


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
[张桂芳1, 2 , 丁在松1 , 赵明1, *  ]
]
]
|
|


稗草( Echinochloa crusgalli)是稻田中的C4光合型杂草, 为了探索稗草 ppc基因(Eppc)对水稻遗传转化的可行性及其对光合速率的调节效应, 首次将含有稗草根型磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase, PEPC)基因 ppccDNA的2个植物表达载体p Ubi-Eppc、p RbcS-Eppc通过农杆菌介导法对水稻进行了遗传转化。对分化植株进行的PCR、RT-PCR、克隆测序和Western杂交等结果均表明稗草 ppc基因已经整合到了水稻基因组中, 并且在转录和翻译水平都得到了表达。转基因水稻PEPC活性和气体交换参数测定结果表明T0代多数植株的PEPC活性高于对照, 最高达到了对照的5.85倍; T0代大多数转基因植株叶片的净光合速率( Pn)比对照提高了20.00%, 最大地提高了47.16%, 同时叶片水分利用效率(WUE)也得到增强; T6代大部分转化植株的PEPC活性及 Pn仍保持高于对照, 本研究表明C3根型 ppc基因过量表达也可以提高水稻的 Pn, 且证明稗草PEPC对光合作用具有较强的调节作用。
Barnyardgrass ( Echinochloa crusgalli) is a C4 weed commonly found in rice field. To fully utilize the photosynthestic potential of Barnyardgrass C4gene, we transformed Barnyardgrass root Phosphoenolpyruvate Carboxylase gene into rice plant with vectors contained promoters of Ubiqitin gene and Rubisco small unit gene by Agrobactirium- mediated transformation. Both marker genes Hygr and ppc were detected by PCR in regenerated plants. RT-PCR and Western blot analysis confirmed that the ppc gene was incorporated into rice plant and expressed with stable transcripts and proteins. PEPC activity as measured in most of the transgenic rice plants was higher than that in control, being up to 5.85-fold of that in untransformed rice. At T0 generation, net photosynthetic rate ( Pn) in most of transgenic rice plants was 20.00% higher than that in untransformed rice, with the highest increase of 47.16%. Water utilization efficiency (WUE) in transgenic rice was also improved. At T6 generation, PEPC activity and Pn of transgenic lines remained higher than those of the wild type. These indicate that over-expressing C3 Eppc gene also can improve rice photosynthesis.
在C4植物的绿色组织中, 磷酸烯醇式丙酮酸羧化酶(PEPC)是参与光合作用CO2原初固定反应的酶。PEPC与CO2有很强的亲和力, 其主要功能是在高温、强光下能把大气中含量很低的CO2以四碳酸的形式固定下来, 进而运输到维管束鞘细胞叶绿体中供C3途径利用。将C4植物的C4型ppc基因导入C3作物如水稻中以提高水稻的光合能力已有多例报道[1, 2, 3, 4, 5]。例如, 来源于玉米和谷子的ppc基因都被成功地导入了水稻, 在一定程度上改善了水稻的光合作用[1, 6]。而稗草作为与水稻共生的主要C4型野生植物, 比水稻具有更强的生长优势, 其ppc基因的利用还鲜有报道。在C3植物及C4植物的其他非光合组织中, 还存在其他形式的PEPC同工酶, 这些同工酶起着碳骨架回补作用, 参与植物的多种生理过程如气孔开放、种子发育与萌发、果实发育等[7, 8]。C3和C4型PEPC除了具有不同的表达特征外, 其蛋白还具有不同的变构调控特征, 以适应其参与的不同生理过程。已有的研究均是将来源于C4植物的C4型ppc基因导入C3植物, 而将来源于C4植物的C3型ppc基因导入水稻中尚未见报道。
我们前期从稗草中克隆了一个属于C3根型的ppc基因[9], 将该基因的cDNA分别与Ubiquitin基因启动子pUbi和1, 5二磷酸核酮糖羧化酶(Rubisco)小亚基基因启动子pRbcS融合构建了2个表达载体, 通过农杆菌介导转化中花8号水稻品种, 建立了稗草根型ppc基因(Eppc)对水稻的转化体系, 获得了一批转基因中花8号水稻植株并对其PEPC活性和气体交换特征进行了检测。
用于遗传转化的水稻品种中花8号、菌株、质粒及载体等均由中国农业科学院作物科学研究所实验室保存; 试剂中的酶类购自TaKaRa公司; 抗生素类购自Sigma; 乙酰丁香酮购自Aldrich。
1.2.1 根癌农杆菌LBA4404的转化 2个植物表达载体pUbi-Eppc和pRbcS-Eppc中, 连接稗草根型ppc基因的启动子分别为pUbi和pRbcS、终止子为nos, 采用三亲杂交法将pUbi-Eppc、pRbcS-Eppc分别转入根癌农杆菌LBA4404。
1.2.2 水稻的遗传转化 水稻种子去除颖壳后, 用次氯酸钠消毒30 min; 然后接种于诱导培养基(NB培养基, 脯氨酸500 mg L-1、谷氨酰铵500 mg L-1、酪蛋白水解物300 mg L-1、蔗糖30 g L-1、2, 4-D 2 mg L-1、植物凝胶2.6 g L-1、pH 5.8)诱导愈伤组织生长, 每10 d继代1次。按照Toki的方法[10]进行水稻的遗传转化。
1.2.3 GUS报告基因组织化学检测 取转化处理后待分化的愈伤组织块和新分化苗(T0代)的幼根(切段), 在新配置的GUS染色液中37℃浸泡1~2 h, 观察颜色变化情况。
