{kind=link}
枯萎病菌诱导感、抗陆地棉品种的转录因子表达变化
[韩泽刚1  , 赵曾强
, 赵曾强1 , 何兰兰1 , 柴蒙亮2 , 李会会1 , 张薇1, * ]
, 赵曾强]
|
|


第一作者联系方式: E-mail: hanzegang501706851@126.com, Tel: 13779593063
以陆地棉抗病品种中棉所12和感病品种新陆早7号为材料, 利用Solexa高通量测序技术调查枯萎病菌诱导后不同时间感、抗陆地棉品种转录因子家族及转录因子的表达情况。结果表明, 枯萎病菌诱导后, 抗病品种中棉所12有39个转录因子家族的433个转录因子在至少一个比对组中表达活性发生变化; 感病品种新陆早7号则有52个转录因子家族的588个转录因子在至少一个比对组中表达活性发生变化。新陆早7号对枯萎病菌响应的转录因子及转录因子家族的数目明显多于中棉所12, 且2个品种的下调基因数目均多于上调基因。随着枯萎病菌诱导后时间延长, 两品种对枯萎病菌诱导响应的转录因子家族及转录因子数量均呈现先增加后降低的变化趋势, 中棉所12在枯萎病菌诱导后6 h达最多, 而新陆早7号在诱导后3 h达最多。在6个比对组中, 表达活性均发生变化的重叠转录因子, 中棉所12中有9个, 隶属于6个转录因子家族; 新陆早7号中有31个, 隶属于17个转录因子家族。不同抗病性品种对枯萎病的响应有较强的品种特异性, 除37个共有的转录因子家族外, 2个转录因子家族是中棉所12所特有, 15个转录因子家族是新陆早7号所特有。
The Solexa sequencing technique was used to study the expression changes of transcription factors and their families in upland cotton cultivars susceptible and resistant to Fusarium wilt. A total of 39 transcription factor families and 433 transcription factors in Zhongmiansuo 12 and 52 transcription factor families and 588 transcription factors in Xinluzao 7 were changed in expression in at least one comparison group induced by Fusarium wilt. The number of transcription factor families responsed to Fusarium wilt was more in Xinluzao 7 than in Zhongmiansuo 12, and the number of down-regulated genes was more than that of up-regulated genes in two varieties. After the induction by Fusarium wilt, in the process of time, the number of transcription factor families and transcription factors responsed to the Fusarium wilt in two cultivars showed the same changing trend of increase first and decrease then. The maximum number was reached after six hours in Zhongmiansuo 12, while at three hours after the inducing in Xinluzao 7. Among six comparison groups, there were nine overlapping transcription factors in six transcription factor families of Zhongmiansuo 12 and 31 overlapping transcription factors in 17 transcription factor families of Xinluzao 7. The expression of transcription factor showed strong cultivar specificity among varieties tolerant to Fusarium wilt. Except for 37 transcription factor families shared between two cultivars, two transcription factor families were unique to Zhongmiansuo 12 and Zhongmiansuo 15 transcription factor families were unique to Xinluzao 7.
