{kind=link}
{kind=link}
软质小麦溶剂保持力关联分析
[张勇 , 张晓, 郭杰, 高德荣, 张伯桥
, 张晓, 郭杰, 高德荣, 张伯桥* ]
, 张晓, 郭杰, 高德荣, 张伯桥]
|
|


第一作者联系方式: E-mail: zy@wheat.org.cn
溶剂保持力(SRC)是软质小麦鉴定评价的重要指标。为获得与SRC关联的分子标记, 提高育种效率, 对不同硬度类型的176份品种的乳酸SRC、碳酸钠SRC、蔗糖SRC和水SRC进行SSR标记检测, 并结合其在江苏里下河地区连续3个生长季的4种SRC表型数据, 利用MLM模型进行了关联分析。以236对SSR引物共检测出1340个等位变异, 平均每个位点5.5个等位变异, 平均PIC值为0.4663。共检测到28个关联位点( P< 0.005), 单个位点可解释3.19%~21.84%的表型变异; 与乳酸SRC、水SRC、蔗糖SRC和碳酸钠SRC相关联的位点分别为13、7、6和2个; 与水SRC关联的gwm642-1D在3年中均被检测到。在这些关联位点上发现有利等位变异, 其中降低水SRC的等位变异有gwm642-A186、gwm642-A188和gwm337-A178, 降低蔗糖SRC的等位变异有gwm337-A178和gwm337-A186, 降低碳酸钠SRC的等位变异有cfa2257-A129等。这些结果为利用分子标记进行SRC辅助选择提供了重要信息。
Solvent retention capacity (SRC) is an important index for identification and evaluation of soft wheat varieties. This study aimed at identifying SRC associated markers for marker-assisted selection. One hundred and seventy-six wheat varieties (lines) in different hardness types were screened with 236 pairs of SSR primers and their SRC values of lactic acid, water, sucrose, and sodium carbonate were evaluated in three growing seasons in Lixiahe, Jiangsu Province. The association analysis was carried out using the mixed-linear model (MLM). A total of 1340 fragments were amplified on the 236 SSR loci with an average of 5.5 alleles per locus. The average polymorphism information content was 0.4663. Twenty-eight loci were identified to be associated with lactic acid SRC (13), water SRC (7), sucrose SRC (6), and sodium carbonate SRC (2) at the significant level of P < 0.005, and a single locus explained 3.19%-21.84% of phenotypic variation. Marker gwm642 associated with WSRC was detected in three years. Some favorable alleles associated with SRCs were found, such as gwm642-A186, gwm642-A188, and gwm337-A178 for reducing water SRC, gwm337-A178 and gwm337-A186 for reducing sucrose SRC, and cfa2257-A129 for reducing sodium carbonate SRC. These results are informative for marker-assisted selection on SRC properties in wheat.