1.2.4 PCR扩增和测序鉴定 提取转基因T0代植株总DNA, 对标记基因潮霉素抗性基因(Hygr)用引物Shyg (5'-AAA AAG CCT GAA CTC ACC GC-3')和Ahyg (5'-ACT TCT ACA CAG CCA TCG GT-3')扩增; Eppc基因用引物P1C(5'-ATC CGC AGA ACC CCT CCC ACT CCT CAA G-3')和P2C(5'-GGC GTT TCT CCT CCG ACC ACT CAG CAT A-3')扩增。将扩增后的目的基因片段切胶并回收测序。
1.2.5 RT-PCR检测 分别提取转基因T0代水稻植株和非转基因中花8号(作对照)的总RNA, 进行反转录(Reverse Reaction) PCR扩增, 同样以P1C和P2C为扩增目的基因引物, 同时以Sactin(5'-CCC TCC CAC ATG CTA TTC T-3')和Aactin(5'-AGA GCC TCC AAT CCA GAC A-3')为引物扩增Actin基因作为内参。
1.2.6 Western杂交 提取转基因T0代水稻植株总可溶性蛋白, 通过SDS-PAGE电泳分离不同大小蛋白质, 转移至硝酸纤维素膜上依次用一抗(大豆PEPC)和二抗(碱性磷酸酶标记羊抗兔IGg)进行杂交, NBT/BCIP显色。
1.2.7 叶片PEPC活性的测定 采用分光光度计法, 反应体系总体积1 mL, 组分为: 100 mmol L-1 Tris-HCl (pH 8.0), 5 mmol L-1 MgCl2, 3 mmol L-1 PEP, 0.2 mmol L-1 NADH, 10 mmol L-1 NaHCO3, 10 U MDH, 以加入20 μ L酶粗提物来启动反应, 检测340 nm的吸光值下降的速率, 以单位质量可溶性蛋白来标定PEPC活性。
1.2.8 气体交换参数测定 用LI-COR-6400型光合系统测定, 设定光强为1200 μ mol m-2 s-1, 环境温度30℃左右。分别测得净CO2同化速率(Pn)、气孔导度(Gs)、细胞间隙CO2浓度(Ci)、蒸腾速率(E)。叶片水分利用效率(WUE)为同步测定的Pn与E的比值。
接种在诱导培养基上的脱颖壳消毒后的成熟种子根伸长至3 cm左右时, 愈伤组织块在根部形成(图1-A); 转化后接种在筛选培养基上的大部分愈伤组织在短时间内发生褐化, 部分愈伤组织块约4周左右有新的黄色愈伤组织生成(图1-B); 经过抗性筛选后的愈伤组织接种至分化培养基上光照培养1周后产生绿色分化组织, 继续培养数周分化出绿色小幼苗(图1-C); 将幼苗转移到壮苗培养基上培养2周(图1-D)后成为具有根茎叶的完整水稻植株。2个载体pUbi-Eppc和pRbcS-Eppc分别获得转基因水稻植株18株和16株。
 | 图1 愈伤组织诱导、转化及培养 A: 成熟胚愈伤组织诱导; B: 转化愈伤组织的筛选; C: 幼苗分化; D: 壮苗与生根。Fig. 1 Calli induction, transformation and cultivation A: calli inducement from mature embryo; B: calli transformation and selection; C: growth of regenerated plants; D: Rooting and seedling strengthening. |
分化前挑取愈伤组织块在GUS染色液处理后大部分愈伤组织出现了面积大小不同的蓝色晕斑; 对所获得的T0代34株水稻苗幼根进行Gus基因组织化学检测发现: 转入2个载体pUbi-Eppc和pRbcS-Eppc的幼苗分别有16株和14株呈蓝色。表明带有目的基因的表达载体已转入水稻, 报告基因在所检测88.2%的中花8号转化苗中得到了的表达。