棉花枯萎病是一种危害严重的世界性病害。枯萎病侵染棉花后, 前期往往大量死苗, 后期叶片及蕾铃大量脱落, 甚至造成植株死亡, 严重影响棉花的纤维品质及产量, 对棉花生产造成极大威胁[1, 2]。
转录因子即反式作用因子, 是一种能够与真核生物启动子特定DNA序列相结合的蛋白质, 其功能是激活或抑制基因的转录。已有研究认为, 当植物受到病原物侵袭时, 首先感受到相应信号, 然后通过一系列途径传递到作用位点, 进而作出防御应答。植物虽然对不同病原信号的反应不同, 但信号分子均可将信号传递到转录因子并将其激活, 通过转录因子与启动子相互结合启动/抑制靶基因的转录表达, 或者形成同源或异源二聚体, 或者直接与蛋白质互作引起一系列生理、生化反应, 使植株表现出抗病性[3]。
转录因子及其家族在植物逆境胁迫中起重要的调控作用。赵明辉等[4]利用基因芯片技术研究发现, 低氮胁迫下水稻品种沈农196和风锦剑叶中分别有53个和27个转录因子相关基因表达发生变化, 同时低氮胁迫响应转录因子相关基因表现出品种(系)特异性。马廷臣等[5, 6]利用基因芯片技术研究发现PEG胁迫下耐旱品种和干旱敏感品种中分别有95个和129个转录因子转录本与对照相比表达发生变化, 同样发现PEG 胁迫响应转录因子转录本表现出品种特异性。由于基因芯片技术只能分析已知序列, 加之背景噪音及假阳性率高, 具有无法检测出低丰度基因和新转录本等缺点, Solexa测序技术则克服了以往DNA微矩阵芯片等技术的不足, 在研究差异表达基因方面显示出强大优势[7, 8]。
近几年, 已有一些学者利用Solexa高通量测序技术, 对芝麻、大豆、花生、棉花等植物在转录水平上基因的表达变化进行了分析[9, 10, 11, 12], 该技术在研究植物的逆境胁迫方面也得到了广泛应用[13, 14, 15, 16, 17, 18, 19]。本研究利用Solexa高通量测序技术调查枯萎病菌诱导后陆地棉感、抗品种在不同时间转录因子家族及转录因子的表达变化, 为进一步阐明转录因子在棉花与枯萎菌互作中的调控机制及筛选抗枯萎病相关基因奠定基础。
陆地棉抗病品种中棉所12和感病品种新陆早7号由石河子大学棉花研究所提供。挑选经浓硫酸脱绒籽粒饱满的种子, 经0.1%氯化汞浸泡15 min消毒, 无菌水冲洗4~5遍, 种植于无菌蛭石, 待棉苗长至3 cm左右, 将其移入盛有Hoagland营养液的塑料盒(用带有孔洞的泡沫板漂浮棉苗), 于25℃、光16 h/暗8 h的培养箱中继续培育, 每周换一次培养液[20]。
将生长于PDA培养基的枯萎病菌菌落(菌种为枯萎病7号生理小种强致病力菌系F430, 由石河子大学植物保护系张莉老师提供)接种到装有已灭菌的查氏培养液, 28℃振荡培养4 d, 配制成浓度为7× 106 mL-1的孢子悬浮液。将长至二叶一心的棉花幼苗浸泡到孢子悬浮液45 min后转入Hoagland营养液, 并分别采集中棉所12和新陆早7号被侵染后0、1、3、6、12、24和48 h的根部组织, 经液氮速冻后保存于-80℃冰箱。然后将棉花根部组织样品送往深圳华大基因测序。
以测序得到的clean reads作为后续试验的基础, 测序结果经分析合格后, 运用RPKM法(Reads per kb per million reads)计算基因表达量[21]。在此基础上, 采用Audic和Claverie[22]描述的方法, 对表达谱中每个基因进行显著性分析。错配率(false discovery rate, FDR)则被用于确定P的阈值[23]。符合P< 0.005, FDR≤ 0.001以及│log2 ratio│≥ 1的基因被定义为显著差异表达基因, 这些基因被认为表达活性发生了变化, 反之可认为表达活性未发生变化。
