

{kind=link}
{kind=link}
{kind=link}
棉花溶血磷酸酯酰转移酶(LPAT)家族基因的发掘和表达分析
[丁检, 吴双, 蔡彩平, 郭旺珍*  ]
]
]
|
|


棉花油脂合成相关代谢在控制油分形成、纤维发育等进程中起着重要作用。溶血磷脂酰基转移酶(LPAT)是植物油脂代谢过程中的一个关键酶。本研究利用雷蒙德氏棉全基因组数据库, 通过生物信息学技术, 鉴定并获得8个棉花LPAT家族基因的全序列和染色体定位等信息。通过序列比对进行系统进化和分类分析, 明确这8个家族基因分布在6条染色体上, 分为4亚类, 其中第I类和第III类中各含2个基因、第II类1个、第IV类3个。组织表达分析表明这8个基因在棉花营养和生殖阶段均表现表达多样性。 LPAT6和 LPAT8在17 d胚珠中表达水平很高, 推测在油脂合成代谢调控中起重要作用。8个LPAT基因均在纤维中优势表达, 其中 LPAT2、 LPAT3和 LPAT4表达水平更高, 推测LPAT家族基因的表达参与棉纤维发育进程。
Metabolism related to lipids synthesis plays an important role in regulating both oil biosynthesis and fiber development in cotton. Lysophosphatidic acid acyltransferase (LPAT) is a key enzyme in oil biosynthesis pathway in plant. In this study, eight cotton LPAT family genes were identified and their gene sequences, chromosome location were obtained, based on G. raimondii genome database (http://www.phytozome.net/) and bioinformatic method. These LPAT members were anchored on six chromosomes in G. raimondii. Phylogenetic analysis showed that LPAT candidate genes were classified into four groups, with two members each in group I and group III, one in group II, and three in group IV. The expression patterns of LPAT genes revealed their important roles in diverse functions in the developmental stages of vegetative and reproductive growth in cotton. LPAT6and LPAT8 showed the highest expression level in ovules at 17 days post-anthesis, which might