
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
基于生物量的油菜越冬前植株叶片空间形态结构模型
[张伟欣1, 2  , 曹宏鑫
, 曹宏鑫2, * , 朱艳1, * , 刘岩2 , 张文宇2 , 陈昱利2 , 傅坤亚1, 2 ]
, 曹宏鑫, 朱艳, 刘岩|
|


第一作者联系方式: E-mail: nkyzwx@126.com
油菜越冬前的形态建成是油菜苗后期乃至整个生长中、后期的物质基础, 叶片是该期最重要的营养器官。为了明确油菜植株的形态结构要素与器官生物量的关系, 以3个甘蓝型油菜品种为材料, 于2011—2013年分别设置品种和肥料试验、品种、肥料和密度试验、品种试验, 越冬前测定油菜植株不同叶位叶片形态指标, 分析油菜主茎叶片形态参数与叶片干物重的关系, 构建了基于生物量的油菜越冬前植株叶片空间形态结构模型。建模后以独立试验数据检验, 除短柄长、叶切角和叶弦角、不施肥品种叶片干物重分配系数值(partitioning coefficient of leaf blade dry weight, CPLB)误差较大外, 油菜越冬前植株叶片空间形态结构模型观察值与模拟值一致性较好, 所建模型可靠性较高, 具有一定的解释性。
Rapeseed morphogenesis at pre-overwintering stage is the basis of growth and development of rapeseed in whole growth stage, and the leaf blades are important vegetative organ in this stage. To quantify the relationships between rapeseed plant architecture indices and the corresponding organ biomass, we used three cultivars including (V1) Ningyou 18 (conventional variety), (V2) Ningyou 16 (conventional variety), and (V3) Ningza 19 (hybrid variety) in the field experiments, and designed treatment of variety-fertilizer, variety-fertilizer-density, and variety tests in 2011-2012 and 2012-2013, with three fertilizer levels of no fertilizer, normal fertilizer (N, P2O5, K2O are 90 kg ha-1), and high fertilizer (N, P2O5, K2O are 180 kg ha-1), and three density levels of D1(6×104 plant ha-1), D2(1.2×105 plant ha-1), and D3(1.8×105 plant ha-1). Morphological indices were determined at pre-overwintering stage, the biomass-based rapeseed aboveground structure model was established with morphological indices, and the relationships between leaf blade indices and leaf blade biomass were analyzed. The models were verified using independent experiment data in 2011-2012, and 2012-2013, showing that the simulated values from the