

{kind=link}
{kind=link}
{kind=link}
{kind=link}
小麦-华山新麦草异代换系DH2322的分子细胞遗传学鉴定
[王秀娟 , 陈新宏, 庞玉辉, 敬樊, 张军, 胡思远, 昝凯, 武军, 杨群慧, 赵继新
, 陈新宏, 庞玉辉, 敬樊, 张军, 胡思远, 昝凯, 武军, 杨群慧, 赵继新* ]
, 陈新宏, 庞玉辉, 敬樊, 张军, 胡思远, 昝凯, 武军, 杨群慧, 赵继新]
|
|


第一作者联系方式: E-mail: wangxiujuanluck@163.com
利用分子标记、细胞学、基因组原位杂交(GISH)等技术, 结合田间农艺性状调查, 对小麦-华山新麦草七倍体材料H8911与硬粒小麦D4286杂交F4代分离群体中株系DH2322进行了综合鉴定。华山新麦草基因组特异SCAR标记鉴定表明, DH2322含有华山新麦草遗传物质; 细胞遗传学观察显示, DH2322染色体构型为2 n=42=21 II; 有丝分裂和花粉母细胞减数分裂中期I基因组原位杂交(GISH)鉴定表明, DH2322的染色体由40条小麦染色体和2条华山新麦草Ns染色体构成, 且2条Ns染色体能完全配对为一个二价体; SSR和STS分子标记分析表明, DH2322缺失小麦D基因组的2D染色体, 而含有华山新麦草的2Ns染色体; 农艺性状分析结果表明, DH2322具有双亲的形态学特征, 结实性好, 穗长和穗粒数显著大于亲本。
, CHEN Xin-Hong, PANG Yu-Hui, JING Fan, ZHANG Jun, HU Si-Yuan, ZAN Kai, WU Jun, YANG Qun-Hui, ZHAO Ji-Xin
A wheat- P. huashanica alien substitution line DH2322 was isolated from the F4 progeny of heptaploid hybrid H8911 × Triticum durum cv. D4286 and identified by using SCAR markers, genomic in situ hybridization (GISH) , SSR markers, and STS markers. The results using SCAR marker RHS141 showed that DH2322 carrys chromosome derived from P. huashanica. Cytogenetics observation indicated that DH2322 has a chromosome karyotype of 2 n = 42 = 21 II. GISH showed that DH2322 is a line with 40 chromosomes from wheat and two Ns chromosomes from P. huashanica, which formed a ring bivalent at metaphase I in pollen mother cells (PMC). According to SSR and STS analysis, the wheat chromosome 2D in DH2322 was substituted by the chromosome 2Ns from P. huashanica. Evaluation of the agronomic traits showed that DH2322 presented favorable agronomic traits in spike length and kernels per spike. Development of this new germplasm will be useful for wheat breeding programs in the future.
华山新麦草(Psathyrostachys huashanica Keng, 2n=14, NsNs)是禾本科(Gramineae)、小麦族(Triticeae)、大麦亚族(Hordeinae)、新麦草属(Psathyrostachys)多年生异花授粉植物, 具有抗寒、抗旱、耐瘠薄、早熟、优质、矮秆、抗病等优良特性[1], 仅分布于我国秦岭山脉陕西华山段, 是我国特有种[2], 也是小麦(Triticum aestivum, 2n=14, AABBDD)品种改良的优异种质资源。越来越多的研究者开始将研究目标瞄准华山新麦草的利用并取得了一些进展。Kang等[3, 4]利用中国春ph2b基因突变体和华山新麦草杂交获得了含有ph2b基因的衍生后代、并利用面包小麦“ J-11” 和华山新麦草杂交获得一个八倍体属间杂种PHW-SA (2n=8x=56, AABBDDNsNs)。Wang等[5]研究发现华山新麦草3Ns染色体上携带抗小麦条锈病的基因, Kang等[6]将华山新麦草3Ns染色体导入小麦, 获得了抗条锈病的小麦-华山新麦草中间材料。另外, Liu等[7]和Wang等[8]利用简单重复序列标记(simple sequence repeats, SSR)技术对华山新麦草种群基因的多样性进行了分析。
