
{kind=link}
{kind=link}
{kind=link}
河南省近年审定小麦品种基于系谱和SNP标记的遗传多样性分析
[曹廷杰1, 2  , 谢菁忠
, 谢菁忠1 , 吴秋红1 , 陈永兴1 , 王振忠1 , 赵虹2 , 王西成2 , 詹克慧3 , 徐如强4 , 王际睿5 , 罗明成6 , 刘志勇1, * ]
, 谢菁忠]
|
|


第一作者联系方式: E-mail: caotingjie893@163.com
为了解河南省近年小麦品种的遗传基础, 利用Illumina 90k iSelect SNP标记技术对豫麦34及该省2000—2013年审定的小麦品种共96个进行全基因组扫描, 分析了其遗传多样性和遗传基础。结果表明, 在所有SNP位点中, 多态性比率为47.39% (38 661/81 587), 多态性标记在基因组间分布呈现B>A>D。96个品种亲缘关系较近, 两两遗传相似系数的平均值为0.719, 变幅为0.552~0.998, 且94.3%的品种间遗传相似系数在0.652~0.812之间; 按UPGMA法将96个品种划分为7个类群。综合SNP和系谱分析, 近10年河南省审定的96个小麦品种遗传多样性不够丰富, 多数品种亲缘关系较近, 在育种中迫切需要引入新的种质资源, 拓宽遗传背景。
, XIE Jing-Zhong
To understand the genetic diversity of wheat varieties in Henan Province at whole genome level, we selected 96 registered wheat varieties in the past decade for SNP genotyping using the Illumina 90k iSelect SNP chip. The results showed that 47.39% of the SNPs (38 661 out of 81 587) were polymorphism between varieties. The genome distribution of polymorphic SNPs showed a tendency of B > A > D. The genetic similarity of the 96 varieties ranged from 0.552 to 0.998 with an average value of 0.719. However, genetic similarity of 0.652 to 0.812 was found for 94.3% of the tested varieties, indicating highly similarity for most of the varieties in Henan province. The 96 wheat varieties could be classified as seven groups by UPGMA analyses. Based on pedigree and SNP analyses, very narrow genetic background was observed for the registered varieties in