rapeseed plant leaf space morphological structure models, such as leaf blade length, leaf blade width, leaf blade bowstring length, leaf blade petiole length, and leaf blade angle from 2011 to 2012 were goodness of fit to observed values, and their da values and RMSE values were -0.231 cm, 2.102 cm ( n=63); -0.273 cm, 0.484 cm ( n=63); -0.343 cm, 1.963 cm ( n=63); 0.412 cm, 2.095 cm ( n=36); -0.635 cm, 1.006 cm ( n=27); 4.421°, 14.734° ( n=63); 6.642°, 21.817° ( n=63), respectively. The correlation between observation and simulation in the morphological indices were significant at P<0.001, but the dap values were less than 5% for the leaf blade length and the leaf blade bowstring length, which indicated that these models’ accuracy is high. The simulated values of the models had better consistency and better reliability with the observed values at pre-overwintering stage except for petiole length of the short-petiole leaves, leaf tangent angle, leaf bowstring angle, and the CPLB (partitioning coefficient of blade dry weight) under the condition of no fertilizer.
油菜是世界四大油料作物之一, 在世界各地都有大面积种植, 常年种植面积约2.43亿公顷[1]。功能-结构油菜模型研究对油菜育种中的株型选择及油菜栽培中的株型调控都具有重要的理论和应用价值。以生物量为基础将油菜生长模型和形态结构模型结合, 是建立功能-结构油菜模型的重要方法之一。随着作物模型的发展, 国内外有关油菜作物模拟研究得到广泛关注。Kiniry等基于EPIC模型[2]构建了EPR95[3]油菜生长模型, Peterson等[4]利用DAISY (differential algebra for identifiability of systems)建立了评价油菜生长发育的DAR95 (DAISY- Rape)[5]模型, Habekotte[6]借鉴LINTUL (light interception and utilization simulator) 建立了LINTUL- BRASNAP模型[7], Gabrielle等[8]基于CERES (crop- environment resource synthesis)建立了油菜CERES- Rape模型, Robertson等[9]把APSIM (agricultural production system simulator)应用于油菜, 形成了油菜生长发育模拟模型APSIM-Canola; Zhang等[10]、刘洪等[11]、廖桂平等[12, 13]、刘铁梅等[14]、胡立勇等[15]、曹宏鑫等[16]和汤亮等[17]都分别建立了油菜生长模型。与生长模型相比, 国内外关于油菜形态结构模型的研究报道较少。