小麦是重要的粮食作物, 目前年种植面积约2333万公顷, 年均总产量超过1亿吨。随着生活水平的提高, 小麦生产和育种也从追求产量转向优质高产协调发展, 从20世纪80年代中期开始, 我国就已开展小麦品质育种研究, 但当时大多数育种单位都强调强筋小麦而忽视弱筋小麦, 在弱筋小麦遗传特性、种质创新、品种选育以及快速鉴定方法等方面均缺乏研究[1, 2]。自2001年我国小麦品质区划[3]制定和发布后, 弱筋小麦开始受到重视, 宁麦9号、扬麦13和宁麦13等弱筋品种相继育成和大面积推广, 促进了我国弱筋小麦生产的发展, 但现有弱筋品种仍然存在品质性状年度间和地点间波动较大等问题, 弱筋品质总体水平尚待进一步提高[4, 5]。
溶剂保持力(solvent retention capacity, SRC)是指以含水量14%的面粉为基准, 经过离心后, 面粉所保持的溶剂质量占面粉干重的百分比, 该指标主要用于软质小麦的品质评价, 1999年通过AACC认定(AACC 56-11), 已成为美国评价软麦品质的主导方法[6, 7, 8]。检测使用的溶剂有5%乳酸溶液、5%碳酸钠溶液、50%蔗糖溶液和蒸馏水4种, 其中乳酸SRC与麦谷蛋白特性相关, 碳酸钠SRC与破损淀粉数量相关, 蔗糖SRC与戊聚糖特性相关, 而水SRC受面粉所有成分的影响, 反映了面粉的综合特性[8]。SRC主要受基因型控制[9, 10, 11, 12, 13]。大多数研究表明, 上述4种SRC值与饼干直径呈极显著负相关[7, 13, 14, 15], 可以利用SRC值对面粉的饼干品质特性进行评价。饼干为含水量低的烘焙食品, 要求面粉具有较低的吸水率、淀粉破损率和戊聚糖含量, 不同SRC值为评价饼干品质提供了更多的面粉品质特性信息[14, 16]。
关联分析是利用自然群体中的连锁不平衡, 鉴定目的性状与遗传标记或者候选基因之间关系, 从而挖掘或定位基因的分析方法[17]。2001年Thornsberry成功地将该方法应用于玉米开花期变异的研究[18]。由于关联分析研究对象为自然品种群体, 可利用材料相对广泛, 是解析复杂性状、鉴定目标性状优异等位基因的有效方法[19, 20], 迄今为止, 在玉米、水稻、小麦等作物中已有较多应用实例[21, 22, 23, 24]。
小麦品质育种过程中, 往往需对种子检测后再进行选择, 无法对育种分离世代材料进行田间直接筛选, 而且在回交或滚动回交育种过程中必须根据籽粒测定结果进行选择后再于下一季进行配组, 严重影响育种进度。2001年Campbell等[25]采用软白麦NY18× 硬白麦CC的78个重组自交系对碱水保持力进行了QTL作图, 2008年Smith和Souza等[26]曾在ASA-CSSA-SSSA年会中介绍了利用美国品种对溶剂保持力关联分析的结果, 目前, 还没有利用自然群体对4种SRC进行关联定位的报道。本研究在江苏扬州连续3个生长季种植176份小麦品种(系), 通过分子标记检测和表型分析, 筛选与SRC相关的分子标记, 为品质育种辅助选择提供参考信息。