play an important role in regulating oil biosynthesis. Eight genes showed the preferential expression level in fiber development stages. Among them, LPAT2, LPAT3, and LPAT4showed the higher expression level in fiber than in other tissues and organs, implying their association with cotton fiber development.
植物油脂的主要成分是甘油三脂肪酸酯, 又称三酰甘油(TAG)。其生物合成主要在内质网上完成, 由甘油-3-磷酸酰基转移酶(GPAT)、溶血磷脂酸酰基转移酶(LPAT, 又名AGPAT)和二酰甘油酰基转移酶(DGAT)调控, 这3种酰基转移酶依次将酰基CoA的脂肪酸转移到甘油, 该途径称为依赖酰基CoA的Kennedy途径[1]。LPAT是甘油三酯合成途径中的第2个酰基转移酶, 其功能是将脂肪酰基CoA结合到甘油三酯sn-2位上形成磷脂酸(PA), 产生的磷脂酸既可以继续脱磷酸进而合成TAG, 也可以进入磷脂合成途径, 参与生物膜的形成[2]。因此, LPAT具有极其重要的生理功能。目前, LPAT的同源基因已在拟南芥(Arabidopsis thaliana)[3]、蓖麻(Ricinus communis)[4]和花生(Arachis hypogaea)[5]等植物中被分离克隆。研究表明拟南芥中至少存在9个LPAT基因[6], 虽然其基本功能相似, 但作用底物、表达特性以及亚细胞定位等存在极大差异[3, 7, 8]。植物中, 不同LPAT对不同酰基CoA具有特异的选择作用, 它在sn-2位上优先转移的底物是不饱和酰基链。由于TAG的不饱和脂肪酸通常位于sn-2位, 因此调控LPAT基因表达对利用基因工程改良种子脂肪酸成分, 尤其是降低饱和脂肪酸含量具有重要的研究价值。在拟南芥和油菜中过量表达酵母LPAT基因突变型(SLC1-1), 可增加TAG sn-2位上长链脂肪酸比例和含量, 使种子含油量提高8%~48%[9]。在拟南芥种子中特异过表达编码油菜微粒体的2种LPAT基因, 转基因后代种子重量和脂肪酸含量分别比对照平均增加6%和13%[10]。
棉花是重要的纤维作物, 也是世界第六大油料作物。棉纤维发育是一个非常复杂的过程, 涉及大量代谢途径, 上千个特异基因参与表达调控。油脂类物质具有重要的生理功能, 可通过不同代谢途径形成结构各异、功能相关的一组化合物, 调节相关蛋白表达, 从而影响纤维的生长发育。冷雪等[11]通过基因芯片方法对棉花纤维伸长相关基因研究发现, 参与油脂代谢的多个基因在棉纤维伸长阶段上调表达。Qin等[12]研究表明长链脂肪酸与纤维的伸长高度相关。另一方面, 棉籽作为棉花的副产物, 其种子含油量约23%~26%, 是重要的植物油料资源。棉籽油是生产食用油、工业原料[13]以及生物柴油的理想原料[14]。因此, 明确棉花油脂代谢过程中一些关键基因及其表达特征, 利用油脂代谢基因工程技术定向改善棉花油分和纤维品质, 对阐明棉纤维发育机制, 提高棉花综合利用效率均有重要意义。本研究基于棉属二倍体野生种雷蒙德氏棉基因组序列信息[15], 利用生物信息学手段, 进行全基因组棉花LPAT家族基因的鉴定, 染色体定位和分类研究, 并明确其同源基因在栽培陆地棉不同组织器官的表达特征, 为深入阐明该类基因功能特征及育种利用奠定基础。