Groer等[18]和Mü ller等[19]以油菜为对象, 将生长模型LEAFC3-N与FSPMs (functional-structural plant model)结合, 建立了油菜功能-结构模型。该模型可根据输入的气象参数计算每张叶片碳同化, 使形态结构模型可响应环境变化, 但这种响应以叶面积为纽带。Jullien等[20]利用GreenLab建立了基于“ 源-库” 关系的油菜分枝与花扩展模型, 但尚未涉及油菜植株的其他器官。廖桂平等[21]和欧中斌[22]分别采用基于参数L-系统, 以油菜花朵和花序为对象, 建立了油菜花朵、花序的生长可视化模型。岳延滨等[23]综合油菜各器官形态生长过程与主要环境影响因子之间定量关系, 构建了基于生长度日的油菜主茎叶片、分枝叶片、主茎、分枝、花柄、花蕾及角果的形态结构模型。赵丽丽等[24, 25]也提出了基于形态特征参数的油菜花序三维形态数学模型以及交互式设计方法。本研究旨在通过2011— 2012和2012— 2013年油菜越冬前田间试验数据, 借鉴水稻地上部形态结构模型研究思路[26], 拟建立基于生物量的油菜越冬前植株叶片空间形态结构模型, 以期为今后油菜其他生长阶段形态结构模型研究提供借鉴和参考。
甘蓝型油菜品种宁油18 (V1)(常规品种)、宁油16 (V2)(常规品种)和宁杂19 (V3)(杂交品种)均由江苏省农业科学院经济作物研究所育成。V1为越冬半直立品种, 植株高度中等, 分枝部位高, 株型较紧凑; V2为越冬半直立品种, 株高中等, 分枝上举, 株型紧凑; V3为幼苗半直立品种, 叶片宽大, 叶色浅绿, 叶缘锯齿状, 匀生分枝类型。
于2011— 2012年在江苏省农业科学院院部实验农场, 采用裂区设计, 主区为施肥, 设施肥(纯氮180 kg hm-2、P2O5 120 kg hm-2、K2O 180 kg hm-2和硼沙15 kg hm-2)和CK (不施) 2个水平, 副区为品种(V1、V2和V3), 共6个处理, 3次重复, 18个小区, 随机排列, 小区面积为7.0 m × 5.7 m= 39.9 m2。行距40 cm, 株距17~20 cm。每小区栽13行, 小区间空一行。于2011年10月15日播种, 11月4日移栽。
施肥区基肥∶ 苗肥∶ 腊肥=5∶ 3∶ 2纯氮, 共基施复合肥(N + P2O5+ K2O) (质量分数≥ 25%) 16.3 kg, 补施过磷酸钙(质量分数12%) 7.3 kg、农用硫酸钾(质量分数33%) 6.4 kg。苗肥2.12 kg、腊肥1.41 kg均为尿素(总N质量分数≥ 46.2%)。其他田间管理按大田管理方式进行。
于2012— 2013年在江苏省农业科学院院部实验农场, 采用裂区设计, 主区为施肥, 分别设N0(不施肥)、N90(纯氮、P2O5、K2O各90 kg hm-2)和N180(纯氮、P2O5、K2O各180 kg hm-2) 3个水平, 副区为品种与密度, 供试品种为V1和V3, 各设每公顷单株6× 104(D1)、1.2× 105(D2)和1.8× 105(D3) 3个密度水平, 共18个处理, 54个小区, 随机排列, 重复3次。小区面积3.99 m× 3.50 m = 13.97 m2, 行距42 cm, 株距按密度计算, 每小区栽9行, 2小区间空1行。于2012年10月8日播种, 11月9日移栽。
基肥∶ 腊肥∶ 薹肥=5∶ 3∶ 2, 并于抽薹后叶面喷施硼沙15 kg hm-2。其他栽培措施同高产大田管理。
于2012— 2013年在江苏省农业科学院院部实验农场, 采用随机区组设计, 3个品种为V1、V2、V3, 施肥和密度统一采用试验2的N90(纯氮、P2O5、K2O各90 kg hm-2)和D2水平, 共3个处理, 随机排列, 重复3次。小区面积7.98 m × 3.50 m = 27.93 m2, 行距42 cm。其他同试验1.3。
土壤为黄棕土, 含有机碳13.8 g kg-1、速效氮58.95 mg kg-1、速效磷29.25 mg kg-1、速效钾109.05 mg kg-1, pH 7.84。