最后对所有差异表达基因进行GO功能注释及分类, 在分子功能注释本体中找到转录因子活性(nucleic acid binding transcription factor activity)类别, 并将各个序列在NCBI中进行Conserved Domain Search (http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi?) 查找保守域, 找出所属的转录因子家族。
中棉所12在枯萎病菌侵染后1 h与0 h (对照)进行比对记为Z1; 侵染后3 h与0 h比对记为Z3; 侵染后6 h与0 h比对记为Z6, 侵染后12 h与0 h比对记为Z12, 侵染后24 h与0 h比对记为Z24, 侵染后48 h与0 h比对记为Z48。新陆早7号则依次记为X1、X3、X6、X12、X24和X48。
表1表明, 共有39个转录因子家族的433个转录因子表达活性在至少一个比对组中发生了变化。其中比对组Z1中有21个转录因子家族的65个转录因子基因(上调基因31个, 下调基因34个)表达活性发生变化。在比对组Z3中有32个转录因子家族的94个转录因子基因(上调基因21个, 下调基因73个) 表达发生变化; 与Z1相比, Z3缺少Jmjc和zf-C2H2转录因子家族, 却增加了ANK、ARS2、CBFB_ NFYA、E2F_TDP、EIN3、GAGA_bind、MBF1、R3H、Ras-like-GTPase、Sigma70 like、TCP、TOP4c、ZnF_GATA等13个转录因子家族。Z6中有34个转录因子家族的97个基因(上调基因16个, 下调基因81个)的表达活性发生变化; 与Z3相比, Z6缺少了ARS2、HMG-box和Ras-like-GTPase转录因子家族, JmjC和zf-C2H2在短暂消失后又重新在Z6中响应表达, 同时增加的还有Auxin_resp、PLN00014和HD-ZIP_N。中棉所12在受到枯萎病菌诱导后12 h开始, 转录因子的种类和数量开始减少。Z12中有27个转录因子家族的92个转录因子(上调基因17个, 下调基因75个)表达活性发生变化, 与Z6相比, 转录因子家族DEXDc开始发挥作用, 而Auxin_resp、E2F_TDP、FAR1、GAGA_bind、PLN00014、TCP、zf-Dof以及ZnF_GATA却未见进一步调控表达。而后Z24中的转录因子家族数量继续减少至23个, 表达活性发生变化的转录因子基因也减少为55个(上调基因10个, 下调基因45个); 与之前对比发现, ARS2重新开始响应, SET则被首次激活, 同时CBFB_NFYA, EIN3、H4、JmjC、R3H、SRPBCC缺失。最后在Z48中只有15个转录因子家族的30个转录因子基因(上调基因13个, 下调基因17个)表达活性发生变化。Auxin_resp、H4、SRPBCC又开始重新发挥作用, 而ANK、ARS2、DEXDc、GRAS、HSF、KNOX、MADS、MBF1、SANT、SET和TOP4c则停止进一步调控表达。
| 表1 中棉所12枯萎病菌诱导后转录因子家族的种类和数目 Table 1 Types and number of transcription factor family induced by Fusarium wilt in Zhongmiansuo 12 |