选用陆地棉遗传材料7235, 于2012— 2013年种植于南京农业大学网室。在盛花期, 采集开花后0和5 DPA (days post-anthesis)的胚珠和纤维混合物, 10、14、17、20和23 DPA的纤维及胚珠组织, 迅速置液氮中, 在-70℃冰箱保存。根、茎和叶组织取自有5~6片真叶的棉花幼苗。
棉花二倍体野生种雷蒙德氏棉全基因组数据下载于Phytozome网站(http://www.phytozome.net/)。使用Pfam数据库工具发掘LPAT家族基因, 获得其保守结构域Acyltransferase (PF01553)的种子文件[16], 再通过HMMER 3.0软件[17]检索已有的数据库。从GenBank EST数据库(http://www.ncbi.nlm.nih.gov/dbEST/)下载陆地棉(G. hirsutum)、海岛棉(G. barbadense)、亚洲棉(G. arboreum)和二倍体野生种雷蒙德氏棉(G. raimondii)的EST序列。采用GSDS程序[18]完成基因结构分析。利用DNAstar (http://www.dnastar.com/)完成ORF、等电点和分子量预测。根据数据库中棉花LPAT基因序列信息, 获得每个基因在雷蒙德氏棉染色体上的分布情况。
利用ClustalX软件[19]进行多重序列比对。使用MEGA软件(Ver 5.05)[20]通过多重序列比对, 利用Maximum-likelihood法(BootStrap检验1000次)构建系统发育树。拟南芥和其他植物中的LPAT基因和蛋白序列分别下载于TAIR (http://arabidopsis.org/)和NCBI (http://www.ncbi.nlm.nih.gov/)。
采用CTAB-酸酚法[21]分别提取10、14、17、20和23 DPA纤维和胚珠的总RNA。采用RNA试剂盒(Bioer Technology, 北京)提取0、5 DPA胚珠和纤维混合物, 及根、茎和叶的总RNA。采用微量分光光度计(Nanodrop, USA)和1%琼脂糖凝胶检测RNA的浓度和质量。取2 μ g总RNA用于cDNA第一链的合成, 研究方法参照试剂盒说明书(Vazyme Biotech, 南京)。
参照棉花不同LPAT基因的转录组序列, 利用Primer5.0 (http://www.premierbiosoft.com/)设计各基因特异的qRT-PCR引物, 扩增片段为75~200 bp, 退火温度设定在58~60℃ (表1)。引物合成由南京金斯瑞生物科技有限公司完成。20 μ L反应体系包含SYBR Green Realtime PCR Master mix 10 μ L, LPAT-F/R引物各1 μ L, cDNA模板1 μ L, 去离子水8 μ L。扩增条件为95℃ 3 min; 95℃ 15 s, 60℃ 20 s, 72℃ 20 s, 40个循环; 72℃ 10 min。以棉花持家基因His3(AF024716)作为内参基因[22]。利用ABI实时荧光定量PCR仪(7500型)检测目标基因的表达情况。每个样品重复3次。采用2-Δ CT法进行数据的相对定量分析[23]。