自三叶期标记叶位, 选取移栽前每品种长势相近的5株, 移栽后每处理长势一致的3株, 测定主茎各叶位叶片的长和宽、叶柄长和叶弦长, 以及茎叶夹角等。测后按叶位将叶片与植株分离, 并装入牛皮纸袋, 105℃下杀青30 min, 80℃烘至恒重后称量。
1.5.1 叶片测量 叶长为叶片伸直状态下由叶尖至叶基部的长度; 叶宽为最大叶宽值(位于叶片中部), 多次测量, 取平均值; 随着叶片的伸长, 由于重力等作用, 叶片发生形变, 向下弯成一个弧面, 从叶柄基部到叶尖的空间直线距离称为叶弦长, 多次测量取其平均值; 叶柄长为从叶片在主茎上的着生点至叶基部的直线距离。
1.5.2 角度测量 采用量角器直接测量叶柄与主茎的夹角为叶切角, 多次测量取其平均值; 采用量角器直接测定叶弦与主茎的夹角为叶弦角, 多次测量取其平均值。
采用SPSS Ver.19.0与Microsoft Excel 2007统计软件分析试验数据。2011— 2012年不同品种和施肥水平的部分数据用于建模及参数确定; 2011— 2012年建模以外数据和2012— 2013年独立试验资料用于模型测试和检验。
采用根均方差RMSE (root mean square error)、相关系数(r)、平均绝对误差(da)、平均绝对误差(da)与观察值平均数的比值(dap), 对模型精度进行统计分析[27]。同时, 绘制观测值与模拟值1∶ 1的关系图直观显示模型的精度和可靠性。RMSE、da与dap的值越小, R2值越大, 模拟值与实测值的一致性越好, 模拟值和实测值之间偏差越小, 即模型的模拟结果越准确、可靠。
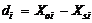
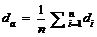
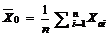
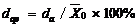
其中, i为样本数, Xoi为观测值, Xsi为模拟值, n为样本容量。
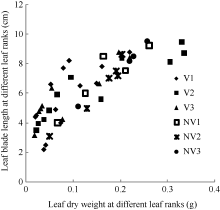
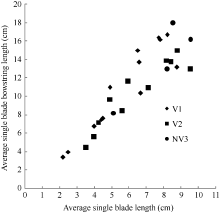
2.1.1 叶片长度模型 叶片是油菜植株最重要的光合作用器官, 而生物量是叶片生长的物质基础。2011— 2012年试验数据表明, 油菜越冬前植株主茎不同叶位叶长随其干物重的变化大致呈直线上升趋势(图1)。
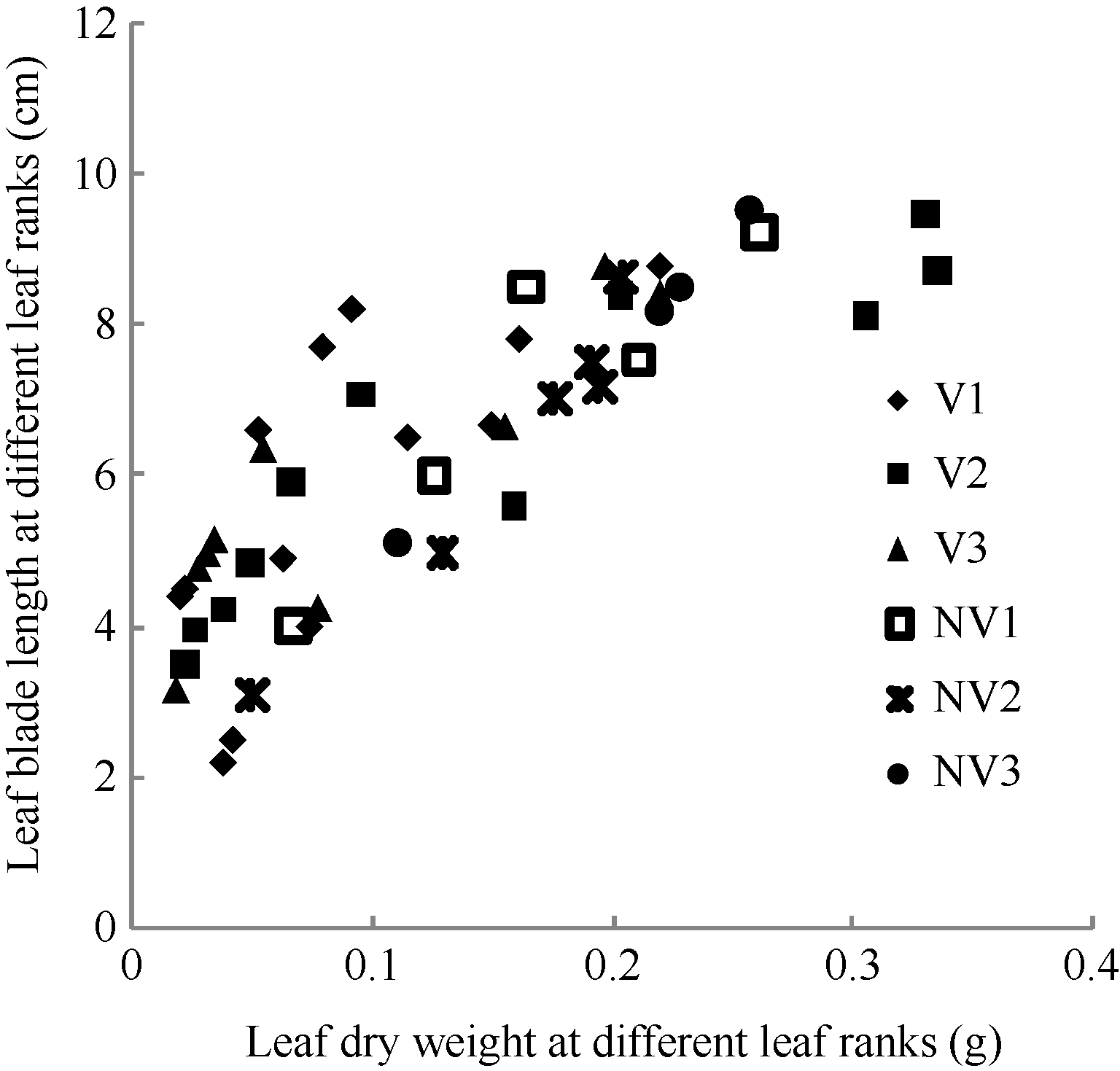 | 图1 2011-2012年不同处理叶长随叶干物重变化规律V1: 宁油18施肥; V2: 宁油16施肥; V3: 宁杂19施肥; NV1: 宁油18不施肥; NV2: 宁油16不施肥; NV3: 宁杂19不施肥。Fig. 1 Changes in the average single leaf blade length with the average single leaf dry weight for different treatments in 2011-2012V1: Ningyou 18 with fertilizer; V2: Ningyou 16 with fertilizer; V3: Ningza 19 with fertilizer; NV1: Ningyou 18 with no fertilizer; NV2: Ningyou 16 with no fertilizer; NV3: Ningyou 19 with no fertilizer. |
因此, 根据油菜越冬前主茎不同叶位叶长与对应叶干重之间的定量关系, 油菜出苗第y天主茎第j叶位叶片长度LLj(y) (cm)可表示如下:
LLj(y) = DWLBj(y) RLWj(y) (1)
DWLBj(y) = CPLBj(y) PDWSP(y) (2)
DWSP(y) = MDWSP(y) PSDWSP(y) (3)
MDWSP(y) = DWCP(y)/DES (4)