中棉所12的6个比对组中, 表达活性均发生变化的重叠转录因子家族有9个, 分别为AP2、AUX/ IAA、bZIP、BBOX、HLH、Homeodomain (HD-other family)、LRR、NAC以及WRKY。其中bZIP (59个, 13.6%)、NAC (41个, 9.5%)、AP2 (31个, 7.1%)、AUX_IAA (30个, 7.0%)、HLH (26个, 6.0%)以及WRKY (25个, 5.8%)这6个转录因子家族中参与枯萎病菌诱导响应的转录因子数量较多, 共占表达发生变化的转录因子总数的49.0%。此外参与枯萎病菌诱导响应的转录因子数量较多的转录因子家族还有SANT, 它在5个比对组中重叠, 共有30个转录因子, 占表达发生变化的转录因子总数的6.9%。
枯萎病菌诱导后, 感病品种新陆早7号响应的转录因子的数目和种类以及涉及到的基因数量均明显多于中棉所12, 共有52类转录因子家族的588个转录因子的表达活性在至少一个比对组中发生了变化。X1中有29个转录因子家族表达活性发生变化, 而后X3中转录因子家族的数量增加至45个, AAA_ 17、ANK、BRLZ、Cupin_2、DEXDc、DUF3353、DUF822、E2F_TDP、GAGA_bind、HALZ、LRR、MBF1、PLN00014、R3H、Sm-like、SpoU_Methylase、TCP、TFIIS和ZnF_GATA等19个转录因子家族开始发挥作用; 而GH64-TLP-SF、HMG-box、PHD则缺失。新陆早7号在受到枯萎病菌诱导后6 h, 转录因子家族的数量开始下降。X6中有40个转录因子家族的表达活性发生变化, 比对组X3增加了ARID、Auxin_resp、JmjC、PHD、TBP_TLF以及zf-Dof等6个转录因子家族, 同时缺少了AAA_17、ARS、BRLZ、CBFB_NFYA、Cupin_2、DUF1762、DEXDc、HALZ、MOZ_SAS、PLN00014和PPR_2等11个转录因子家族。而X12中只有35个转录因子家族的表达活性发生变化, 与之前相比CBFB_NFYA、Cupin_2、DEXDc、HALZ、PLN00014又重新开始响应, 而ARID、Auxin_resp、FAR1、PHD、R3H、SpoU_Methylase、TBP_TLF、TFIIS、zf-Dof、ZnF_ GATA则缺失。在X24中有30个转录因子家族的表达活性发生变化; 与X12相比, X24增加了3个转录因子家族ARID、PPR_2和ZnF_GATA, 缺少了ANK、CBFB_NFYA、Cupin_2、DEXDc、E2F_TDP、H4、JmjC和PLN00014。最后在X48中仅有28个转录因子家族的表达活性发生变化, ANK和H4增加; 而DUF822、GAGA_bind、PPR_2以及ZnF_GATA则缺失。
X1中有80个(13.6%)转录因子表达活性发生变化, 涉及到上调基因30个, 下调基因50个; 而后比对组X3中增加至151个(25.7%)转录因子发生变化, 涉及到上调基因28个, 下调基因123个。比对组X6中表达活性发生变化的转录因子则减少到112个(19.0%), 涉及到上调基因26个, 下调基因86个。比对组X12中同样有112个(19.0%)转录因子的表达活性发生变化, 涉及到上调基因14个, 下调基因98个。随着新陆早7号受到枯萎病菌诱导后的时间继续增加, 在比对组X24中表达活性发生变化的转录因子降低到73个(12.4%), 涉及到上调基因15个, 下调基因58个。最后在比对组X48中还有60个(10.2%)转录因子的表达活性发生变化, 涉及到上调基因15个, 下调基因45个。