| 表1 LPAT家族基因表达分析所用引物 Table 1 Primers for expression analysis of LPAT family genes |
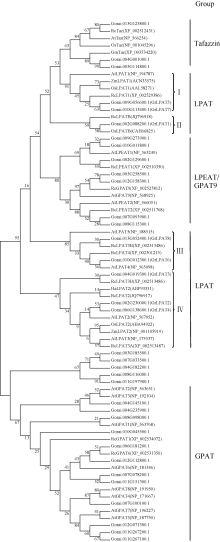
以保守结构域Acyltransferase (PF01553)为种子文件, 通过生物信息学手段搜寻雷蒙德氏棉全基因组数据库, 获得35个含该结构域的候选基因。为了进一步发掘棉花LPAT候选基因, 提取拟南芥、蓖麻、玉米、水稻等作物中已经公布的LPAT、GPAT、LPEAT和Tafazzin等类氨基酸序列, 与棉花中35个含酰基转移酶保守结构域基因进行系统发育分析。参照蓖麻的酰基转移酶相关基因的分类方式[4], 棉花35个酰基转移酶相关基因分属于4个(LPAT、GPAT、LPEAT和Tafazzin)大类, 其中8个基因与其他物种中的LPAT基因亲缘关系较近(图1)。通过比对棉花EST数据库, 8个基因均在陆地棉(G. hirsutum)、海岛棉(G. barbadense)、亚洲棉(G. arboreum)和雷蒙德氏棉(G. raimondii)棉种中发现至少一个与其同源性高的EST序列(E≤ -10), 说明这8个基因属于棉花LPAT家族基因, 且均有转录活性。按照雷蒙德氏棉染色体与四倍体栽培棉种同源染色体整合结果, 明确棉花8个LPAT家族基因分布在6条染色体上。参考四倍体栽培棉种染色体编号, 依照染色体从小到大位置顺序将LPAT基因家族8个成员命名为GrLPAT1~8(表2)。
| 表2 棉花LPAT家族基因的基本信息 Table 2 Basic information of LPAT family genes in cotton |
由系统进化树分析表明, LPAT家族基因主要分为4类, 第I类(LPAT1)包括拟南芥、蓖麻、玉米和水稻的质体型LPAT1基因, 以及2个棉花的LPAT基因; 第II类(LPATB)仅包括蓖麻LPATB, 水稻LPATB和一个棉花LPAT基因; 第III类(LPAT4/5)包括拟南芥的LPAT4和LPAT5, 蓖麻的LPAT4和LPAT5, 以及2个棉花的LPAT基因; 第IV类(LPAT2/3)包括多个物种中已经报道的LPAT2和LPAT3基因, 而棉花中共有3个LPAT基因分布在这一类中。从分类结果可知, LPAT家族基因存在类型的多样性, 这可能与基因功能的分化有关。
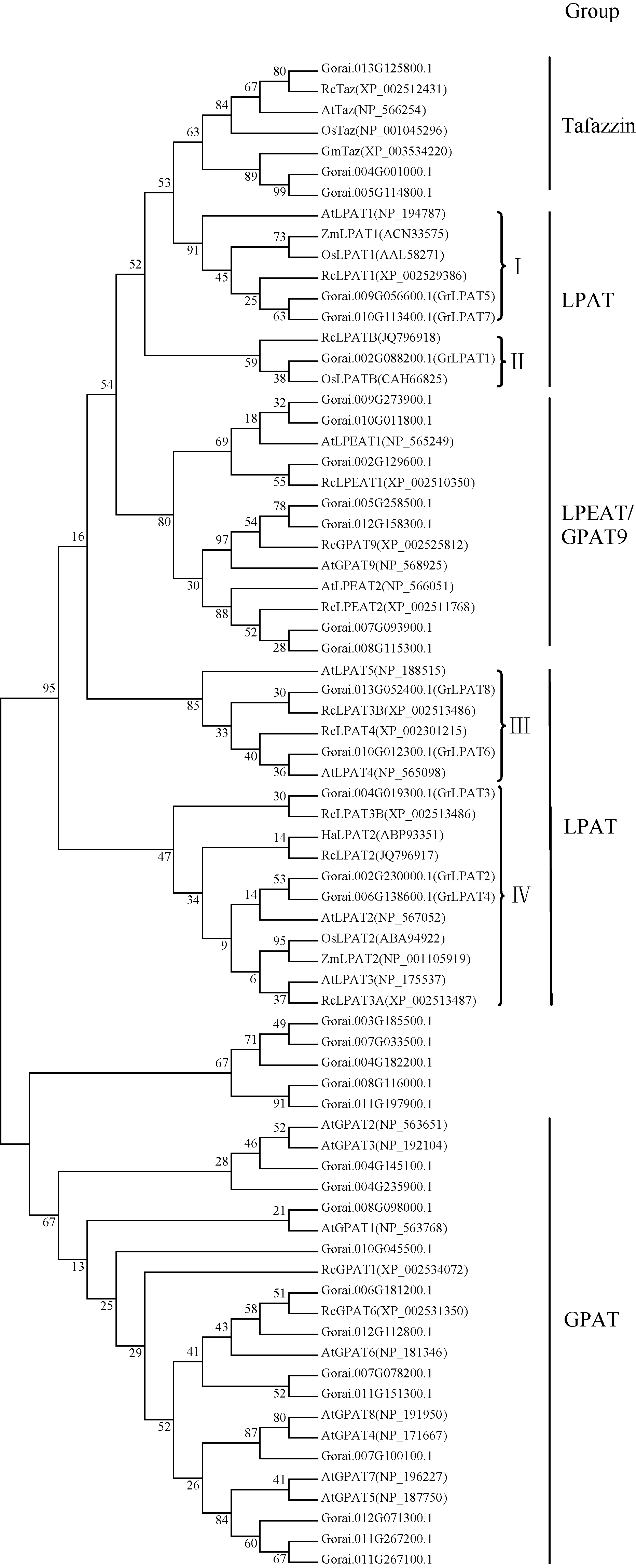 | 图1 不同物种来源LPAT基因家族的系统进化树 At: 拟南芥; Ah: 花生; Os: 水稻; Gm: 大豆; Zm: 玉米; Rc: 蓖麻; Ha: 向日葵; Gorai: 雷蒙德氏棉. I: LPAT1; II: LPATB; III: LPAT4/5; IV: LPAT2/3. 进化树分支上的数字表示BootStrap (1000次)验证中该分支可信度的百分比。