176份不同硬度值类型的小麦品种(系)(见附表)均由江苏里下河地区农业科学研究所种质库收集提供。于2008— 2009、2009— 2010和2010— 2011年度, 连续3个生长季在江苏里下河地区农科所试验基地(江苏扬州)种植, 田间采用随机区组设计, 3次重复, 每小区4行, 每行40粒。行长1.33 m, 行距0.23 m。
收获后将籽粒样品磨粉, 参照AACC 56-11方法[6, 7], 测定5%乳酸SRC、5%碳酸钠SRC、50%蔗糖SRC和水SRC, 标准粉样品用量改为1 g [27, 28]。
选用236个SSR标记检测供试材料, 其中106个为荧光SSR标记, 引物信息从http://www.GrainGenes. org/获得。这些标记覆盖小麦全基因组不同染色体, 每条染色体有6~17个标记, 平均11.2个, 其中2A、5B和5D上有17个, 1A、2B、3B和3D上有14个, 1D上有13个, 2D上有12个, 1B上有11个, 5A、6A、4B和7B上有10个, 3A、7A和7D上有9个, 4A和4D上有7个, 6B和6D上有6个。
取苗期单株叶片, 采用改良CTAB法提取小麦基因组DNA[29]。PCR扩增程序为94℃预变性5 min; 94℃变性45 s, 65℃退火45 s, 72℃延伸55 s, 2个循环; 此后每2个循环退火温度下降2℃, 直至55℃; 保持55℃退火温度扩增25个循环; 最后72℃延伸10 min, 4℃保存。采用6%聚丙烯酰胺凝胶电泳分离普通SSR引物PCR扩增产物并经银染法检测读取数据, 采用DNA测序仪ABI3730分析荧光SSR引物PCR扩增产物, 通过GeneMapper3.0读取扩增条带。
利用SPSS16.0分析SRC数据方差及计算广义遗传力[30], 利用Powermarker V3.25分析等位变异数和多态性信息含量[31], 采用STRUCTRE 2.3.2软件[32]分析群体结构, 计算相应的Q值。在估计最佳群体亚群数目过程中, K值预设范围为2~12, 参数iterations设定为10 000, burn-in period设定为100 000, 每个K值均运行5次, 最终根据似然值最大的原则选取合适的K值[33]。利用TASSEL2.1 (http://www.maizegenetics. net/)中的MLM模型(mixed linear model)对4种SRC与标记变异分别进行关联分析[34], 并计算Kinship。当标记的P< 0.005时认为与SRC性状存在关联。
3年度试验结果表明, 4种SRC在品种间存在广泛差异, 品种间、年度间以及品种× 年度互作效应均达极显著水平(表1)。说明SRC除了品种间存在真实差异外, 还易受外部环境因素的影响。