本课题组自20世纪90年代初开始进行普通小麦品种7182与华山新麦草的杂交研究, 选育出普通小麦-华山新麦草七倍体材料H8911 (2n=49, AABBDDNs)[1]。在此基础上, 侯文胜等[9]、傅杰等[10]、赵继新等[11, 12, 13]、武军等[14]利用细胞遗传学对小麦-华山新麦草异附加系、异代换系进行了分子细胞遗传学方面的研究; Du等[15, 16, 17, 18, 19, 20]选育出一批附加华山新麦草2Ns-7Ns染色体的小麦-华山新麦草二体异附加系; 王秀娟等[21]获得一个附加两条华山新麦草7Ns染色体的矮秆材料12DH25。
本研究以小麦-华山新麦草七倍体材料H8911与硬粒小麦品种D4286杂交的F4分离群体中筛选的株系DH2322为材料, 从细胞遗传学、分子标记和农艺性状等方面对其进行研究, 以了解其遗传行为、染色体构成、所含华山新麦草染色体的同源群归属及形态特征等, 为小麦品种改良和染色体工程育种提供新的种质资源。
普通小麦品种7182、华山新麦草(Psathyrostachys huashanica)、倍半二倍体H8911 (2n=49, AABBDDNs)及其与硬粒小麦(Triticum durum, 2n=28, AABB)品种D4286杂交并回交后选育的F4代材料DH2322均来自陕西省植物遗传工程育种重点实验室。
2012年10月和2013年10月于西北农林科技大学试验田种植供试材料, 调查田间农艺性状和室内考种, 统计株高、旗叶面积、穗下茎节长度、穗长、小穗数、穗粒数和结实率。用SAS软件分析数据和检验差异显著性。
采用CTAB法[22]提取普通小麦品种7182、华山新麦草、硬粒小麦品种D4286、普通小麦-华山新麦草七倍体材料H8911及DH2322的基因组DNA。
为提高小麦背景中华山新麦草染色质的鉴定效率, 根据Du等[23]开发的华山新麦草染色体特异SCAR标记RHS141, 以7182、D4286、DH2322、H8911和华山新麦草基因组DNA为模板进行扩增。在PTC-200扩增仪上进行PCR, 20 μ L总反应体系包括10× buffer (含Mg2+) 2.0 μ L、引物RHS141 (2.5 μ mol mL-1) 2.0 μ L、模板DNA (200 μ g μ L-1) 2.0 μ L、dNTP (2.5 μ mol mL-1) 1.6 μ L、Taq DNA聚合酶(5 U μ L-1) 0.2 μ L、ddH2O 12.2 μ L。PCR程序为, 94℃预变性3 min; 94℃变性50 s, 60℃复性40 s, 72℃延伸 1 min, 35个循环; 72℃最后延伸10 min, 4℃保存。以1%琼脂糖凝胶135 V恒压条件下电泳30 min, 溴化乙锭(EB)染色, 显影后照相观察。
将种子于室内浸泡至露白, 转移到垫有湿润滤纸的培养皿内发芽。当根长至1~2 cm时, 用镊子夹取根尖放入0~4℃冰水中预处理24 h, 卡诺氏I固定液(无水乙醇∶ 冰醋酸= 3∶ 1)固定, -20℃冰箱保存。45%醋酸洋红染色压片, 用Olympus BX60显微镜观察根尖细胞染色体数, 并照相。
从田间取适龄幼穗(旗叶与倒二叶距离约为4 cm), 用卡诺氏II固定液(无水乙醇∶ 氯仿∶ 冰乙酸= 6∶ 3∶ 1)固定, 45%醋酸洋红染色压片, 用Olympus BX60显微镜观察花粉母细胞减数分裂中期I (PMC MI)的染色体构型, 并照相。
将DH2322的根尖和幼穗固定一周后用体积分数45%醋酸压片, 液氮揭片, 备用。
依照Dig-Nick-Translation Mix试剂盒(Roche, Germany)产品说明书对华山新麦草的全基因组DNA进行标记(4 μ L Dig-Nick-Translation Mix, 1 μ L华山新麦草DNA (1000 μ g μ L-1), 加ddH2O至20 μ L)。参照Han等[24]描述的方法进行原位杂交。每张制片加40 μ L杂交液, 该液含探针DNA 100 ng、100%去离子甲酰胺20 μ L、50% (w/v)硫酸葡聚糖8 μ L、20× SSC 4 μ L、ssDNA (鲑鱼精DNA, 5 μ g μ L-1) 1 μ L、10% (w/v)十二烷基磺酸钠1 μ L、加ddH2O至40 μ L。在Hybrite原位杂交仪上进行杂交反应, 75℃ 6 min, 然后37℃ 16 h以上。用荧光素(FITC)检测杂交信号, 洗脱后用PI复染及H-1300 (抗退色剂)封片, OLYMPUS BX60荧光显微镜观察检测, Pixera Penguin 150CL CCD成像系统照相。