the past decade in Henan province. It is an urgent task for wheat breeders to explore and introduce diversified germplasm resources into wheat breeding program to increase the yield potential and stress tolerance in the future.
小麦(Tritium aestivum L.)是世界上主要粮食作物之一。在我国, 小麦是仅次于水稻和玉米的第三大粮食作物, 在农业生产中占有十分重要的地位。河南省是我国小麦主产区和商品粮产区, 也是小麦育种工作起步较早、发展较快的省份。自20世纪40年代以来, 河南小麦育种取得了很大成就, 促成生产用种9次大规模的更换。然而, 近十几年来, 由于在小麦育种中大量使用相同和相近的亲本, 导致新育成品种遗传基础日益狭窄、遗传变异率低[1, 2], 极大限制了小麦产量和品质的进一步改良, 而且还使小麦生产应对不良环境的能力大大降低, 增加潜在的用种风险。因此, 对河南省近年来审定小麦品种进行遗传多样性分析, 全面了解这些品种的遗传基础和亲缘关系, 对发掘利用优异的育种材料, 拓宽遗传基础, 加快小麦育种改良进程具有重要意义。
根据系谱分析小麦育成品种的遗传多样性及其与骨干亲本的亲缘关系, 虽然可以大致判断品种间遗传差异, 但难以量化, 存在一定局限性。一个新品种的选育过程往往是育种家对某一亲本经过连续多年的遗传改良, 最终选择出优良品系, 其遗传构成可能源于多个亲本, 加之部分品种系谱来源复杂或存在不确定性, 通过系谱追踪难以准确判断其遗传物质来源。近年来随着DNA分子标记技术的出现, 为在DNA水平上评估不同品种间的亲缘关系及计算品种间的遗传距离提供了新的方法和手段。Sun等[3]利用RAPD标记技术分析了西藏小麦、普通小麦和欧洲斯卑尔托小麦遗传多样性, 发现欧洲斯卑尔托小麦和西藏小麦遗传多样性远高于普通小麦; Wang等[4]利用SSR标记技术分析发现我国云南和西藏小麦品种的遗传多样性高于新疆小麦品种; Talebi等[5]利用AFLP标记分析表明伊朗小麦品种遗传多样性丰富。虽然RAPD、SSR和AFLP等分子标记均已应用于小麦品种的遗传多样性研究, 但受方法和位点数量的限制, 无法从全基因组水平上进行高通量遗传研究。
单核苷酸多态性(single nucleotide polymorphism, SNP)是普遍存在于生物基因组中的一种新型分子标记, 是指在基因组水平上由单个核苷酸的变异而引起的DNA序列单个碱基多态性变化。SNP是二等位基因, 具有在基因组中数量最多、分布密度高、无需电泳、可高通量自动化检测等特点[6, 7]。大规模、高通量SNP芯片检测首先广泛应用于人类群体结构的遗传学研究和关联性分析[8], 商业化的SNP芯片也已被应用于一些家畜全基因组关联分析、QTL定位和候选基因筛选[9, 10]。在植物中, Clark等[11]利用高通量SNP芯片技术对模式植物拟南芥不同品系进行了多态性研究。近年来, 随着小麦EST测序和高通量测序研究工作的开展, SNP开发和利用进展迅速[12]。Cavanagh等[13]和Wang等[14]开发了小麦基于Illumina技术平台的9k和90k高通量SNP分析芯片, 已开始应用于小麦遗传连锁图谱的构建、DNA指纹分析、群体结构和连锁不平衡分析, 以及基因定位等诸多领域。SNP标记能真实反映小麦品种间的亲缘关系[15, 16, 17], 可以用于小麦品种遗传多样性研究。