Fig. 1 Phylogenetic tree of LPAT faimly genes from different species At: Arabidopsis thaliana; Ah: Arachis hypogaea; Os: Oryza sativa; Gm: Glycine max; Zm: Zea mays; Rc: Ricinus communis; Ha: Helianthus annuus; Gorai: Gossypium raimondii. I: LPAT1; II: LPATB; III: LPAT4/5; IV: LPAT2/3. BootStrap values (%) based on 1000 replicates are indicated beside the nodes. |
棉花8个LPAT家族基因编码的氨基酸大小在204~387之间, 其等电点范围从8.59~9.81 (表2)。根据NCBI和Pfam保守结构域预测显示, 棉花LPAT蛋白除含有一个酰基转移酶acyltransferase结构域(PF01553), 还包含一个与溶血磷脂酰基转移酶相似的保守区域(LPLAT)[24]。8个基因编码的LPAT蛋白均含有酰基转移酶的功能区段, 与棉花油脂代谢相关。
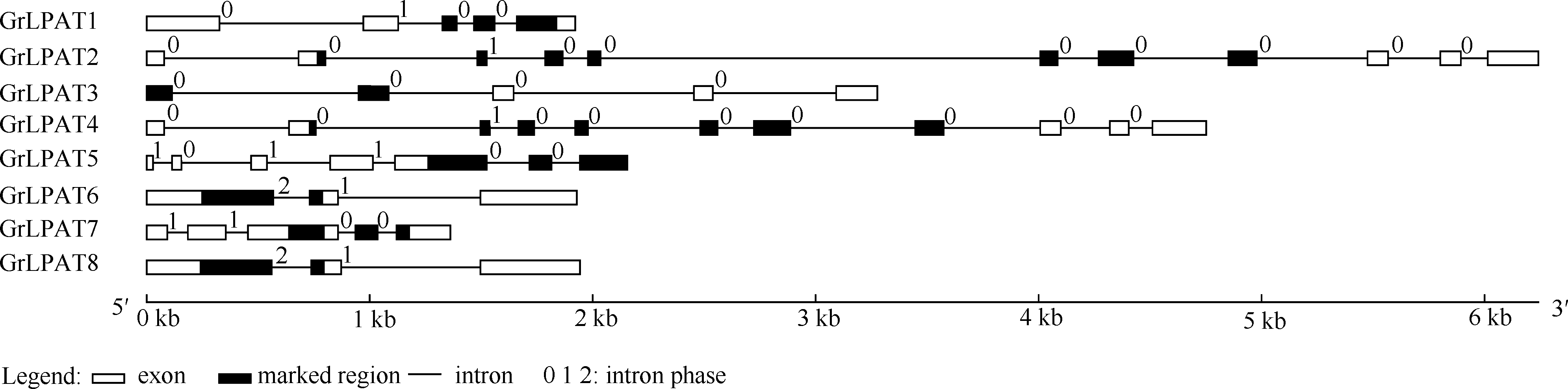 | 图3 棉花LPAT家族基因结构分析 方框代表外显子, 黑色方框代表保守结构域, 黑色细线代表内含子, 0, 1, 2代表内含子插入位置。Fig. 3 Structural characterization of eight LPAT family genes in Gossypium raimondii The boxes, black boxes and black lines represent exon, domain and intron location, respectively. Number 0, 1, 2 represent the inserted location of introns. |
基因结构分析(图3)显示, 所有棉花的LPAT基因都存在2个以上的内含子, 最多含10个内含子。内含子长度介于78~1969 bp之间; 外显子数目为3~ 11个, 长度介于34~569 bp之间。这种内含子-外显子结构严重异化现象可能导致其基因的功能分化。
进化分析表明, 雷蒙德氏棉是异源四倍体D亚基因组亲缘关系最近供体[26]。陆地棉是全球栽培面积最大的异源四倍体棉花, 含A和D亚基因组, 存在雷蒙德氏棉的同源基因。基于定量RT-PCR技术, 进一步分析这8个LPAT家族基因其同源基因在四倍体陆地棉不同组织、器官中表达特征。以陆地棉7235的根、茎、叶、胚珠和纤维混合物(0和5 DPA)、不同发育时期纤维和胚珠(10、14、17、20和23 DPA)等多组织器官cDNA为模板, 以持家基因His3为内参, 定量分析表明, 8个LPAT家族基因在陆地棉中的表达特征呈现多样化。(1) LPAT5和LPAT7属于LPAT1质体型基因, 在供检测的陆地棉不同组织器官中表达水平均较低, 但在17和20 DPA的纤维组织中表达水平相对较高; (2) LPAT1属于LPATB型基因, 在供检测的陆地棉不同组织器官中表达水平也较低, 但在叶中的表达量最高, 在不同发育时期的纤维组织中表达水平相对较高; (3) LPAT6和LPAT8属于LPAT4/5型基因, 在17 d棉花胚珠中表达量很高, 在不同发育期的纤维组织中也优势表达; (4) LPAT2、LPAT3和LPAT4属于LPAT2/3型基因, 该类基因在纤维中表达量高。