根据Rö der等[25]和Pestsova等[26]公布的小麦21条染色体特异SSR标记, 分别选取各条染色体长、短臂上各10对引物进行扩增。从网站http://wheat. pw.usda.gov/SNP/new/pcr_primers.shtm上选取小麦7个部分同源群上的STS标记, 分别选取各同源群6对标记进行扩增。所选引物由生工生物工程(上海)有限公司合成。PCR扩增反应体系和步骤参照本文SCAR标记鉴定, 退火温度依引物而定。扩增产物点样量为6 μ L, 经8%非变性聚丙烯酰胺凝胶电泳, 180 V恒压电泳2~3 h。硝酸银染色, 用数码相机照相。
表型调查结果显示, DH2322农艺性状稳定, 穗长较小麦亲本7182、D4286显著增长, 穗粒数也较D4286和七倍体材料H8911显著增大, 但与小麦亲本7182差异不大(表1)。整体上看, DH2322具有双亲的形态学特征, 结实性好, 籽粒饱满, 是较好的遗传材料。
| 表1 异代换系DH2322及其亲本农艺性状 Table 1 Agronomic traits of the substitution line DH2322 and its parents |
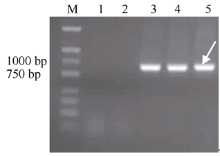
用华山新麦草Ns基因组特异SCAR标记RHS141对普通小麦品种7182、硬粒小麦品种D4286、DH2322、H8911及华山新麦草的基因组DNA进行PCR扩增表明, 华山新麦草、H8911和DH2322的DNA可以扩增出934 bp的华山新麦草特异条带, 而普通小麦品种7182、硬粒小麦品种D4286的DNA不能扩增出条带, 说明DH2322含有华山新麦草Ns基因组染色体(图1)。
 | 图1 华山新麦草基因组特异SCAR标记分析Fig. 1 Amplification profile of the SCAR marker specific to P. huashanicaM: marker D2000; 1: 7182; 2: D4286; 3: DH2322; 4: H8911; 5: P. huashanica. |
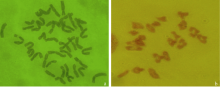
DH2322根尖细胞染色体制片观察, 发现根尖细胞染色体数目为2n=42 (图2-a); 对52个花粉母细胞镜检显示, 47/52 (90.4%)花粉母细胞减数分裂中期I染色体形成21个二价体(图2-b), 其中环状二价体占有很高比例, 平均每个细胞有20.91个(表2); 以华山新麦草总基因组DNA作为探针, 对DH2322根尖细胞和花粉母细胞基因组原位杂交鉴定显示, 其根尖细胞中有两条染色体为黄绿色(图3-a), 花粉母细胞减数分裂中期I细胞有1个完整的二价体显示荧光信号, 表明DH2322中的2条Ns基因组染色体可以完全配对为1对染色体(图3-b)。由此说明, DH2322是一个遗传稳定, 包含40条小麦染色体和1对华山新麦草Ns基因组染色体的小麦-华山新麦草异代换系。
 | 图2 DH2322根尖细胞(a)和花粉母细胞(b)观察Fig. 2 Cytogenetic observation of root-tip cell (a) and pollen mother cell (b) of DH2322 |
 | 图3 DH2322根尖细胞原位杂交(a)花粉母细胞原位杂交(b)Fig. 3 Genomic in situ hybridization (GISH) pictures for root-tip cell (a) and pollen mother cell (b) of DH2322 |
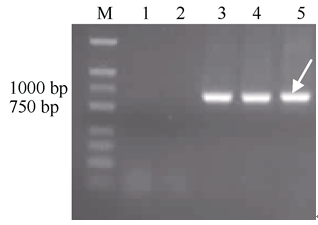
用小麦21条染色体上共210对SSR引物对DH2322及其亲本进行分子标记分析表明, 2D染色体上的3对引物(Xgwm249、Xgwm296、Xgwm484)在小麦品种7182和H8911中扩增出154、182、153 bp的目标条带, 但在硬粒小麦D4286、华山新麦草和DH2322中未扩增出目标条带(图4)。推测DH2322缺失了小麦2D染色体。
| 表2 DH2322花粉母细胞减数分裂中期I (PMC MI)染色体配对 Table 2 Chromosome pairing at metaphase I in pollen mother cells of substitution line DH2322 |
选取定位于小麦7个部分同源群上的42对EST- STS标记, 对普通小麦品种7182、硬粒小麦品种D4286、DH2322、H8911及华山新麦草的基因组DNA进行PCR检测, 结果只有定位于小麦第2同源群的引物CD453246和BQ160526在DH2322中扩增出华山新麦草特异条带(图4)。推测DH2322可能含有华山新麦草2Ns染色体。