近十几年来, 河南省审定的小麦品种有近百个, 但尚未见到从全基因组水平上对其遗传多样性和亲缘关系进行研究的报道。本研究采用Illumina 90k iSelect SNP标记技术对近年来河南省审定的96个小麦品种的遗传基础进行分析, 旨在了解这些品种在全基因组水平上的遗传多样性和亲缘关系, 以期为小麦新品种选育的亲本选配提供理论依据。
除豫麦34外, 其他供试品种均为2000— 2013年河南省审定的品种或生产上种植面积较大的国家审定品种, 共计96个(见附表), 分别由品种育成单位提供, 各品种的系谱资料源自品种参试时育种单位提供的信息。
2012年10月在中国农业大学小麦遗传育种温室种植, 植株长至二叶一心时, 取小麦叶片, 按Saghai-Maroof等[18]的CTAB法提取基因组DNA。
利用Illumina 90k iSelect SNP标记对96个小麦品种进行全基因组扫描, 采用Genome Studio Polyploid Clustering Module v1.0进行样本的原始SNP分型, 由美国加州大学戴维斯分校基因组中心完成。
将SNP分型数据分别记为1 (纯合AA)、2 (杂合AB)、3 (纯合BB)和0 (缺失), 建立原始矩阵。用NTSYS-PC ver. 2.1 统计软件计算品种间遗传相似系数, 采用非加权配对算术平均法(UPGMA)构建遗传关系树状图。
在分布于21条染色体的81 587个SNP位点中, 38 661个被检测到多态性, 多态性比率为47.39%, 其中28 761个定位于染色体上。每条染色体分布106~2685个多态性位点, 差异较大, 且在A、B、D基因组上分布不均。以B基因组最多(14 049个, 占48.85%), 尤其是1B上最多, 2B和5B上次之; A基因组上多态性位点为11 548个, 占40.15%, 以5A、6A和7A上较多; D基因组上多态性位点最少, 仅有3164个, 占11.00%, 尤以4D上最少(图1-A)。整体来看, 1B、5B和2B染色体上多态性位点最多, 4D染色体上多态性位点最少。
多态性SNP位点在7个部分同源群中分布也不均匀, 在第1和第2部分同源群上分布最多, 其次是在第5和第6部分同源群上, 而在第4部分同源群最少(图1-B)。
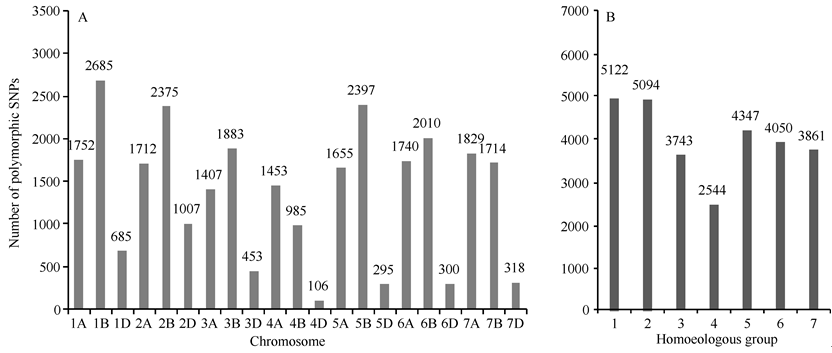 | 图1 多态性位点在染色体(A)和部分同源群(B)上的分布Fig. 1 Distributions of polymorphic SNPs on individual chromosome (A) and each homoeologous group (B) |
基于SNP标记数据的品种两两间遗传相似系数, 其变异范围为0.552~0.998, 平均值为0.719, 其中中育9号与矮抗58间的遗传相似性最低(0.552), 而周麦22与周麦23间的遗传相似性最高(0.998)。