新陆早7号的6个比对组中, 表达活性均发生变化的重叠的转录因子家族有18个, 分别为AP2、AUX_IAA、bZIP、BBOX、EIN3、GRAS、HD-ZIP_N、HLH、HSF、Homeodomain (HD-other family)、KNOX、MADS、NAC、SANT、SRPBCC、Sigma70 like、WRKY和zf-C2H2; 其中bZIP (70个, 11.9%)、SANT (47个, 8.0%)、AUX_IAA (42个, 7.1%)、NAC (41个, 7.0%)、AP2 (35个, 6.0%)、HLH (34个, 5.8%)以及WRKY (29个, 4.9%)这7个转录因子家族中参与枯萎病菌胁迫响应的转录因子数量最多, 占表达发生变化的转录因子总数的50.7%。该结果与中棉所12的结果相同, 推测这7个转录因子家族可能在棉花受枯萎病菌诱导后48 h内发挥重要作用。
| 表2 新陆早7号枯萎病菌诱导后转录因子家族数目和种类 Table 2 Number and variety of transcription factor family induced by Fusarium wilt in Xinluzao 7 |
对中棉所12和新陆早7号2个品种在各个比对组中转录因子家族及转录因子的整体变化分析表明(图1-A), 感病品种新陆早7号对枯萎病菌响应的转录因子家族明显多于中棉所12, 并且新陆早7号受到枯萎病菌诱导后3 h转录因子家族的数量达最多, 而中棉所12则在诱导后6 h转录因子家族才达最多。二者在6个比对组中响应枯萎病菌诱导的转录因子家族数量都呈现相似的变化趋势, 即在诱导后的1 h部分转录因子家族率先响应, 在3 h或6 h达最多, 而后随着时间的推移开始呈现下降的趋势, 诱导后48 h响应枯萎病菌诱导的转录因子家族数量降到最低水平。图1-B和1-C表明, 无论是抗病还是感病品种, 在6个比对组中, 其下调基因的数目均多于上调基因, 且响应枯萎病菌诱导的转录因子总数的变化趋势与转录因子家族的变化趋势类似, 也表现出先增加后逐渐减少的变化趋势, 转录因子的数量也是中棉所12在枯萎病菌诱导后的6 h达最多, 新陆早7号则在诱导后的3 h达到最多。
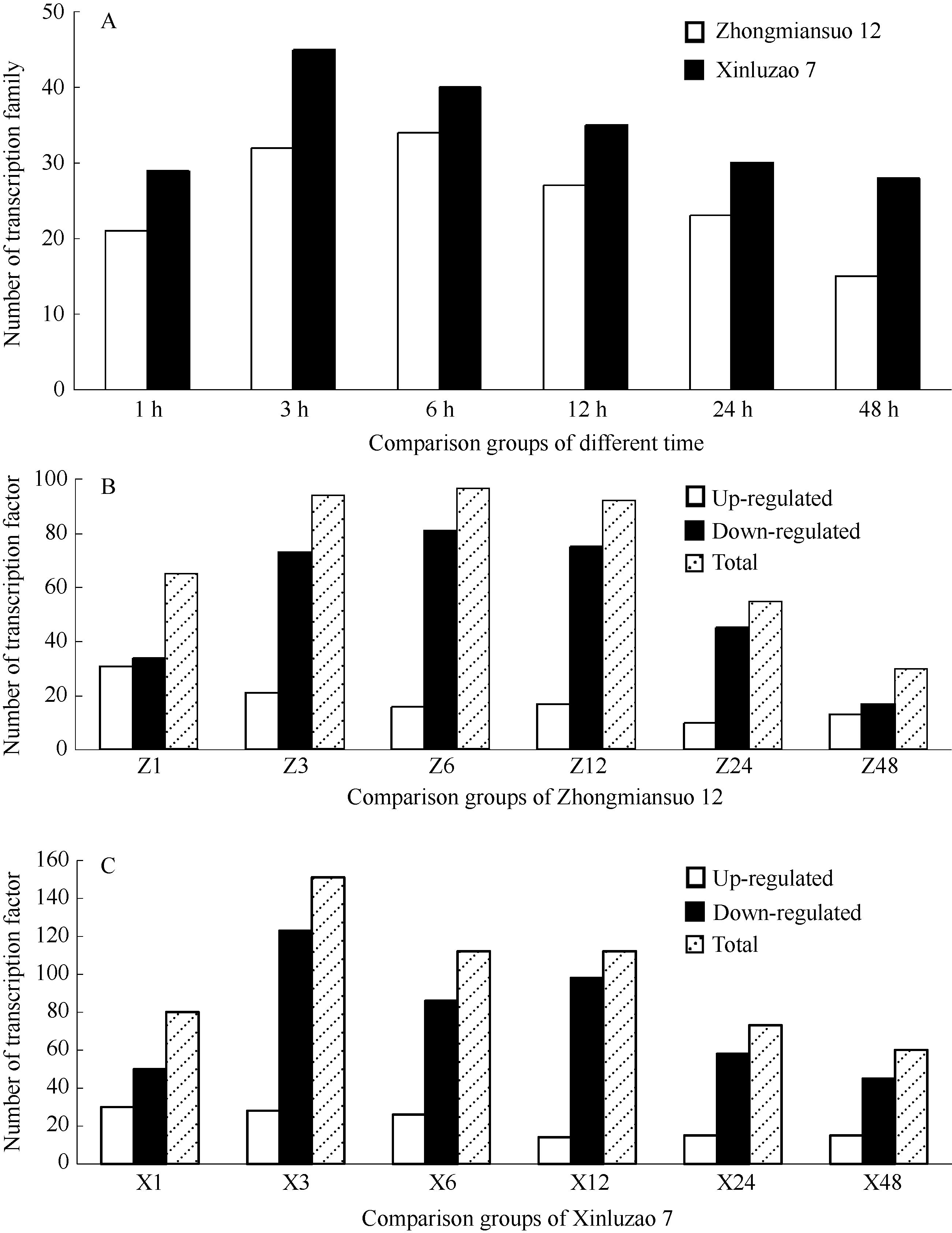 | 图1 中棉所12和新陆早7号转录因子家族及转录因子数量分析Fig. 1 Summary of transcription factor family and transcription factors of Zhongmiansuo 12 and Xinluzao 7 |