其中LPAT2在14 DPA的纤维中表达量达到最高, 在10、17和20 DPA的纤维组织中其表达量也很高; LPAT3在纤维中的表达水平随着纤维发育进程逐渐增高, 在23 DPA的纤维组织中表达量最高; LPAT4在所有供试组织器官都表达, 在根、茎、叶, 早期的胚珠中表达水平较高, 在10 DPA的纤维组织中表达量最高。研究发现, 属于同一类的不同基因有不同的表达特征, 表明进化进程中, 家族内同类基因其结构的相似性和功能分化的多样性。
植物体内甘油脂类代谢涉及很多酶类, 如溶血磷脂酸酰基转移酶(LPAT)、甘油-3-磷酸酰基转移酶(GPAT)、溶血磷脂酰乙醇胺酰基转移酶(LPEAT)和心磷脂酰基转移酶(Tafazzin)等, 它们都有一个共同的酰基转移酶保守结构域Acyltransferase (PF01553)[27]。本研究利用生物软件和Pfam数据库进行棉花LPAT家族基因发掘, 初步得到35个包含有多个亚家族成员的候选基因。基于拟南芥和其他作物中已经公布的LPAT、GPAT、LPEAT和Tafazzin等类基因的氨基酸序列, 通过聚类分析, 将二倍体雷蒙德氏棉基因组中的35个候选基因明确分为GPAT、LPAT、LPEAT/GPAT9以及Tafazzin 4大类(图1), 并从中发掘出8个棉花LPAT家族基因成员。前人报道, LPATs呈现底物特异性与类型多样性的双重特点[28]。进一步利用陆地棉TM-1全基因组序列信息(内部交流资料)发掘到16个LPAT家族基因, 均表现为二倍体雷蒙德氏棉基因组中的1个基因对应四倍体陆地棉TM-1基因组中2个基因。聚类结果表明其分类与本文研究结果一致, 没有出现其他类型。因此, 是否在不同棉种中存在其他类型的LPAT家族成员, 需进一步分析。
目前已从拟南芥、蓖麻等植物中完成LPAT家族基因鉴定, 其中拟南芥至少存在9个家族成员[6], 蓖麻中存在7个家族成员[4]。这些家族基因在亚细胞定位、表达特性和功能上呈现多样性, 如拟南芥LPAT1定位于质体, 与胚胎发育密切相关[8]; LPAT2定位于内质网, 影响雌配子的发育; LPAT3定位未知, 在花粉中优势表达[3]; 在体外检测不到LPAT4和LPAT5的溶血磷脂酸酰基转移酶活性, 它们可能与
心磷脂合成有关[7]。聚类分析表明, 拟南芥、蓖麻等作物中存在的不同类型LPAT, 在棉花中均被鉴定。从不同组织器官的表达特征看, 8个棉花LPAT基因呈现表达多样性, 即使是同一类型的LPAT, 表达特征也各不一样, 反映出功能分化的多样性, 其具体的功能特征有待进一步研究。