96个供试品种两两计算, 共获得4560个遗传相似系数, 以0.02为组距进行次数分布分析, 可见, 96个品种间的遗传相似系数在0.7处分布密度最大, 两侧不对称(图2)。经卡平方测验, 不符合正态分布(P < 0.01), 呈左偏态分布; 大于0.7的遗传相似系数有2812个(61.67%), 小于0.7的有1748个(38.33%); 主要分布在0.63~0.83之间, 占95.94% (4375/4560)。
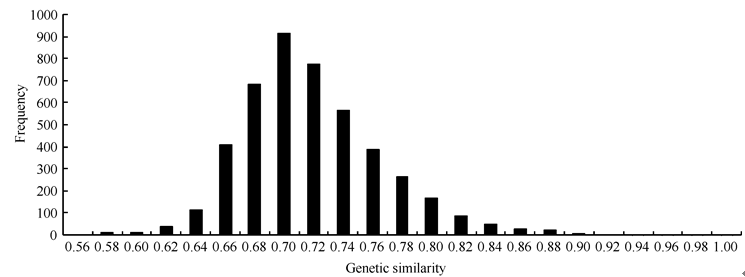 | 图2 品种间遗传相似系数的次数分布Fig. 2 Distribution of genetic similarity between varieties |
为了探讨近十几年来河南省审定小麦品种遗传多样性的演变趋势, 将96个品种按审定年份分为3组, 每组的品种数接近, 可见组内品种的遗传相似性均较高, 且自2005年以后平均遗传相似系数有升高趋势(表1)。
| 表1 不同阶段审定品种的遗传相似系数 Table 1 Genetic similarity coefficient of Henan wheat cultivars at different period |
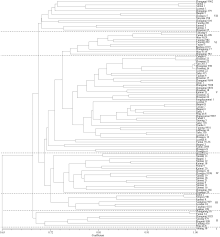
根据小麦SNP遗传相似系数矩阵进行聚类分析, 96个品种被划分为7大类群, 其中矮抗58单独组成第I类, 第II和第III类各有7个品种, 第IV类有17个品种, 第V类41个品种, 第VI和第VII类分别包括10个和13个品种(图3)。
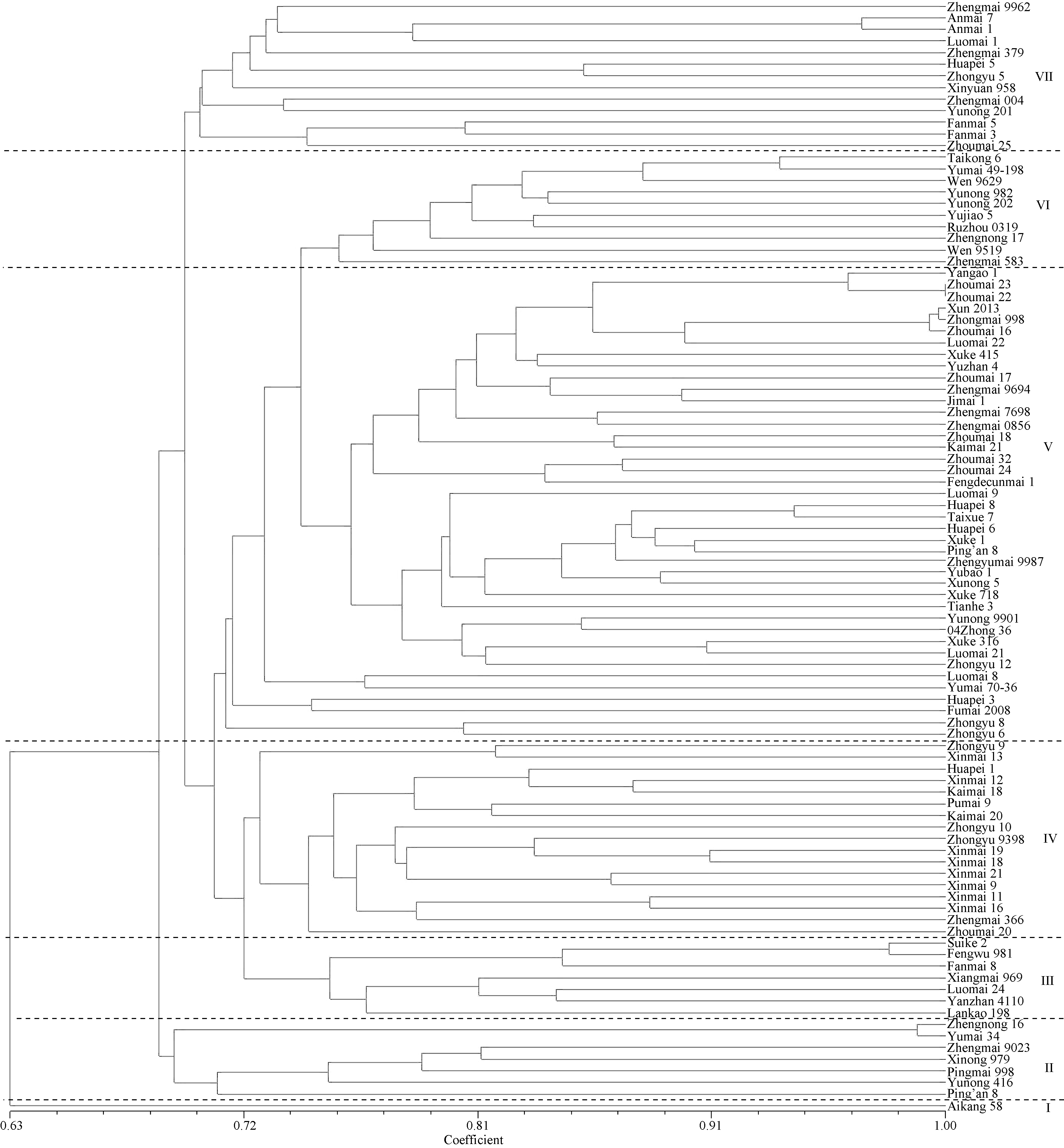 | 图3 河南省96个审定小麦品种的SNP数据聚类图Fig. 3 Phylogenetic tree of 96 registered wheat varieties in Henan Province revealed by SNP |
第II类品种间的遗传相似系数变幅在0.658~ 0.988之间, 平均0.735, 有4个为优质强筋品种, 其中豫麦34是郑农16 (豫麦34/小偃6号)的亲本之一, 二者相似系数为0.988; 郑麦9023、西农979和平麦998均是以小偃6号作为间接亲本而育成的品种, 在遗传基础上具有很高的亲缘性。
第III类品种间遗传相似系数为0.702~0.976, 平均0.784。这些品种均具有早熟特性, 除泛麦8号和洛麦24外, 其他均为直接或间接利用豫麦18育成的品种。丰舞981是漯麦4号的系选品种, 而漯麦4号的母本是豫麦18; 睢科2号的母本是漯麦4号, 所以丰舞981与睢科2号的亲缘关系较近, 其遗传相似系数为0.976; 洛麦24并非由豫麦18直接选育而成, 但其亲本之一的洛太911在田间表现与豫麦18极为相似, 且均为偃师4号的后代。
第IV类的17个品种间遗传相似系数变幅在0.681~0.906, 平均0.761。这些品种是以新乡市农业科学院选育的品种及其衍生品种为主, 它们大多是以百泉3047-3、百泉3199、内乡82C6为亲本选育出的品种。如新麦18和新麦19都是以(C5/新乡3577)F3 d1s作为母本, 以新麦9号作为父本选育而成的小麦品种, 其遗传相似系数为0.906; 周麦20的亲本之一也是新麦9号。