为了进一步了解转录因子在抗病品种和感病品种中的表达变化, 分别对枯萎病菌诱导后2个品种在6个比对组中表达活性均发生变化的重叠的转录因子分析表明, 中棉所12在6个比对组中重叠的转录因子有9个, 隶属于6个转录因子家族(表3), 即bZIP、Homeodomain (WUSCHEL related homeobox 4 bZIP)、MADS、NAC、SANT和SRPBCC, 占该品种表达活性发生变化的转录因子总数的2.1%, 其中枯萎病菌诱导后表达活性在6个比对组中均下调的转录因子有6个。新陆早7号在6个比对组中的重叠转录因子有31个, 隶属于17个转录因子家族(表4), 即AP2、bZIP、CBFB_NFYA、GAGA_bind、GRAS、H4、HD-ZIP_N、HSF、KNOX、MADs、NAC、PHD、SANT、SRPBCC、TOP4C、WRKY和zf-C2H2, 占该品种表达活性发生变化的转录因子总数的5.3%, 其中表达活性在6个比对组中均下调的转录因子有22个, 上调的转录因子有4个。
| 表3 中棉所12在6个比对组中的重叠转录因子 Table 3 Overlapped transcription factors of Zhongmiansuo 12 in six comparison groups |
| 表4 新陆早7号在6个比对组中的重叠转录因子 Table 4 Overlapped transcription factors of Xinluzao 7 in six comparison groups |
通过分析感、抗品种对枯萎病菌诱导响应转录因子的表达活性变化发现, 不同抗病性品种对枯萎病的响应有一定品种特异性(表5)。中棉所12和新陆早7号在受到枯萎病菌诱导后6个比对组中分别有39个和52个转录因子家族的表达活性发生变化, 其中37个转录因子家族为2个品种共有, 即ANK、AP2、ARS2、AUX_IAA、Auxin_resp、bZIP、BBOX、BRLZ、BFB_NFYA、DEXDc、E2F_TDP、EIN3、FAR1、GAGA_bind、GRAS、H4、HD-ZIP_N、HLH、Homeodomain (HD-other family)、HSF、JmjC、KNOX、LRR、MADS、MBF1、NAC、PLN00014、R3H、SANT、Sigma70 like、SRPBCC、TCP、TOP4c、WRKY、zf-C2H2、zf-Dof和ZnF_GATA。另外, 有2个转录因子家族(包含2个转录因子)是中棉所12特有, 占中棉所12在枯萎病菌诱导后表达活性发生变化的39个转录因子家族的5.1%, 包括在Z3比对组中出现的转录因子家族Ras-like-GTPase以及在Z24比对组中出现的SET, 二者仅在中棉所12受枯萎病菌诱导后3 h和24 h出现一次, 且均为下调表达。
另有15个转录因子家族(包含45个转录因子)是新陆早7号特有, 占新陆早7号在枯萎病菌诱导后表达活性发生变化的52个转录因子家族的28.8%。除仅在X1中出现的隶属于GH64-TLP-SF家族的1个转录因子为上调表达外, 其余的44个转录因子(隶属于14个转录因子家族)均为下调表达。转录因子家族AAA_17和TBP_TLF分别在X3和X6比对组中出现一次。其余的转录因子家族ARID、Cupin_2、DUF1762、DUF3353、DUF822、HALZ、MOZ_SAS、PHD、PPR_2、Sm-like、SpoU_Methylase以及TFIIS均在2个或2个以上的比对组中对枯萎病菌诱导响应。
| 表5 2个棉花品种转录因子家族的品种特异性 Table 5 Cultivar-specificity of transcription factor family of Zhongmiansuo 12 and Xinluzao 7 |