式(1)至(4)中, DWLBj(y)表示出苗第y天第j叶干物重(g); RLWj(y)表示出苗第y天第j叶叶长与该叶干物重的比值, 即比叶长重(cm g-1); CPLBj(y)表示出苗第y天第j叶干物重与该叶所在单株地上部干物重的比值, 即叶片干物重分配系数(g g-1); DWSP(y)表示出苗第y天单株干物重(g plant-1); MDWSP(y)表示出苗第y天单株地上部干物重平均值(g plant-1), SDWSP(y)表示出苗第y天每株地上部干物重标准误差(可通过试验确定)(g plant-1); DWCP(y)表示出苗第y天单位面积地上部干物重(g m-2), 可由试验确定或生长模型获得; DES是单位面积株数(plant m-2)。
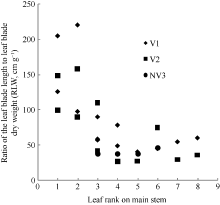
2011— 2012年试验数据表明, 不同处理油菜比叶长重随叶位变化近似于S曲线(图2), 其相关系数r=0.720 (n=28, R(26, 0.01)=0.479, P< 0.01)和R2=0.518, 模型参数及显著性测验见表1, 方程F值、t值和各参数均达P< 0.001水平显著性。因此, 不同品种和施肥处理油菜比叶长重RLWj(y) (cm g-1)随叶位变化模型可表述如下:
RLWj(y)=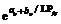
式(5)中, LPjy是油菜出苗第y天主茎第j叶的叶位, a0, b0为参数。
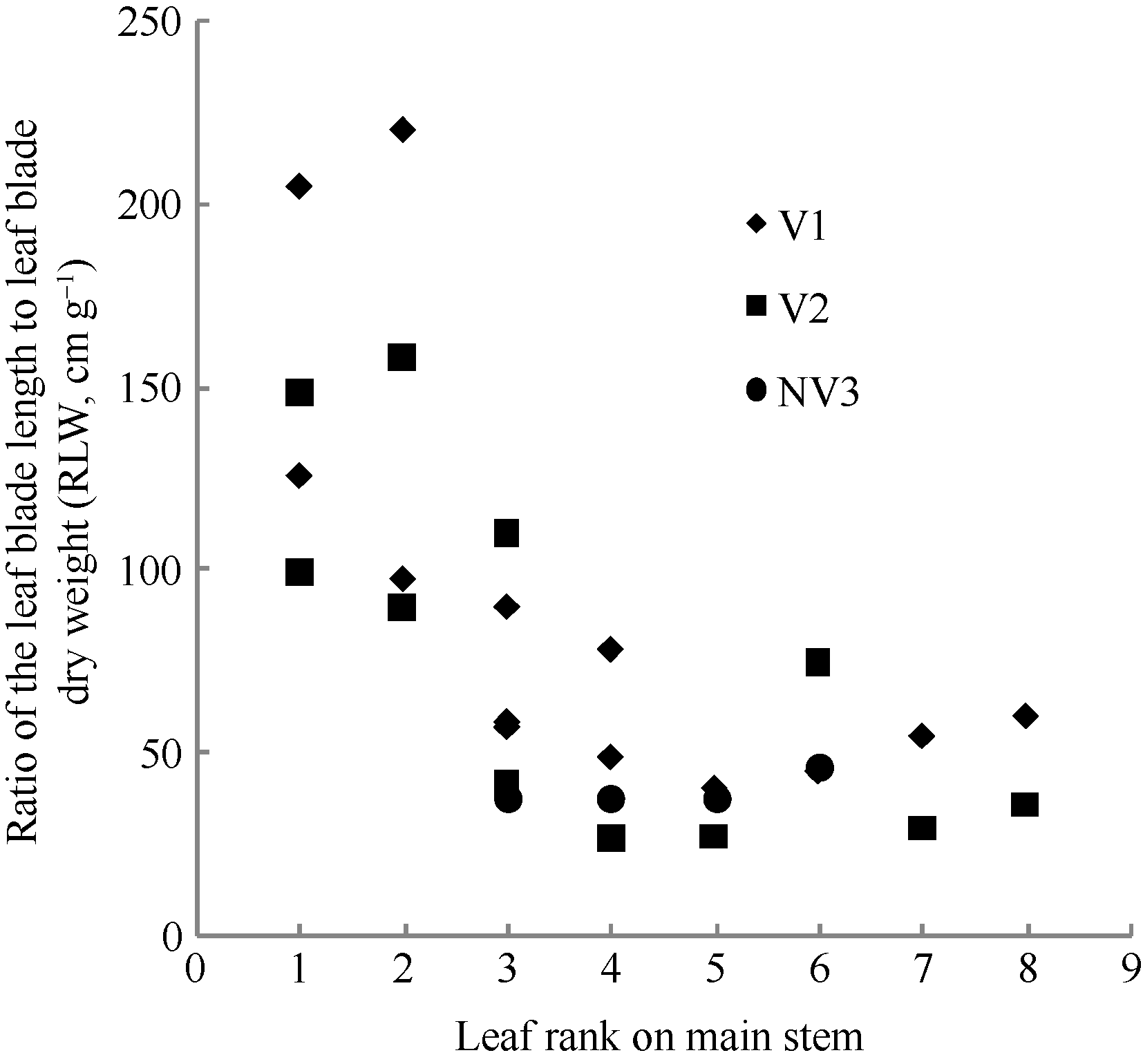 | 图2 2011-2012年不同处理比叶长重随叶位的变化规律Fig. 2 Changes in RLW values of different treatments with leaf rank on main stem in 2011-2012 |
| 表1 模型参数的确定和显著性检验 Table 1 The determination of model parameter and significance test |
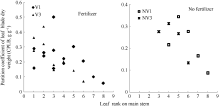
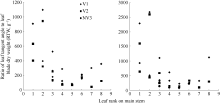
2011— 2012年试验表明, 不同品种施肥与不施肥水平下, 干物重分配系数随叶位均呈指数变化(图 3), 施肥品种相关系数r=0.695 (P< 0.001, n=22, r(20, 0.001)=0.652), R2=0.483; 不施肥品种相关系数r=0.750 (P< 0.05, n=9, r(7, 0.05)=0.666), R2=0.562; 模型参数及显著性检验见表2。方程F值和t值分别达到P< 0.001和P< 0.05显著水平, A1和A2均达P< 0.001显著水平。因此, 不同品种施肥与不施肥水平下, 干物重分配系数CPLBj(y) (g g-1)随叶位变化模型可分别表述如下:
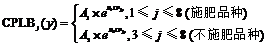
式(6)中, LPjy是油菜出苗第y天主茎第j叶的叶位, A1、B1、A2、B2均为参数。
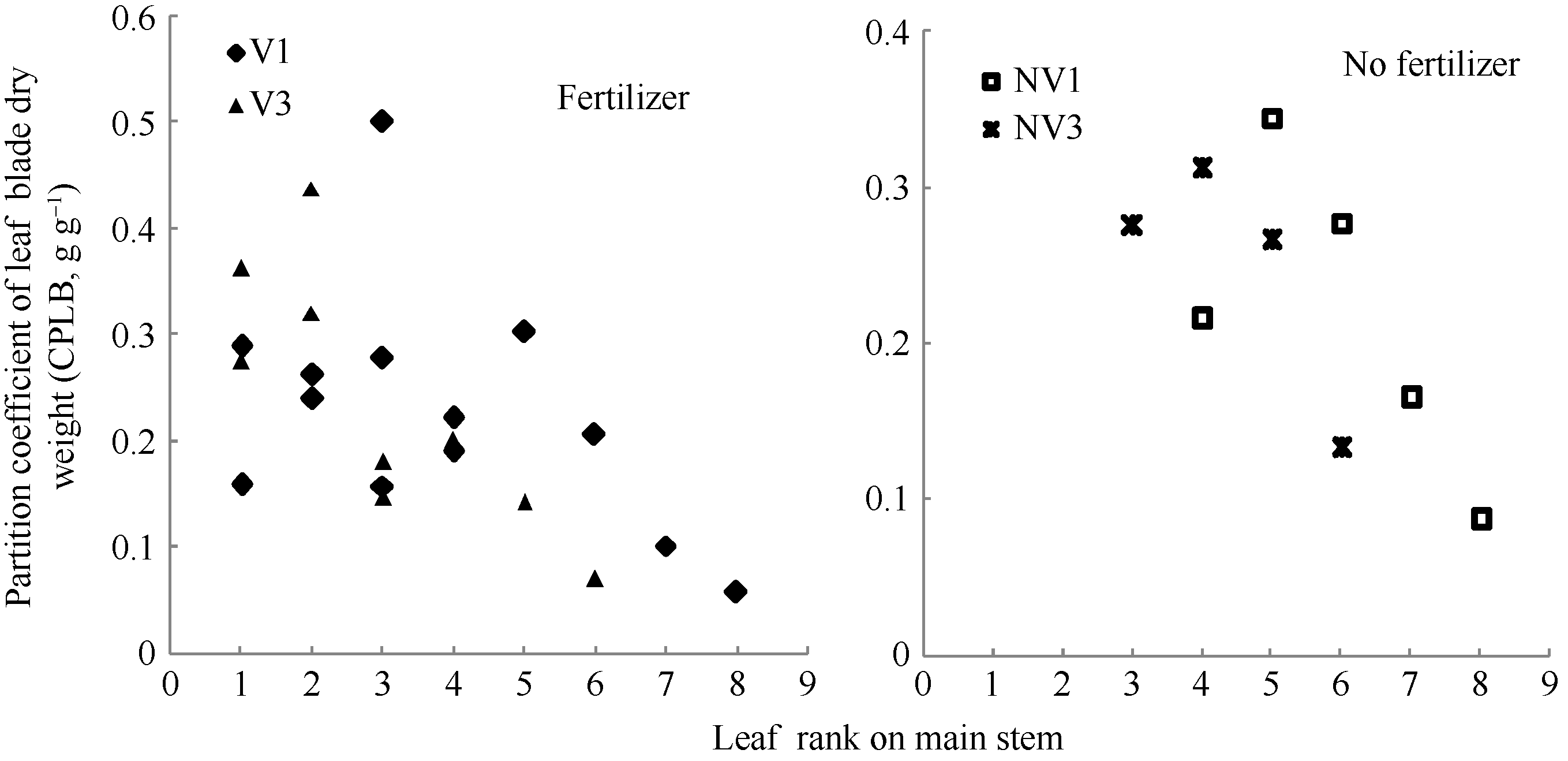 | 图3 2011-2012年不同处理CPLB随叶位的变化规律Fig. 3 Value of the CPLB of different treatments with leaf rank on main stem in 2011-2012 |
| 表2 模型参数的确定和显著性检验 Table 2 Determination of model parameter and significance test |
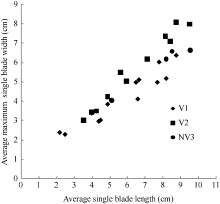
2.1.2 最大叶宽模型 2011— 2012年试验表明, 不同品种与施肥处理油菜最大叶宽随叶长变化呈线性函数增长(图4)。其相关系数r=0.930 (P< 0.001, n=28, r(26, 0.001)=0.588), R2=0.864, 模型参数及显著性测验见表1, 方程的F值达到P < 0.001显著水平, t值和a1达到P < 0.001显著水平。因此, 不同品种与施肥处理油菜最大叶宽LWj(y) (cm)随叶长变化模型可表述如下:
LWj(y)=a1LL j(y)+b1, 1≤ j≤ 8 (7)
式(7)中, LLj(y)是油菜出苗第y天主茎第j叶的叶长, a1、b1为参数。
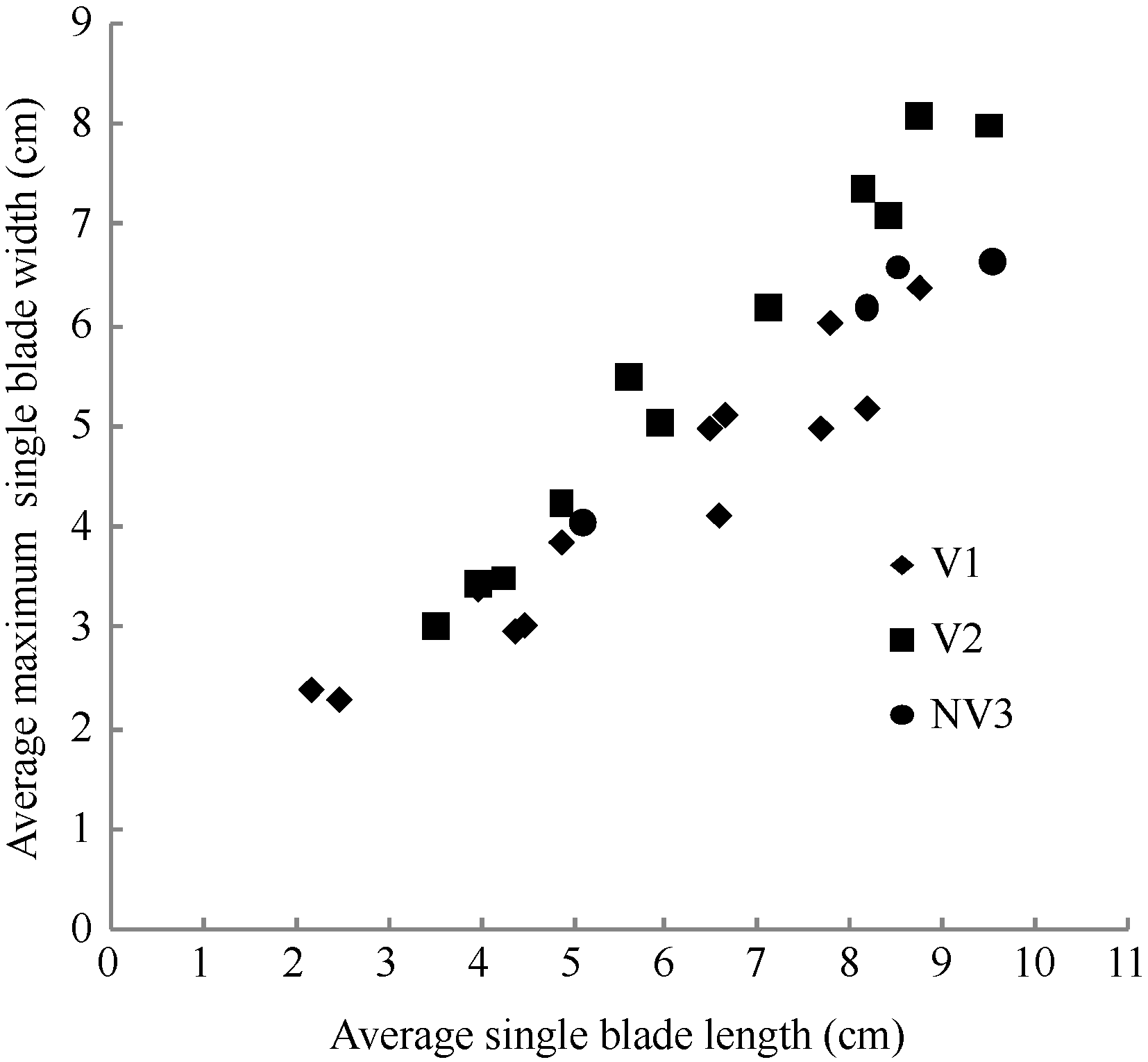 | 图4 2011-2012年不同处理最大叶宽随叶长变化规律Fig. 4 Changes in values of the maximum single blade width of different treatments with leaf blade length for three cultivars in 2011-2012 |
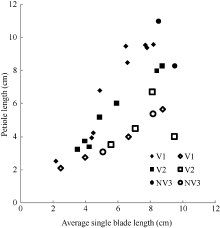
2.1.3 叶弦长模型 叶弦长是叶片弯曲度的度量, 因此, 最大叶弦长LBBLj(y)max=LLj(y)。2011— 2012年试验数据表明, 不同处理油菜叶弦长随叶长变化亦呈线性函数(图5)。其相关系数r=0.903 (P< 0.001, n=28, r(26, 0.001)=0.588), R2=0.816, 模型参数及显著性测验见表1, 方程的F值达到P< 0.001显著水平, t值和a2达到P< 0.001显著水平。因此, 不同品种与施肥处理油菜叶弦长LBBLj(y) (cm)随叶长变化模型可表述如下:
LBBLj(y)=a2[LLj(y)] - b2, 1 ≤ j≤ 8 (8)
式(8)中, LLj(y) (cm)是油菜出苗第y天主茎第j叶的叶长, a2、b2均为参数。
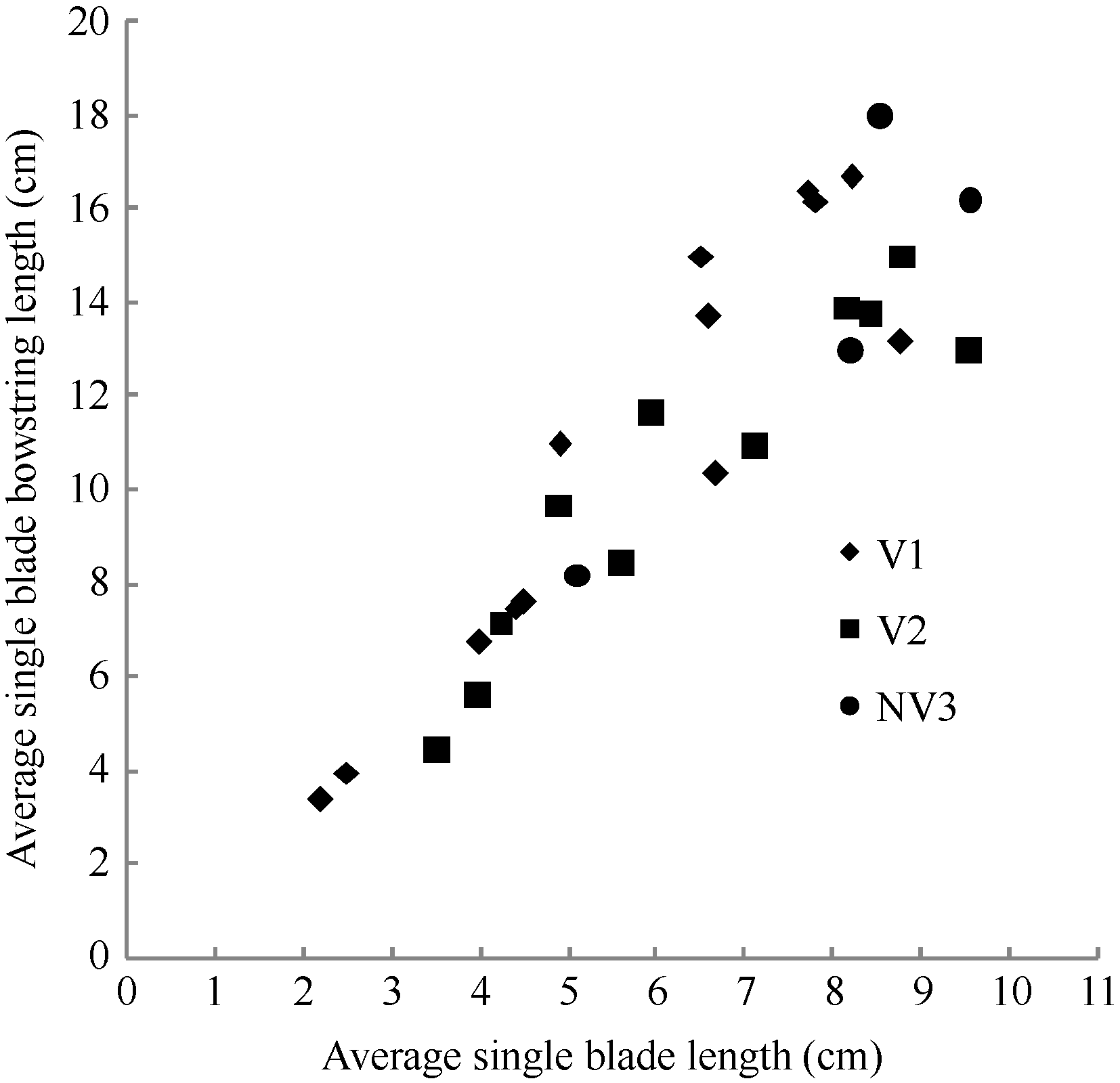 | 图5 2011-2012年不同处理叶弦长随叶长变化规律Fig. 5 Changes in LBBL values of different treatments with leaf blade length for three cultivars in 2011-2012 |
2.1.4 叶柄长度模型 油菜叶片可分为长柄叶、短柄叶和无柄叶3种类型, 本文主要涉及前二者。2011— 2012年不同处理油菜长柄叶柄长和短柄叶柄长随叶长变化均呈幂函数增长(图6), 其相关系数分别为r=0.928, P< 0.001, n=18, r(16, 0.001)=0.708, R2=0.862; r=0.887, P< 0.01, n=10, r(8, 0.01)=0.765, R2=0.786, 模型参数及显著性测验见表1, 方程的F值、t值和a3、b3、a4均达到P< 0.001显著水平, b4达到P< 0. 01显著水平。因此, 不同品种不同处理长柄长LPLj(y) (cm)和短柄长SPLj(y) (cm)随叶长变化模型分别表述如下:
LPLj(y) = a3LLj(y)b3, 1 ≤ j≤ 4 (9)
SPLj(y) = a4LLj(y)b4, 5 ≤ j≤ 8 (10)
式(9)和(10)中, LLj(y) (cm)是油菜出苗第y天主茎第j叶的叶长, a3、b3、a4、b4为参数。
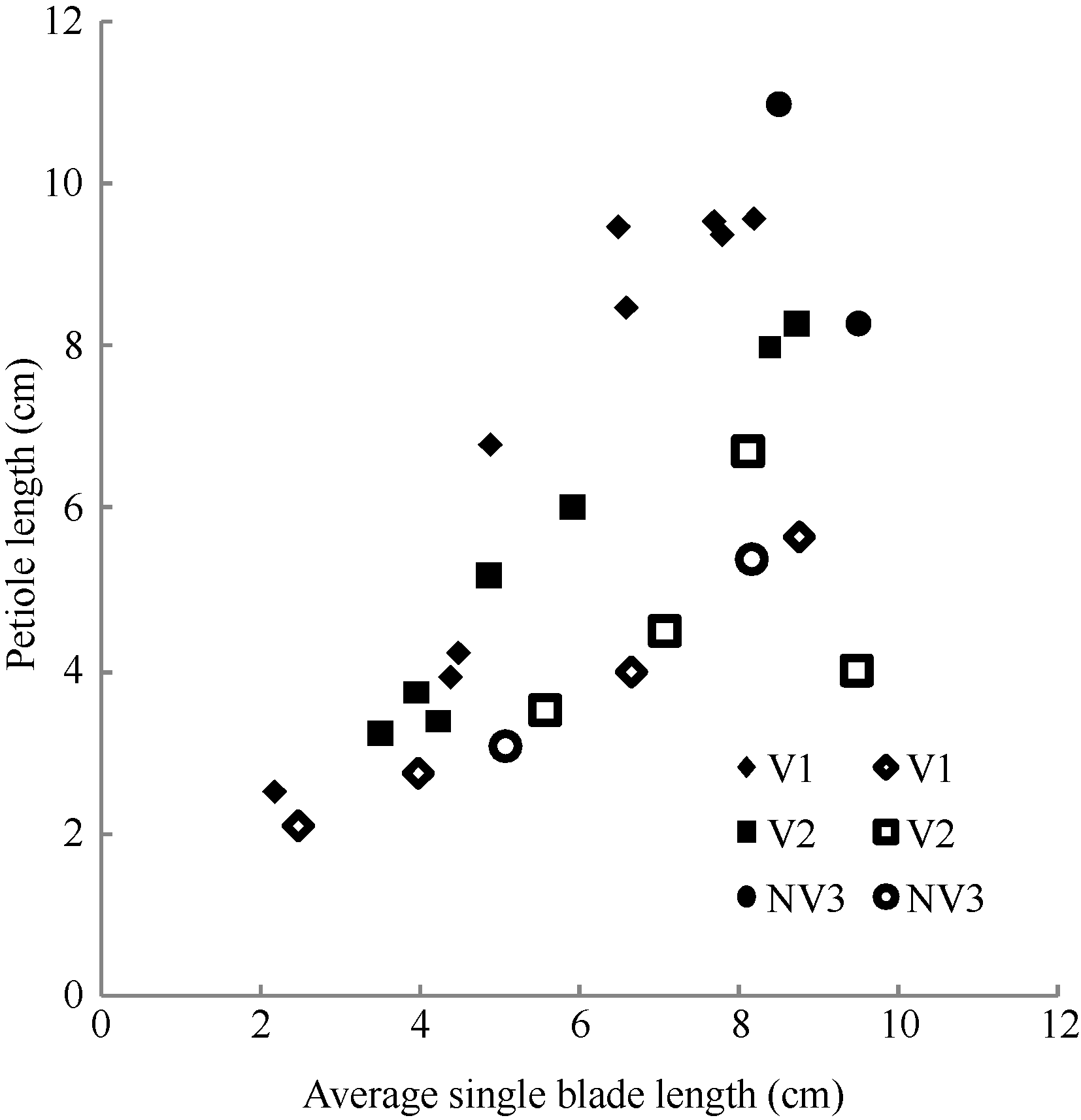 | 图6 2011-2012年不同处理叶柄长随叶长变化规律Fig. 6 Changes in PL values of different treatments with leaf blade length for three cultivars in 2011-2012 |
2.1.5 叶片角度模型 为了准确测量, 定义了2个油菜叶片与主茎的夹角(图7), 分别为叶切角和叶弦角。叶切角和叶弦角的平均值可用于表示茎叶夹角。而理论上叶片角度与叶片干物重关系密切。因此, 叶切角(TAj)( ° )和叶弦角(BAj) ( ° )可表示为:
TAj(y)=DWLBj(y) × WRTWj(y) (11)
BAj(y)=DWLBj(y) × WRBW j(y) (12)
ASLj(y)= [TAj(y) + BA j(y)]/2 (13)
AOj(y)=180 - ASLj(y) (14)