第V类品种数最多, 有41个, 占42.7%, 品种间遗传相似系数为0.647~0.998, 平均0.769。这些品种的亲本大多含有豫麦21、周麦11及8425B中的1个或2个, 有的则源自这3个品种的衍生系, 如以周麦16、周麦13等为亲本选育出的品种, 均聚在这一类群中。
第VI类的10个品种间遗传相似系数变幅最小(0.719~0.933)之间, 平均值最高(0.801)。它们主要是以豫麦2号或者其衍生系豫麦49及豫麦41为亲本之一育成的品种。如太空6号是豫麦49通过航天诱变选育而成; 豫麦49-198为豫麦49的系选品种, 两者的相似系数为0.933。
第VII类包含13个品种, 其遗传相似系数变幅在0.664~0.966之间, 平均值为0.721, 均是以偃师4号衍生系, 如豫麦18、周麦8826、豫麦10号、豫麦21为亲本之一育成的品种, 并且都继承了偃师4号的优良性状。
聚类分析结果表明, 河南省近十几年来审定的品种在DNA水平上的差异不大, 大部分品种的亲缘关系较近, 遗传基础较窄。
近年来, 有多项研究利用分子标记、系谱和农艺性状分析评价小麦品种的遗传多样性[1, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29]。但由于受环境等自然因素的影响和标记数量的限制, 无法从全基因组水平上进行研究。本研究选用分布于小麦21条染色体上的81 587个SNP位点对河南省近年来审定的96个小麦品种进行了全基因组扫描, 发现SNP位点多态性比率为47.39%, 其中, 以B基因组上的多态性位点最多, A基因组次之, D基因组最少, 与贾继增等[19]运用RFLP标记对来自11个国家的15个小麦品种(系)进行遗传多样性分析研究结果一致。究其原因, 主要是普通小麦进化过程中D基因组与AABB基因组形成六倍体小麦时间较短, 且来源比较单一, 品系间多态性较低。另外D基因组可能携带较多的与普通小麦广适应性、抗逆性和加工品质等重要性状的关键基因, 在长期的选择和育种过程中, 所承受的选择压力大于A和B基因组, 从而造成更多、更强的选择牵连效应发生, 导致其遗传多样性低。本研究中尤其以4D染色体上的多态性位点最少, 这与郝晨阳等[1]和倪中福等[20]研究结果一致。推测与4D染色体上可能携带较多控制重要性状的基因(如育性基因、矮秆基因等), 在育种过程中施加了较强的定向选择所产生的选择牵连作用, 导致其遗传多样性显著偏低。贾继增等[19]报道过与此不同的结果, 他们研究发现, 1D和5D遗传多样性最小, 通过分析多态性RFLP位点在7个部分同源群的分布发现, 第1和第2部分同源群最多, 第4部分同源群最少。这可能是由于不同试验所选取的材料来源不同造成的。
许多研究表明, 我国不同地区都存在小麦主栽品种亲缘关系较近和遗传基础不够广泛的现象[21, 22, 23, 24, 25, 26, 27, 28, 29]。本研究进一步证实河南省近期选育的小麦品种存在亲缘关系近、遗传相似性高的问题, 并且有进一步加重的趋势, 这将导致很难获得突破性育种进展, 降低品种抵御各种自然灾害的能力, 增加潜在的用种风险。造成河南省小麦品种遗传多样性水平低、遗传基础狭窄的原因可能一是核心骨干亲本的反复利用, 二是定向人工选择。本研究通过聚类和系谱分析, 发现河南省小麦骨干亲本品种主要由6大血缘系统组成, 分别是周麦、新麦、偃师4号、豫麦2号、豫麦18和陕西小麦系统。