| 表1 176份小麦品种的溶剂保持力(SRC)变异分析 Table 1 Variations of solvent retention capacity in 176 wheat varieties |
236对SSR引物在176个品种中共扩增出1340个等位变异, 平均每个位点检测到5.5个等位变异, 平均多态性信息含量PIC值为0.4663。从236个SSR引物中选取了64个不连锁的SSR标记, 利用STRUCTURE2.1软件估算了群体结构, 后验概率平均值ln P(D)从-28014.02呈抛物线状逐渐上升至-21592.32, 随K值增加而持续增大, 无法判断亚群数, 因此计算了概率对数变化率Δ K, 结果在K=5时出现明显峰值(图1), 据此确定供试材料亚群数为5。
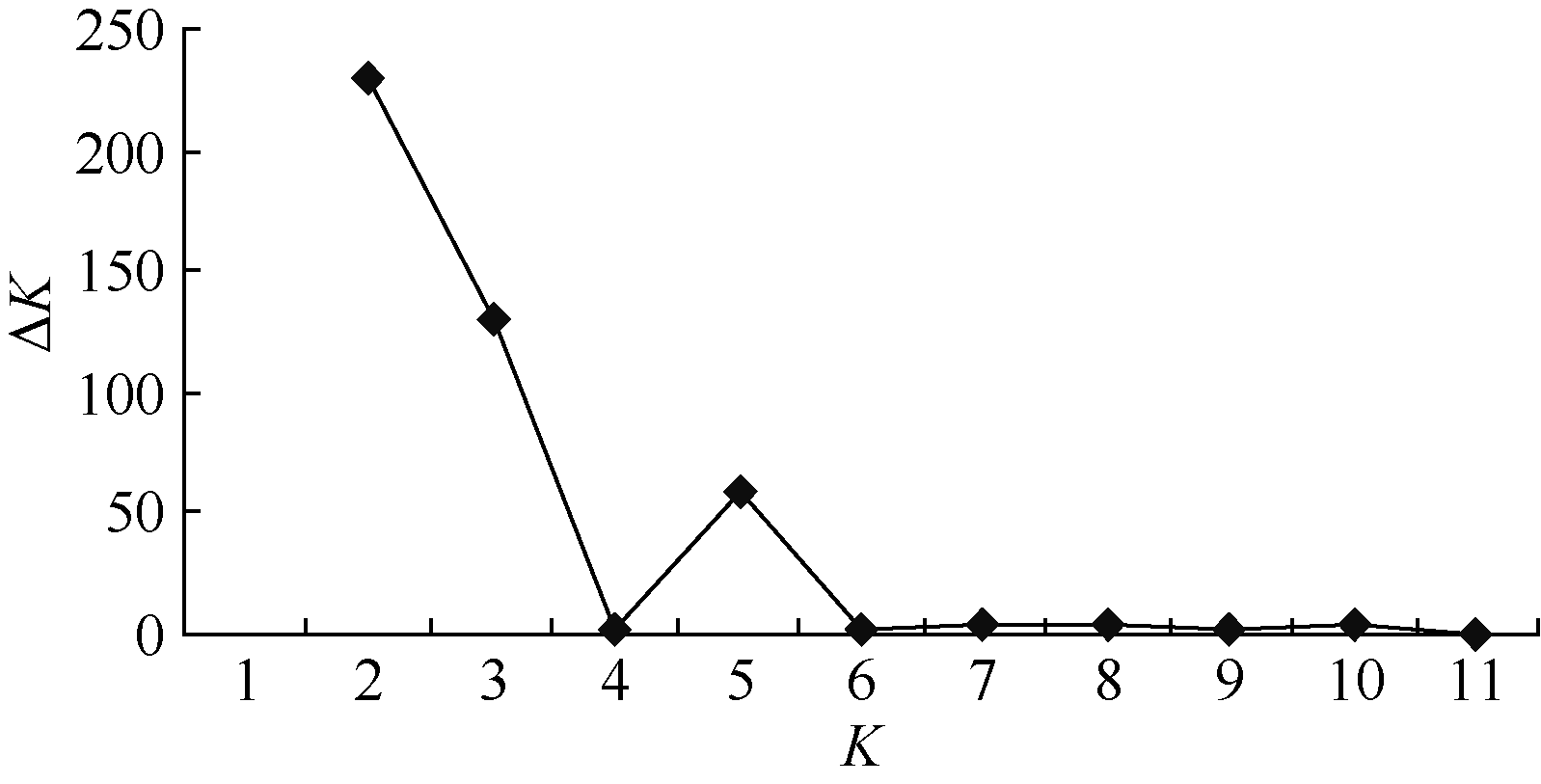 | 图1 概率对数变化率Δ KFig. 1 Rate of change in the log probability values of data between successive K-values (Δ K) |
利用236个分子标记对SRC表型数据进行关联分析, 3年度检测到28个位点与4种SRC相关联(P < 0.005), 其中13个与乳酸SRC关联、7个与水SRC关联、6个与蔗糖SRC关联, 2个与碳酸钠SRC关联, 单个位点的表型解释率为3.19%~21.84% (表2)。
| 表2 与溶剂保持力(SRC)关联的标记位点及其对表型变异的解释率 Table 2 Loci associated with solvent retention capacity (SRC) and their contributions to phenotypic variation |
位于1D的gwm642, 3年度均被检测到与水SRC极显著相关联, 且2010-2011年度还与乳酸SRC相关联, 解释表型变异的8.37%~14.42%; 位于1D的gwm337在3年中均被检测到, 与除碳酸钠SRC外的其他3种SRC相关联, 其表型贡献率为12.29%~ 15.75%; 位于7A的cfa2257在2010— 2011年度被检测到, 与蔗糖SRC和碳酸钠SRC相关联, 分别解释表型变异的4.22%和5.71%; 其他位点均在某一年被检测到, 且只与一种SRC相关联。
根据3年度关联定位结果, 对gwm642-1D、gwm337-1D和cfa2257-7A等标记的有利等位变异进行了挖掘。gwm642-1D与水SRC关联, 其等位变异A186和A188在3年中均表现降低水SRC, 其效应值在-0.55%至-4.57%范围内; 等位变异A174在2010— 2011年度的效应为3.42%, 但在另外两年为较小的负向效应; 等位变异A202在2009— 2010年度的负向效应较小, 而2008— 2009和2010— 2011年度均表现增加水SRC, 效应值分别为1.94%和2.42% (图2-A)。