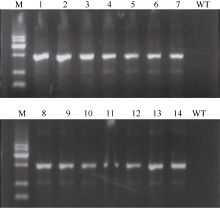
对Gus报告基因检测呈阳性的所有植株进一步的PCR鉴定发现, 30株均能扩增出约1000 bp的筛选标记基因Hygr的片段(图2为部分植株的PCR检测结果), 而未转基因的对照(WT: 中花8号)均无目的条带。
 | 图2 筛选标记基因Hygr的PCR检测 泳道1~7为pUbi-Eppc载体的转基因植株, 8~14为pRbcS-Eppc载体的转基因植株; M为标准DNA长度, 依次为300、500、1000、1500、2000和2500 bp, WT为未转化的品种中花8号。Fig. 2 PCR detection of Hyg resistance gene in transgenic rice Lane 1-7: Transformants of vector pUbi-Eppc; Lane 8-14: Transformants of vector pRbcS-Eppc; M: DNA marker, being 300, 500, 1000, 1500, 2000, 2500 bp in length; WT: untransformed Zhonghua 8. |
对导入的目的基因Eppc的PCR扩增发现, 30株Gus检测和Hygr扩增检测呈阳性的植株多数能扩增出Eppc基因0.8 kb的片段, 2个载体pUbi-Eppc和pRbcS-Eppc分别有13株和12株呈阳性(图3显示了载体pRbcS-Eppc转基因植株的PCR扩增结果), 对扩增条带回收后测序发现序列与GenBank 数据库中已登录的稗草ppc基因[8]序列完全吻合。说明扩增出的片段的确为Eppc基因片段, 进一步证明Eppc基因已经整合到了水稻基因组内。
 | 图3 转基因水稻中Eppc基因的PCR扩增检测 泳道1~12为pRbcs-Eppc转基因水稻样本扩增产物, WT为对照, M为DNA标准长度, 依次为500、1000、2500、5000、7500 bp)Fig. 3 PCR detection of Eppc gene in transgenic rice Lane 1-12: Transformants of vector pRbcS-Eppc; M: DNA marker, being 500, 1000, 2500, 5000, 7500 bp in length; WT: untransformed Zhonghua 8. |
在已扩增出Eppc基因的转化水稻中(pRbcS-Eppc), 随机取其中7株的RNA样本进行RT-PCR检测, 7株转基因样品Eppc基因的表达量存在差异, 且都明显比对照高, 6号样品的扩增量相对比其他样品稍低, 但高于对照; 内参基因Actin的表达量在转基因样品和对照中花8号(WT)之间基本无差别(图4)。
 | 图4 转基因水稻中Eppc基因的RT-PCR检测 泳道1~7为不同的转pRbcS-Eppc基因水稻植株, WT为对照中花8号。Fig. 4 RT-PCR analysis of Eppc in transgenic rice Lane 1-7: transformants of vector pRbcS-Eppc; WT: untransformed Zhonghua 8. |
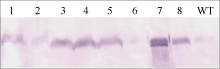
从转入2个表达载体pUbi-Eppc和pRbcS-Eppc的水稻植株中分别取4株进行Western杂交检测, 结果如图5。8个转化株的PEPC蛋白的表达量存在很大的差异。其中2号和6号样品比对照(WT, 中花8号)略低, 其他6个转基因样品的PEPC表达量均明显高于对照, 7号样品最高。