大量研究表明, 转录因子在植物与病原菌的互作中发挥着重要的调控作用。目前已知与抗病反应有关的转录因子家族主要有bZIP型、ERF型转录因子、锌指蛋白中的WRKY家族、Homeodomain蛋白、HSF蛋白等类型, 其中Homeodomain蛋白、HSF蛋白参与植物防卫反应的报道较少[24]。
本研究表明, 无论是抗病还是感病品种, 在枯萎病菌诱导后, 均有大量转录因子家族及转录因子的表达活性发生改变, 中棉所12中有39个转录因子家族的433个转录因子的表达活性发生变化, 其中上调表达的转录因子有108个, 下调表达的转录因子有325个; 新陆早7号中有588个转录因子隶属于52个转录因子家族的表达活性发生变化, 其中上调表达的转录因子有128个, 下调表达的转录因子有460个。进一步分析6个比对组发现, bZIP、SANT、AUX_IAA、NAC、AP2、HLH和WRKY这7个转录因子家族在2个品种中参与枯萎病菌胁迫响应的转录因子数量均最多, 推测其可能在棉花受枯萎病菌诱导后48 h内发挥重要作用。
综合中棉所12和新陆早7号各个比对组可以看出, bZIP家族中对枯萎病菌响应的转录因子数量最多, 分别占13.6%和11.8%。bZIP (碱性亮氨酸拉链)转录因子家族是真核生物中分布最广、最保守的一类, 仅在拟南芥中就发现75个成员。它们都含有较保守的与特异DNA序列相结合的碱性结构域和参与寡聚化作用的亮氨酸拉链区, 在植物抗病过程中, 这些结构域能够识别抗病基因启动子上核心序列为ACGT的顺式作用元件G盒、C盒、A盒等, 并与其相互作用, 调控下游许多抗病基因的表达[3]。Zhang等[25]研究发现, 小麦的转录因子TabZIP1通过依赖于乙烯/茉莉酸信号途径参与植物对条锈病的抗病反应。而辣椒中的bZIP型转录因子PPI1和CAbZIP1能在病毒和病原细菌侵染后诱导表达[26]。
此外, AP2转录因子家族在中棉所12中共有31个转录因子、在新陆早7号中共有35个转录因子响应枯萎病菌的诱导, 分别占表达活性发生变化的转录因子总数的7.1%和6.0%。大量研究发现, 植物中庞大的AP2基因家族成员广泛参与植物响应外界环境胁迫、生长发育相关的转录调控。ERF是AP2家族中的一个亚族, 它编码的蛋白能够参与植物抗病反应[27]。何兰兰等[28]研究发现, ERF-B3亚组转录因子基因GhB301, 在受到枯萎病菌诱导后, 其表达量在抗病品种中棉所12与感病品种新陆早7号中均明显增加, 但在抗病品种中表达量的增加显著高于感病品种。
除上述2个转录因子家族外, 在感、抗品种各比对组中同样占有较大比例的是WRKY家族, 它是仅存在于高等植物中的一类锌指蛋白, 可以和许多植物防卫反应基因启动子的高度保守作用元件W-box结合, 在抗病信号转导途径中起重要调控作用[29]。王瑞等[30]克隆了TaWRKY44基因, 并发现该基因可能作为一个转录抑制子参与逆境胁迫信号转导过程。另外, 拟南芥AtWRKY18[31]、水稻OSWRKY71[32]和棉花GaWRKY1[33]等都被证明与植物抗病反应有关。
本研究在中棉所12和新陆早7号中均发现13个隶属于Homeodomain家族的转录因子能够响应枯萎病菌的诱导, 它是由Homeodomain基因编码的约60个氨基酸组成的蛋白。这种蛋白可充当转录因子, 当它与基因结合时能够形成特殊结构, 调控它们的表达[24]。罗红丽等[34]克隆并鉴定了1个水稻转录因子基因OsBIHD1, 编码1个新的水稻Homeodomain蛋白。稻瘟病菌侵染和诱导因子处理可激活该基因的表达, 过量表达OsBIHD1的转基因烟草植株中组成型表达PR-1a基因, 并提高对病毒和真菌病害的抗性[35]。还有研究表明, Homeodomain转录因子OPC3通过茉莉酸信号途径参与植物对腐生病原菌的抗性[36]。