 | 图4 SSR及STS引物扩增图谱Fig. 4 Amplification profile of SSR and STS markersM: marker D2000; 1: 7182; 2: D4286; 3: DH2322; 4: H8911; 5: P. huashanica. |
实现华山新麦草有益基因向小麦的转移, 研究华山新麦草染色体的遗传效应以及获得小麦-华山新麦草中间材料, 如双二倍体、异附加系、异代换系是十分必要的。目前, 育种工作者已获得多个小麦-华山新麦草杂交后代, 它们表现型上多种多样, 如小麦-华山新麦草异附加系有矮株、分蘖数增多、抗条锈病、叶锈病等性状[15, 16, 17, 18, 19, 20, 21]; Cao等[27]鉴定出一个抗条锈病易位系材料。小麦-华山新麦草异代换系DH2322作为小麦与华山新麦草远缘杂交的新类型, 其穗长与穗粒数较亲本小麦品种显著增加, 是创制小麦-华山新麦草易位系的重要桥梁。华山新麦草染色体导入的同时, 必然与小麦遗传背景发生相互作用, 而导致穗长与穗粒数显著增大的原因, 尚需进一步研究。
外源基因的有效转移很大程度上依赖于对导入小麦背景中的外源染色体或染色体片段的快速准确鉴定[28]。GISH可以直观地检测外源染色体或染色体片段在小麦遗传背景中的分布, 在鉴定远缘杂种染色体构型和结构变异方面有重要作用, 但其技术难度大、费时且成本较高, 在初步鉴定大量遗传材料时有一定的局限性。SCAR标记技术的发展为小大麦远缘杂交研究提供了极大方便。RHS141是Du等[23]用RAPD技术筛选的华山新麦草基因组特异DNA片段, 在小麦族其他物种和华山新麦草及全套小麦-华山新麦草二体异附加系中验证发现, 仅含华山新麦草染色质的物种能扩增出长为938 bp的片段。RHS141可用于含华山新麦草特异染色质材料的检测。本研究中RHS141的应用为快速初步判定12DH22中含有华山新麦草遗传物质提供了方便。
SSR分子标记在近几年的小麦遗传作图、基因定位、系谱或亲缘关系分析、标记辅助选择等研究中得到广泛应用[29]。Rö der等[25]建立了小麦的第一张微卫星遗传图谱, 将230对引物扩增的279个位点整合到国际小麦簇作图计划(ITIM)的Opata85 × W7984作图群体遗传框架图上; Pestsova等[26]对从粗山羊草分离的D基因组专化性微卫星标记(GDM)在小麦上进行了作图。国际小麦微卫星协会(Wheat Microsatellite Consortium, WMC)也将开发的66个WMC标记整合到已有的小麦遗传框架图上[30]。本研究利用七倍体H8911与硬粒小麦D4286杂交选育DH2322的过程中, 由于来自H8911的A和B基因组与来自硬粒小麦的A和B基因组, 理论上可以很好地配对而得以完整遗传, 因此推测, 二体异代换系DH2322最大可能是由华山新麦草的1对Ns基因组染色体替换了1对小麦D基因组染色体。为了确定DH2322所缺失的小麦染色体, 本研究采用Pestsova等[26]开发的来自粗山羊草专化性微卫星标记对小麦-华山新麦草异代换系DH2322进行分析, 结果显示, D基因组的2D染色体上的3对引物, 在DH2322中没有扩增出预期的目标条带, 而其他基因组染色体上的引物, 在DH2322中扩增出预期的目标条带。由此推断, DH2322缺失了小麦D基因组的2D染色体。
STS (Sequence-tagged site)即序列标签位点, 是基因组上定位明确, 作为界标并能通过PCR扩增被唯一操作的短的单拷贝DNA序列[31], 由于可直接在DNA分子水平上指出小麦中外源染色体片段的组成和位置, 在外源染色体鉴定方面应用广泛。前人利用STS标记已成功鉴定出小麦背景中的簇毛麦、黑麦等小麦近缘种的染色体片段[32, 33, 34]。Du等[15]利用12对STS标记鉴定出3-6-4-1附加两条华山新麦草2Ns染色体, 其中的2对标记在华山新麦草和DH2322中扩增出与华山新麦草一致的特异条带, 说明DH2322含有华山新麦草Ns基因组的2Ns染色体。CD453246扩增出的H8911的条带本应与华山新麦草特异条带高度一致, 但H8911的条带却普遍高于后代DH2322的扩增条带, 原因尚不清楚。3-6-4-1和DH2322穗长较亲本7182都显著增长3~4 cm, 是否与2Ns染色体上携带的基因有关尚需进一步研究。
DH2322是一个稳定的小麦-华山新麦草2D(2Ns)二体异代换系材料, 结实性好, 穗长和穗粒数等农艺性状上得到改善, 对进一步研究华山新麦草染色体的形态结构特征及遗传机制、挖掘利用华山新麦草优异基因、丰富小麦遗传种质资源具有重要价值。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|