麦矮秆基因的降秆作用较大。已报道的矮秆基因中, 属于强降秆基因的有Rht-Dle、Rht-D1c、Rht5、Rht-Blc、Rht12等, 其降低株高幅度分别为69%、70%、55%、50%和45%[25, 26, 27, 28, 29, 30, 31, 32, 33]; 属于中等降秆强度的基因有Rht-B1b(19.0%~24.6%)、Rht-D1b(20.0%~30.4%)、Rht4(17.0%)和Rht13 (16.5%); 属于弱降秆强度的基因有Rht-B1d(11%)、Rht8(7%)和Rht9(7%)。目前小麦生产上利用最多的还是中等和弱降秆强度的矮秆基因, 如Rht-B1b、Rht-D1b和Rht8。我们通过分析人工合成六倍体小麦的株高, 发现在六倍体遗传背景下矮蓝麦所含矮秆基因具有中等或弱的降秆强度。利用矮蓝麦创制的人工合成小麦, 其株高仍高于生产上对栽培普通小麦的要求(< 100 cm)。因此, 在育种中可能需要聚合普通小麦中其他矮秆基因。矮蓝麦的矮秆基因在实际育种中的价值, 仍需进一步评价。矮秆番麦是四倍体小麦地方品种属于圆锥小麦, 原种植于陕西省南郑县, 株高80~105 cm[34], 为赤霉素敏感型[10], 其半矮秆性状由1对隐性基因控制, 该基因定位于7A染色体短臂上, 并命名为Rht22, 与SSR标记Xgwm471、WMC497和Xgwm350连锁[35]。本研究表明, 矮蓝麦也是四倍体圆锥小麦地方品种, 其矮秆基因也定位在7A染色体短臂上的类似区域; 进一步根据矮蓝麦与矮秆番麦杂种的等位测验结果, 我们认为矮蓝麦的矮秆基因是Rht22。这一推测被高通量分子标记检测结果证实, 矮蓝麦与矮秆番麦的遗传背景高度相似。因此, 我们推测矮蓝麦与矮秆番麦历史上是同一份材料, 在地方品种的传播过程中, 使用了不同的名字。至于两者的名称差异, 可能与该品种的传播有关。据《汉语词典》, 古代皇帝利用“ 番” 来称谓外国的或外族的, 矮蓝麦原产于我国四川遂宁一带, 而矮秆番麦原种植于陕西省南郑县, 可能由于人口流动而将矮蓝麦从四川带到陕西, 被称为番麦, 而后取名“ 矮秆番麦” 。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|