周麦系统主要是以周麦品种和周8425B为亲本材料, 利用该血缘系统育成的品种数量最多, 占整个参试品种总数的42.7%, 高产、稳产性特点突出; 新麦系统主要以百泉3047-3、百泉3199、内乡82C6为亲本; 偃师4号系统包括以偃师4号为间接亲本选育的品种; 豫麦2号系统是以豫麦2号及其后代豫麦49作为育种亲本; 豫麦18系统均包括豫麦18血统, 突出特点是早熟; 陕西小麦血统为亲本选育的品种的主要特点是优质强筋。定向人工选择可能是河南省小麦遗传多样性降低的另一个主要原因。河南省地处黄淮麦区, 生态条件适宜小麦的生长发育和产量潜力的发挥, 育种家根据对产量有利的目标性状进行选择的趋势, 导致部分多样化基因位点丢失, 使品种的优良性状趋于类似。由于周麦系列具有较好的综合农艺性状和丰产性, 得到大部分育种家的认可和偏爱, 成为选配高产组合的首选亲本材料, 因此导致近十几年来河南省审定小麦品种中有将近一半直接或间接含有周麦品种血缘。
虽然聚类结果与品种特性和系谱信息有较好的一致性, 但也有例外。如矮抗58是以周麦11为母本, 以豫麦49和郑州8960杂交后代为父本选育而成的品种, 但它并没有与周麦11和豫麦49的后代品种聚在一类; 浚2016和众麦998之间的相似系数为0.996, 田间长相也十分接近, 但系谱记载表明两者没有直接的亲缘关系。其可能原因一是供试种子来源, 由于亲本材料的搜集渠道很多, 可能会存在材料本身与名称不符的现象; 二是育种家定向选择的结果, 多年的选择往往都集中在某些或某个位点, 从而导致其他某些或某个位点的丢失, 尤其在同一生态区; 三是系谱记载不完善。另外, 遗传变异(含重组)也可能是产生上述现象的重要原因。
值得注意的是, 尽管近年来河南省小麦育种中因大量使用相同或类似的骨干亲本或骨干亲本衍生材料做亲本, 致使育成的小麦品种遗传差异不明显、遗传基础狭窄、多样性水平较低, 但可以看出, 河南省的小麦品种间仍然存在一定的类群分化和遗传多样性, 可分为周麦类型、新麦类型、陕西优质类型、豫麦2号类型和早熟豫麦18类型。纵观河南省生产上得以大面积推广利用的代表性小麦品种, 它们都具有独特的遗传构成, 其中以矮抗58最为典型, 该品种表现高产、稳产、广适、抗性强, 在聚类图中自成一类, 众多优异性状的组合是否来自有利基因的聚合和重组, 有待深入研究。
针对河南省小麦育种策略, 建议一方面要继续广泛收集种质资源, 加强与国内外育种单位资源材料间的交流与合作, 不断拓宽小麦亲本的遗传基础; 另一方面要大力加强种质资源的创新研究和利用, 利用当地的高产小麦品种与从国内外引进的优异基因资源进行聚合杂交和回交转育, 创造新的优异中间育种材料, 突破主产区小麦育种的瓶颈, 使河南省小麦育种水平跨上新台阶。
多态性SNP位点在不同染色体和7个部分同源群间分布不均匀, 其中以4D染色体上的多态性位点最少。品种间的遗传相似度较高, 遗传相似系数平均值为0.719, 并且有增加的趋势。河南省小麦品种间存在一定的类群分化和遗传多样性, 代表性小麦品种都具有其独特的遗传构成。
| 附表 用于SNP分析的小麦品种名称及系谱 Supplementary table The wheat varieties for SNP genotyping and their pedigrees |
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|