携带等位变异gwm642-A188的品种共51个, 按照硬度值分类为软质类型36个, 中硬类型11个, 硬质类型4个, 包括了国内现已育成的主要弱筋品种, 如宁麦9号、扬麦9号、扬麦13、扬麦19、扬辐麦3046、扬辐麦5号、郑麦004、皖麦18、徐州20、绵阳26、绵阳31、石家庄8号等。该等位变异的水SRC平均值为0.8532 (2008— 2009)、0.7597 (2009— 2010)和0.7121 (2010— 2011), 在不同等位变异中是均值最低的。
携带等位变异gwm642-A202的品种共16个, 包括9个硬麦类型, 如藁城8901、中优16、济麦20、鲁麦2号、安农8455、安农2号、扬麦4号、宁麦11等。该等位变异的水SRC平均值分别为0.8997 (2008— 2009)、0.7867 (2009— 2010)和0.7821 (2010— 2011), 在不同等位变异中处于较高水平。
携带等位变异gwm642-A186和A174的品种一半为软麦类型, 另一半为硬麦类型, gwm642-A186的水SRC平均值3年度分别为0.8748、0.7773和0.7317, 在不同等位变异中处于中等偏低水平; 而gwm642- A174的水SRC平均值3年度分别为0.8788、0.7892和0.7920, 在不同等位变异中处于中等偏高水平。
标记gwm337-1D 3年度分别与3种SRC关联, 其等位变异gwm337-A178在3年中均表现负效应, 对水SRC、乳酸SRC和蔗糖SRC的效应值分别为-1.50%、-2.36%和-3.27%; 等位变异gwm337-A168 在2008— 2009年度表现降低水SRC, 效应值为-1.0%, 在2009— 2010年度对乳酸SRC的效应值几乎为0, 在2010— 2011年度对蔗糖SRC表现负效应(-0.5%); 该位点的A182、A184、A188等位变异对3种SRC均为正效应, 3年检测结果一致, 效应值范围为2.0%~ 4.7%; 等位变异gwm337-A186在2010年对乳酸SRC的效应值为0.54%, 在2010— 2011年度则对蔗糖SRC表现负效应, 其值为-3.0% (图2-B)。
在cfa2257-7A位点, 等位变异A129在2010— 2011年度表现降低蔗糖SRC和碳酸钠SRC, 其效应值分别为-1.1%和-2.1%; 等位变异A131则对蔗糖SRC和碳酸钠SRC表现增效, 其效应值分别为1.5%和2.9% (图2-C)。