此外, 各个比对组中其他转录因子在植物中的作用也同样重要。如NAC与植物的生长发育、逆境胁迫的应答、细胞次生壁的生物合成等有关[37]; KNOX基因家族编码同源异型盒蛋白, 在植物生长发育过程中起重要调控作用[38]; bHLH转录因子家族是植物转录因子中最大的家族之一, 在植物抗逆、生长发育、生物合成及信号转导中发挥重要作用[39]。但是植物中转录因子的种类繁多, 仅拟南芥中至少有1533个编码转录因子的基因[3]。在本研究中, 各个比对组中除目前已研究的部分转录因子外, 仍有大量报道不多的转录因子, 这些转录因子的结构、功能仍需要后续的大量试验研究。
进一步分析6个比对组发现, 与诱导前相比, 枯萎病菌诱导后6个时间点表达活性均发生变化的重叠的转录因子, 中棉所12中仅有9个, 隶属于6个转录因子家族; 新陆早7号有31个, 隶属于17个转录因子家族。与马廷臣等[5]的研究相比, 重叠的转录因子数量明显偏少, 推测原因可能是在植物对枯萎病菌响应的过程中, 基因的表达存在复杂的反馈调控机制, 导致基因的表达量呈现不连续的变化, 有的基因表达量与对照相比不够显著, 因此在筛选表达活性发生变化的基因过程中被忽视。此外, 马廷臣等[5]在分析不同干旱胁迫对干旱敏感和耐旱水稻品种转录因子的表达变化的研究中, 仅有3个干旱胁迫处理, 而本研究感、抗品种受到枯萎病菌诱导后48 h共取样6次, 形成6个比对组。比对组数的增加, 加之在枯萎病菌诱导后1 h和48 h表达活性发生变化的转录因子数目都较少, 因此在筛选6个比对组中表达活性均发生变化的重叠转录因子时, 部分转录因子无法统计在内, 这可能也是造成本研究中感、抗品种重叠转录因子数目偏少的原因。
值得注意的是, 2个品种在受到枯萎病菌诱导后, 有的转录因子家族中存在的转录因子既有上调表达, 也有下调表达, 并且同一转录因子在不同比对组中表现不同的表达变化, 这种表达的差异可能是因为在受到枯萎病菌诱导后, 植物通过改变相关基因的表达水平, 提高某些生命活动物质的数量, 或者抑制另一些物质的表达, 以此调节植物体内各种生理生化活动, 以适应病原菌的胁迫, 减少植物受到的伤害。另外, 转录因子在对植物进行调控过程中细微的表达差异也可能对植物的性状起重要作用, 在差异表达基因筛选过程中, 部分基因与对照相比表达量差异不够显著而被忽略, 这部分基因仍可以作为后续研究的重点, 进一步分析其表达变化。
棉花枯萎病菌诱导后1~48 h, 抗病品种中棉所12有39个转录因子家族的433个转录因子在至少一个比对组中表达活性发生了变化; 感病品种新陆早7号则有52个转录因子家族的588个转录因子在至少一个比对组中表达活性发生变化。随着诱导后时间延长, 两品种对枯萎病菌诱导响应的转录因子家族和转录因子数量均呈现相似的先增加后降低的变化趋势; 在6个比对组中表达活性均发生变化的重叠转录因子, 中棉所12中有9个, 隶属于6个转录因子家族; 新陆早7号中有31个, 隶属于17个转录因子家族。不同抗病性品种对枯萎病的响应有较强的品种特异性, 除37个共有的转录因子家族外, 2个转录因子家族是中棉所12所特有, 15个转录因子家族是新陆早7号所特有。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|