式(11)至(14)中, TAj是主茎与叶柄的夹角(° ), BAj是主茎与叶弦的夹角(° ), RTWj(y)是比叶切角(° g-1), RBWj(y)是比叶弦角(° g-1), ASLj(y)是茎叶夹角(° ), AOj(y)是叶倾角(° )。
 | 图7 叶片角度示意图Fig. 7 Schematic diagram of the leaf blade angle |
2011— 2012年试验表明, 比叶切角与叶位的关系近似于S型曲线(图8)。其相关系数r=0.661, P< 0.01, n=28, r(26, 0.01)=0.479, R2=0.438, 模型参数及显著性测验见表1, 方程的F值、t值、a5和b5均达到P< 0.001显著水平; 比叶弦角与叶位的关系近似于幂函数(图8)。其相关系数r=0.599, P< 0.01, n=28, r(26, 0.01)=0.479, R2=0.359, 模型参数及显著性测验见表1, 方程的F值、t值和a6均达到P< 0.001显著水平, b6达到P< 0.01显著水平。因此, 不同品种与施肥处理比叶切角RTWj(y)(° g-1)和比叶弦角RBWj(y) (° g-1)随叶位变化模型分别表述如下:
RTWj(y)=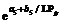
RBWj(y)=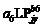
式(15)和(16)中, LPjy是油菜出苗第y天主茎第j片叶的叶位, a5、b5、a6、b6均为参数。
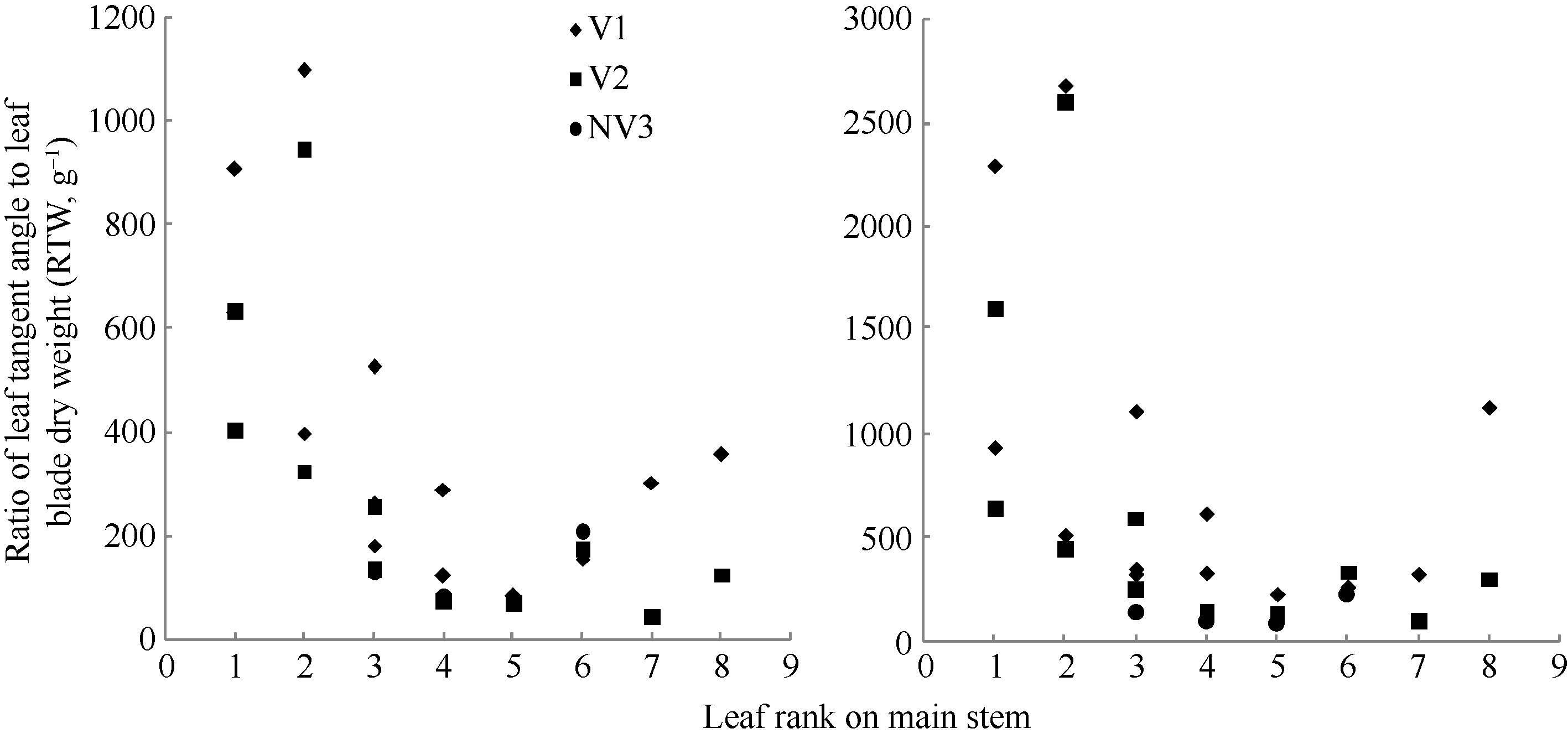 | 图8 2011-2012年不同处理比叶切角和比叶弦角随叶位变化规律Fig. 8 Changes in RTW, RBW values of different treatments with leaf rank on main stem for three cultivars in 2011-2012 |
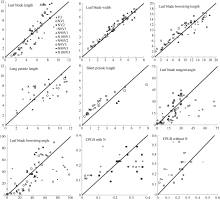
2011— 2013年油菜越冬前植株地上部形态参数叶长、叶宽、叶弦长、柄长(长柄和短柄)以及叶切角和叶弦角观察值与模拟值的da值和RMSE值分别为-0.231 cm, 2.102 cm (n=63); -0.273 cm, 0.484 cm (n=63); -0.343 cm, 1.963 cm (n=63); 0.412 cm, 2.095 cm (n=36); -0.635 cm, 1.006 cm (n=27); 4.421° , 14.734° (n=63); 6.6424° , 21.817° (n=63)。以上形态参数观察值与模拟值的相关系数(r)均达到P< 0.001显著水平, 但其dap值小于5%的仅有叶长和弦长, 其模型精度最好, dap值在5%~7%之间的为叶宽和长柄长, 其模型精度较好, dap值大于10%的为短柄长、叶切角和叶弦角, 其模型精度较差(表3)。与施肥品种相比, 不施肥品种CPLB观察值与模拟值的相关系数(r)达到P< 0.001显著水平, 但其dap值大于10%, 说明所建的不施肥品种CPLB模型精度较差。施肥品种观察值与模拟值的统计参数da和RMSE的值都较小, 且dap值小于10%, 其模型精度较高, 说明观察值与模拟值的拟合度较好(表3)。以上观察值与模拟值的1∶ 1关系图见图9。
| 表3 2011-2013年油菜越冬前植株地上部形态结构模型观察值与模拟值比较的统计参数 Table 3 Comparison of statistical parameters of simulation and observation in rapeseed aboveground architectural parameter models at pre-overwintering stage in 2011-2013 |
从表3和图9可见, 短柄长、叶切角和叶弦角、不施肥品种CPLB值误差较大, 说明该模型在今后工作中仍需进一步改进。总体上, 2011— 2013年油菜越冬前试验资料检验效果较好。