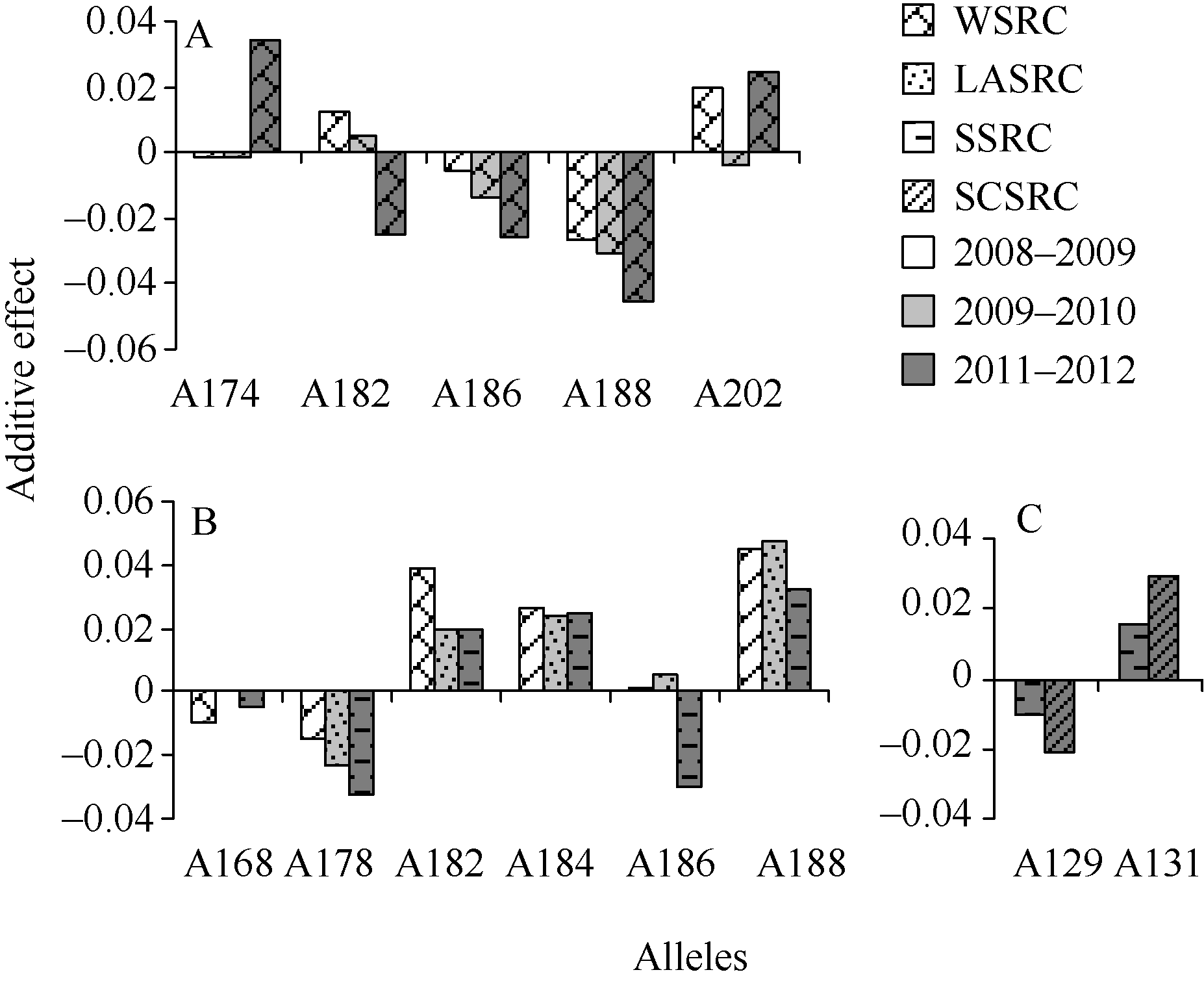 | 图2 标记gwm642-1D (A)、gwm337-1D (B)和cfa2257-7A (C)不同等位变异的表型效应不同花纹图形表示不同类型SRC, 不同背景颜色表示不同环境(年度)。WSRC: 水SRC; LASRC: 乳酸SRC; SSRC: 蔗糖SRC; SCSRC: 碳酸钠SRC。Fig. 2 Phenotypic effects of alleles on loci gwm642-1D (A), gwm337-1D (B), and cfa2257-7A (C)SRC type and environment (growing season) are distinguished by graphic patter and background color, respectively. WSRC: water SRC; LASRC: lactic acid SRC; SSRC: sucrose SRC; SCSRC: sodium carbonate SRC. |
本研究通过3年度的关联分析定位了28个与SRC极显著的标记(P < 0.005), 除1B、3D、5A、5B、6B和6D外, 其余染色体上均有标记被关联到。而这些被关联的标记大多仅一年被检测到, 只有2个标记3年度均被检测到, 其中一个标记与水SRC相关联(gwm642-1D), 另一个在不同年度分别与不同的SRC相关联(gwm337-1D)。SRC受基因型控制, 但环境对其影响较大[8, 10, 36, 37]。尽管在试验过程中从田间种植管理到收获等各方面均力求一致, 但不同年度间的气候条件存在差异, 从而影响了定位结果的稳定性。此外, 由于4种SRC间存在较高的相关性[36], 可能某些位点能同时影响多个SRC性状(表3)[26]。本文所定位的gwm337-1D在3年度与不同类型SRC极显著相关联, 推测该位点可能与多个SRC性状有关, 而由于不同年份间不同SRC的变幅不同, 因此可能出现上一年度部分SRC被极显著关联到而到下一年度却又与另一种SRC相关联的现象, 此定位结果还有待进一步的验证。由于本研究为同一地点3年度试验, 为使定位的结果能够更为准确, 应增加试验地点, 多年多点的品质检测结果将能更好地验证和补充已有结果的准确性。
对gwm642-1D不同等位变异比较发现, A188主要存在于软麦品种, 即在该等位变异存在的情况下品种应该大多表现出软麦的品质特性, 但仍然会存在一些硬麦类型品种, 因此利用该标记对SRC进行辅助筛选时还要考虑品种的硬度表现。此外, 对参试品种分析发现有些弱筋品种存在A188等位变异但硬度值偏高, 如郑麦004, 其硬度值为61, 3年度水SRC均较低, 分别为0.7447、0.7515和0.7129, 徐麦25其硬度值为52, 3年度水SRC也较低, 分别为0.8365、0.7472和0.7255。对于这类中等硬度类型的品种, 仍然可以通过标记进行辅助筛选, 但须结合除硬度外其他品质指标进行综合评价。