溶血磷脂酰基转移酶(LPAT)是植物油脂代谢过程中的一个关键酶。棉花是重要的纤维和油料作物。实时荧光定量PCR分析表明, 8个棉花LPAT家族基因在不同发育时期的胚珠和纤维中呈现表达水平多样性。戚维聪等[29]研究油菜种子发育过程中Kennedy途径的各种酶活性变化, 发现LPAT活性最高, 且高含油量品系中LPAT活性高于低含油量品系。陈四龙等[5]研究发现花生LPAT4基因的表达量与种子含油量积累速率一致。棉花LPAT6和LPAT8在发育17 d的胚珠中优势表达, 推测其可能参与种子的油脂合成代谢。棉纤维发育是一个非常复杂的过程, 研究发现, 在棉纤维伸长阶段检测到多个显著上、下调表达基因, 它们主要参与脂肪酸代谢、碳固定、脂肪和淀粉代谢等途径, 通过调控脂肪酸代谢来影响纤维的发育[11]。Qin等[12]研究表明, 油脂合成相关代谢中的3-酮酰-CoA合成酶(KCS)在棉纤维伸长过程中起重要的作用。LPAT作为脂肪酸代谢途径上游的关键基因, 既参与甘油酯合成, 又参与生物膜形成, 其活性高低直接影响脂肪酸和膜脂类物质的生物合成, 进而影响纤维的发育。Wanjie等[30]发现在棉纤维发育进程中, 多条与LPAT基因高度同源的EST序列优势表达, 表明LPAT基因参与的代谢进程与棉纤维的伸长发育密切相关。本研究发现, 棉花8个LPAT基因均在纤维发育进程中优势表达, 其中LPAT2、LPAT3和LPAT4基因相对表达量最高。我们推测LPAT家族基因可能与纤维细胞的快速伸长需要脂类物质提供相应的能量和生物膜相关, 其作用机理有待进一步研究。综上所述, 棉花LPAT家族基因不仅在控制种子油分合成中有重要作用, 在棉纤维生长发育中同样扮演重要角色。
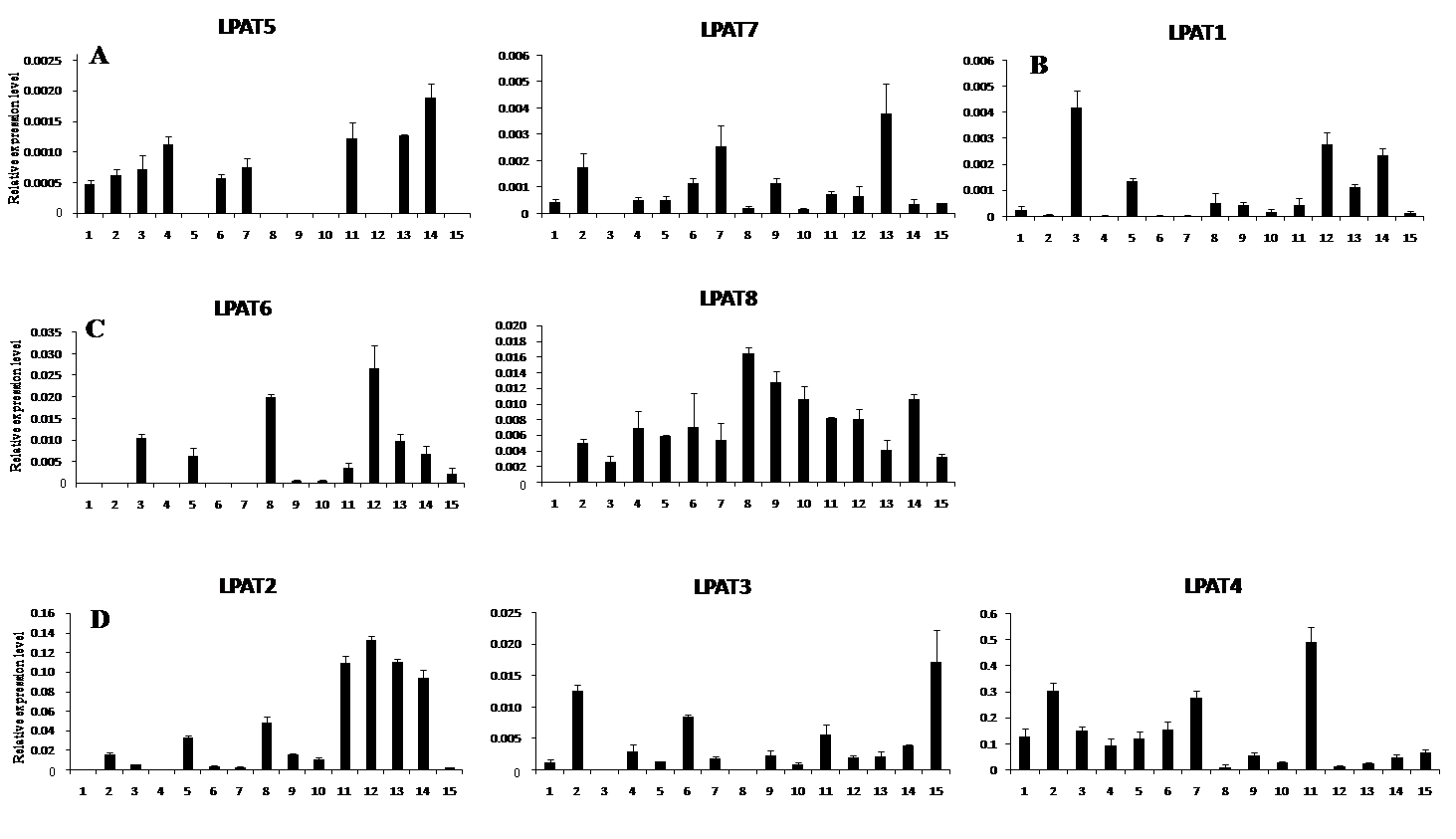 | 图4 LPAT家族基因在陆地棉7235不同组织和器官中的表达特征 1: 根; 2: 茎; 3: 叶; 4: 开花当天胚珠; 5: 开花后5 d胚珠和纤维混合物; 6~10: 分别代表开花后10、14、17、20和23 d胚珠; 11~15: 分别代表开花后10、14、17、20、23 d纤维; A: I类LPAT; B: II类LPAT; C: III类LPAT; D: IV类LPAT。Fig. 4 Expression pattern of LPAT family genes in different tissues and organs in G. hirsutum acc. 7235 1: root; 2: stem; 3: leaf; 4: ovules at 0 day post anthesis (DPA); 5: mixtures of fiber and ovule at 5 DPA; 6-10: ovules at 10, 14, 17, 20, and 23 DPA, respectively; 11-15: fibers at 10, 14, 17, 20, and 23 DPA, respectively. A: group I; B: group II; C: group III; D: group IV. |
基于雷蒙德氏棉全基因组数据库, 发掘出8个棉花LPAT家族基因, 属于4个亚类, 分布在6条染色体上。8个LPAT基因在棉花不同组织器官中具有表达特征多样性。LPAT6和LPAT8在发育17天胚珠中表达水平最高, 可能参与油脂合成代谢调控。8个LPAT家族基因均在纤维发育中优势表达, 其中LPAT2、LPAT3和LPAT4 3个基因相对表达量最高, 表明LPAT家族基因参与棉纤维发育进程。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|