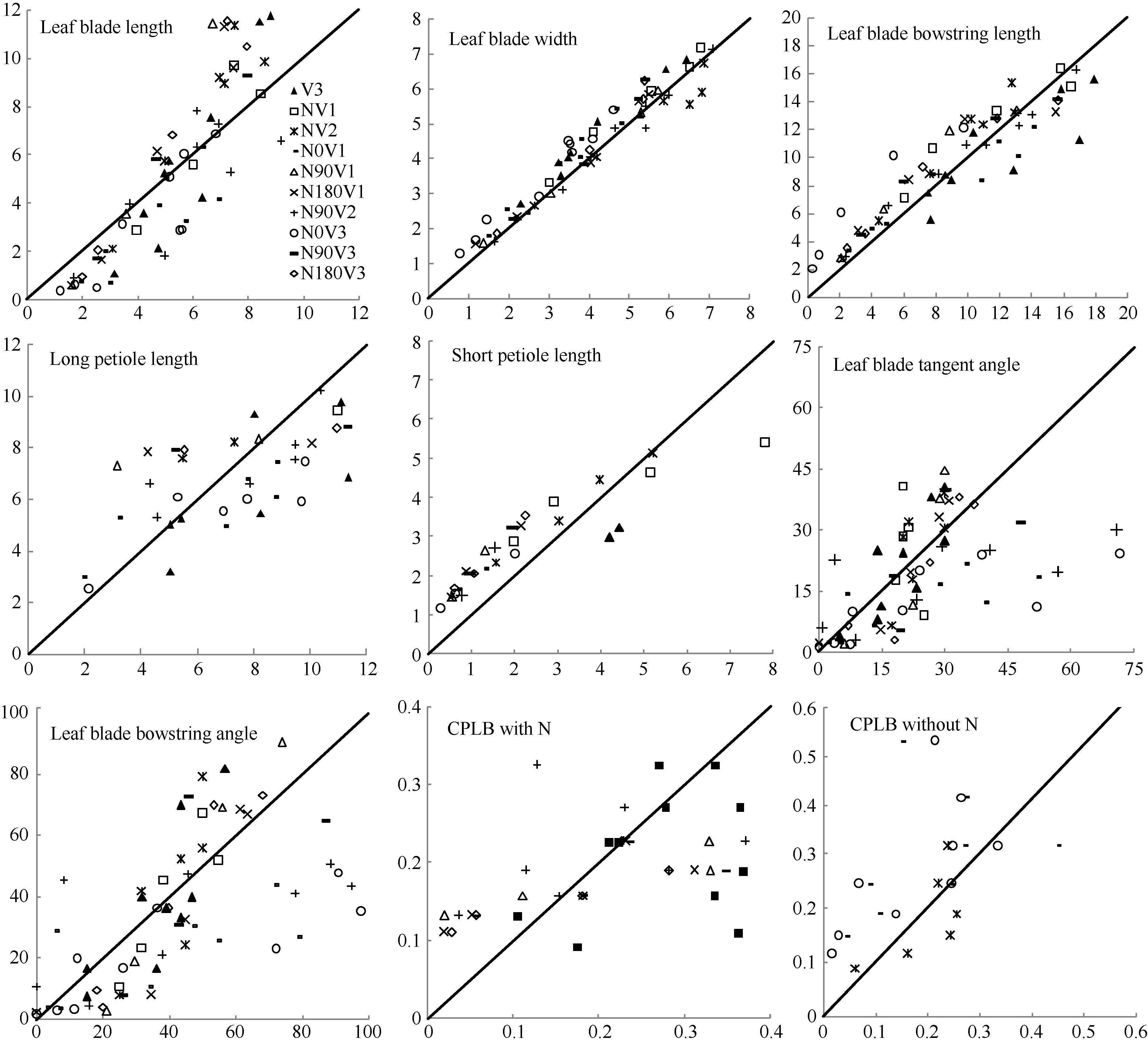 | 图9 2011-2013年观察值与模拟值比较Fig. 9 Comparison of the observed with the simulated in 2011-2013 |
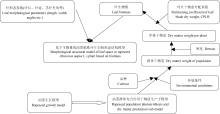
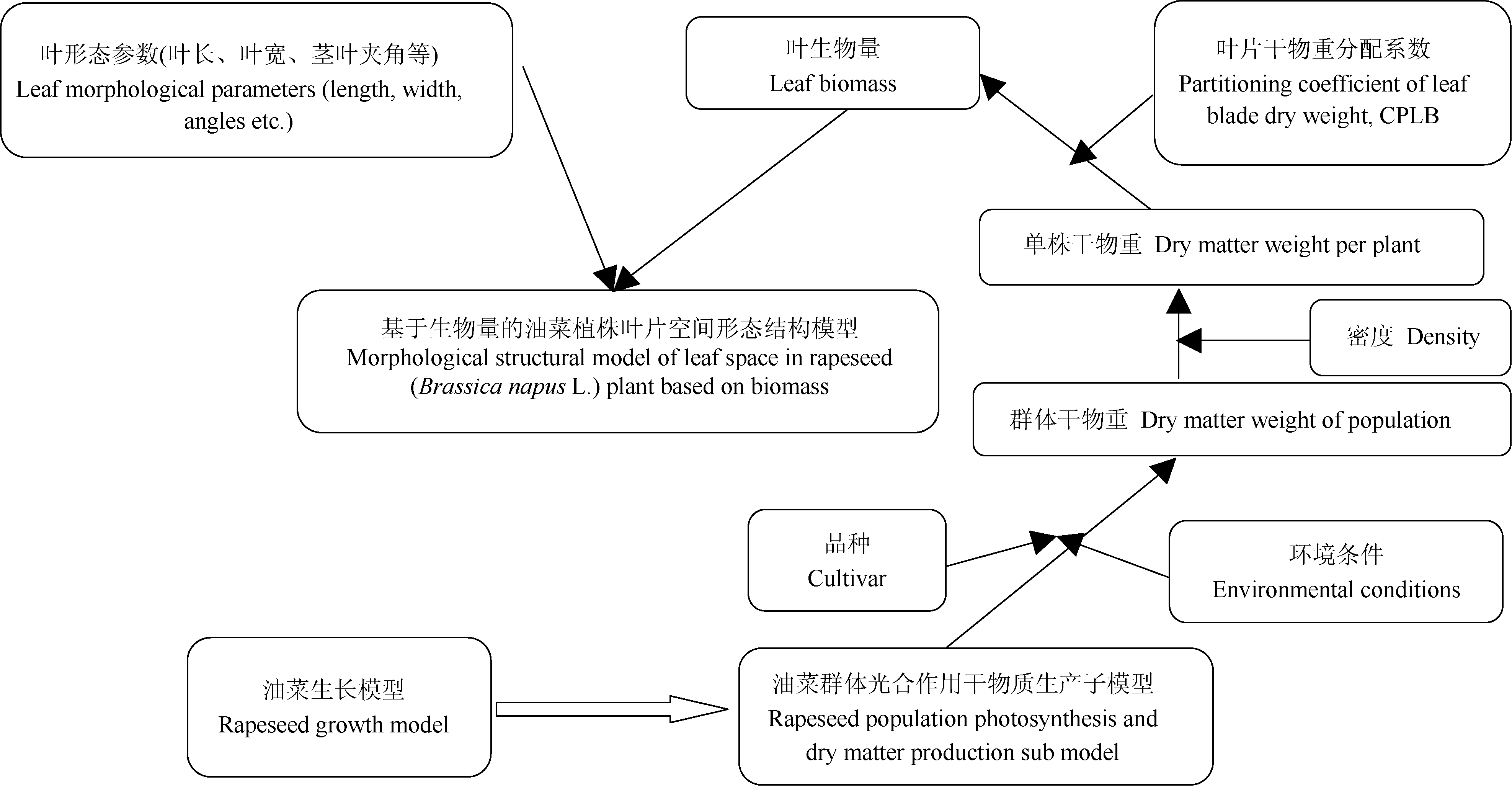
油菜植株生物量是光合产物分配与积累的结果[16], 而光合产物的形成受油菜品种和环境因素(如光、温度、水分、CO2和施肥等)的综合影响, 叶片是光合作用的重要器官。刘岩等[28]根据生物量与叶片几何属性的关系, 建立了基于生物量的水稻主茎叶片几何属性模型。赵泽英等[29]分析了温度对辣椒叶片形态发生的影响, 量化了温度与叶片形态建成的关系, 构建了基于有效积温(GDD)、以Logistic方程为基础的辣椒叶片长度、宽度及数量形态发生的动态模型。本文通过建立油菜叶片生物量与叶片形态参数的关系模型, 可实现油菜生长模型与油菜形态结构模型的结合(图10), 由于油菜生长模型中的群体光合作用与干物质生产子模型可在不同品种和环境条件下模拟油菜群体干物重, 进而与密度结合生成单株干物重, 再通过干物重分配系数生成叶生物量并形成叶片形态参数。可见, 本文的叶片形态参数模型能通过生物量响应品种和环境条件差异。
我们之前已建立了具有机理性和普适性的油菜生长模型[16], 并已验证, 油菜生长模型可以输出不同品种、环境条件下的干物重, 而我们现在建立的形态模型, 找到了干物重和形态之间的关系, 只要有干物重结果, 就可以模拟不同品种和环境下油菜植株的形态。因此, 本文建立生物量和油菜叶片形态参数间联系, 可通过生物量将油菜生长模型与油菜形态结构模型有机结合, 为油菜功能-结构模型建立奠定基础。本文选择叶长与生物量建模, 然后将最大叶宽、叶弦长、叶柄长与叶长建模, 其中, 最大叶宽模型和叶弦长模型都可由线性函数表示, 说明叶宽和叶弦长随叶长的增加而增加, 叶柄长模型随叶长变化均呈幂函数增加。由于油菜叶片形状不规则, 本研究测量的是油菜最大叶长和叶宽, 利用量角器测量油菜茎叶夹角和叶弦角容易产生测量误差, 为避免误差过大, 在测量叶弦角时借助直尺确定叶弦, 然后用量角器量取叶弦角度。为了能较准确测量“ 茎叶夹角” , 本文采用了“ 叶切角和叶弦角” , 而利用“ 叶切角和叶弦角” 的平均值表示“ 茎叶夹角” 和“ 叶倾角” , 以便和常用参数相关联。岳延滨[23]进行了基于生长度日(GDD)的油菜形态结构模型研究, 尚未涉及与生长模型的结合。Groer等[18]、Mü ller等[19]在油菜功能-结构植物模型研究中也采用了将油菜生长模型与油菜形态结构模型相结合方法。目前, 有学者利用三维扫描仪扫描油菜叶片, 建立油菜单株模型, 该模型能够精确描述叶片形态, 并能精确计算叶片面积, 但仅是对油菜结构模型的分析研究, 并不能响应品种与环境因子变化。刘宏伟等[30]从物理学角度, 提出基于实测数据构建的水稻叶片数学模型。该模型利用VC++与OpenGL相结合的方法, 通过具有较明确生物学意义的形态参数, 实现水稻叶片形态和形变过程的矢量化和虚拟生长。油菜叶片颜色也属于油菜形态结构的重要因素之一, 朱艳等[31]以冬小麦为研究对象, 模拟小麦叶色变化的动态过程, 为实现小麦生长的可视化表达奠定基础。本文由于试验条件的限制, 未分析研究油菜叶色。
建立了基于生物量的油菜越冬前植株叶片空间形态结构模型。经检验, 除短柄长、叶切角和叶弦角、不施肥品种CPLB值误差较大以外, 该模型观察值与模拟值一致性较好。
油菜比叶长重(RLW)、比叶切角(RTW)和比叶弦角(RBW), 可将生物量与叶长、叶切角和叶弦角相联系, 是具有生物学意义的油菜形态结构模型参数。油菜叶片的CPLB值、油菜叶切角和叶弦角模型均有明显误差, 造成这些误差的主要因素是油菜叶形呈不规则羽状缺裂及不易测量等复杂性。本研究建立的模型是可行的。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|