Smith和Souza等[26]采用关联分析的方法定位出一部分与SRC等紧密连锁的分子标记(表3), 其结果表明2B上存在着控制4种SRC的相关位点。本研究选用的位于2B的引物有14个, 部分与其所用引物相同, 部分在连锁图上与其所用引物相近。Smith和Souza等所定位的标记gwm429和barc98在本试验中3年度关联结果均未达极显著水平, 而本研究在0.005显著水平下关联到位于2B的标记仅有wmc361, 该标记2008— 2009年度与乳酸SRC极显著相关联, 与Smith和Souza等的定位的结果相距较远。马庆[38]以RIL群体对SRC进行过QTL定位, 检测到1个与水SRC相关的QTL (5DS)、2个与乳酸SRC相关的QTL (5D和6D)和1个与碳酸钠SRC相关的QTL (4D)。本研究使用了与其相同或在连锁图上相近的分子标记, 结果在4D上未检测到与碳酸钠SRC相关联的标记, 而是在7A和7D上定位到2个; 与乳酸SRC极显著相关联的标记有13个, 其中2010— 2011年度在5D上检测到与乳酸SRC极显著相关联的标记gwm174, 与马庆定位标记区间相距较远, 6D上未检测到与乳酸SRC相关联的标记; 控制水SRC的标记主要位于1A、1D、3B和7D上, 其中1D上存在着较为稳定的控制水SRC的位点。本研究定位结果与其他已有研究结果间存在的差异可能与所采用的定位群体材料和定位方法的不同有关。
| 表3 与溶剂保持力(SRC)关联的2B染色体标记均方 Table 3 Mean square terms for association of solvent retention capacity (SRC) data with chromosome 2B markers |
一地三年试验共检测到28个与SRC极显著相关联的标记, 其中与乳酸SRC关联的位点最多, 有13个, 与碳酸钠SRC关联的位点最少, 仅有2个, 单个位点的表型解释率为3.19%~21.84%, 其中gwm642-1D在3年中均被检测到与水SRC极显著相关联。在1D上存在较为稳定的控制SRC的位点。等位变异gwm642-A186、A188和gwm337-A178可降低水SRC值, 等位变异gwm337-A178和A186可降低蔗糖SRC值, cfa2257-A129可降低碳酸钠SRC值。
| 附表 试验品种(系)的硬度值及分类 Supplementary table Hardness and the category of wheat varieties/lines tested |
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|