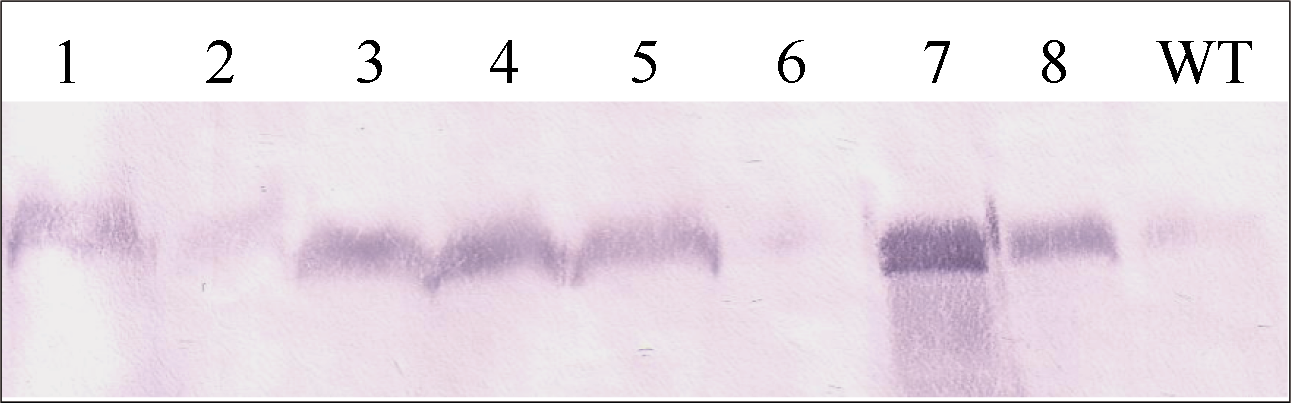 | 图5 转基因水稻PEPC蛋白的Western检测 泳道1~4和5~8分别为pUbi-Eppc和pRbcS-Eppc载体的转基因植株, WT为对照中花8号。Fig. 5 Western analysis of PEPC in transgenic rice plants Lane 1-4: transformants of vector pUbi-Eppc; Lane 5-8: transformants of vector pRbcS-Eppc; WT: untransformed Zhonghua 8. |
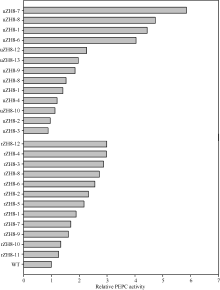
测定了25株(Eppc PCR检测为阳性)转基因水稻(T0代)的PEPC活性(表1)。除2株(uZH8-2和uZH8-3)的PEPC活性(分别为0.0389和0.0358 μ mol mg-1 min-1)低于WT (0.0405 μ mol mg-1 min-1)外, 其余22株的PEPC活性均高于WT, 其中PEPC活性最大的1株(编号: uZH8-7)为0.2371 μ mol mg-1 min-1, 达到了对照的5.85倍。如图6所示的转ppc基因水稻叶片的PEPC相对活性, 2种启动子中以Ubiquitin启动子启动的ppc基因表达效率更高, 相当一部分转化株的PEPC活性比对照提高4倍以上, 而Rubisco小亚基启动子提高的活性不超过3倍。
| 表1 转Eppc基因水稻的PEPC活性和气体交换参数 Table 1 PEPC activity and gas exchange parameters of transgenic rice plants |
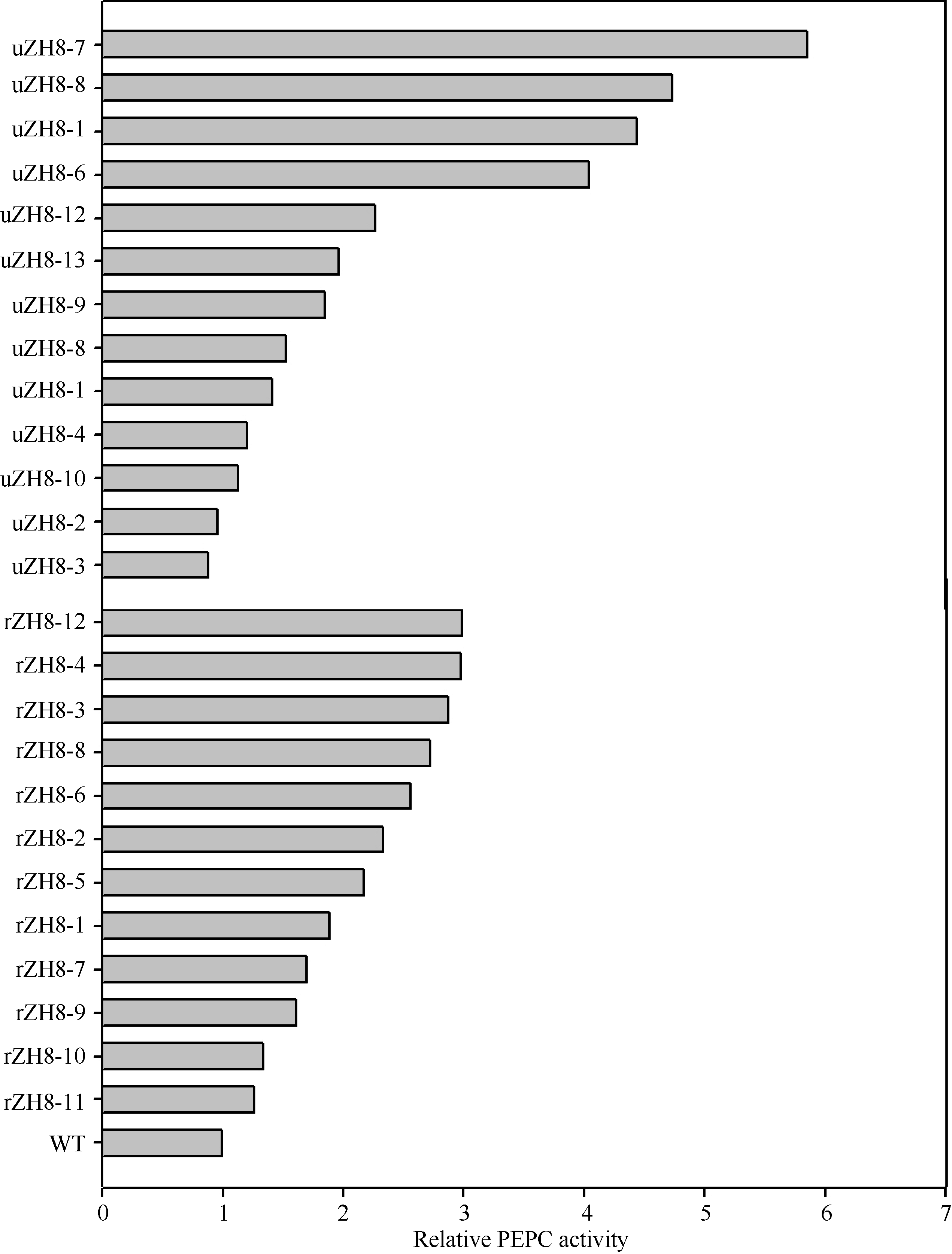 | 图6 转稗草ppc基因植株的PEPC相对活性 uZH8-n: pUbi-Eppc的转化苗; rZH8-n: pRbcS-Eppc的转化苗; WT: 中花8号。Fig. 6 Relative PEPC activity improved by transgenic of Eppc in the control of different promoters uZH8-n: transformants of vector pUbi-Eppc; rZH8-n: transformants of vector pRbcS-Eppc; WT: untransformed Zhonghua 8. |
25株T0代转基因水稻植株的气体交换参数Pn、Gs、Ci及E都存在差别(表1); 其中4株的Pn与对照的差异小于10%, 绝大多数植株(共计15株)的Pn都比对照提高20%以上。说明PEPC活性的增加确实提高了水稻的Pn。
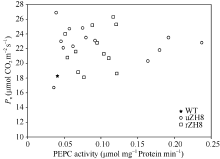
分析T0代光合速率的增加幅度与PEPC活性增加程度并没有显著的相关性(图7)。由Pn/E计算得到的WUE在所有T0代转基因植株中都得到了不同程度的增加。
| 表2 转Eppc基因水稻的PEPC活性及气体交换参数(T6代) Table 2 PEPC activity and Gas exchange parameters of transgenic rice plants (T6 generation) |
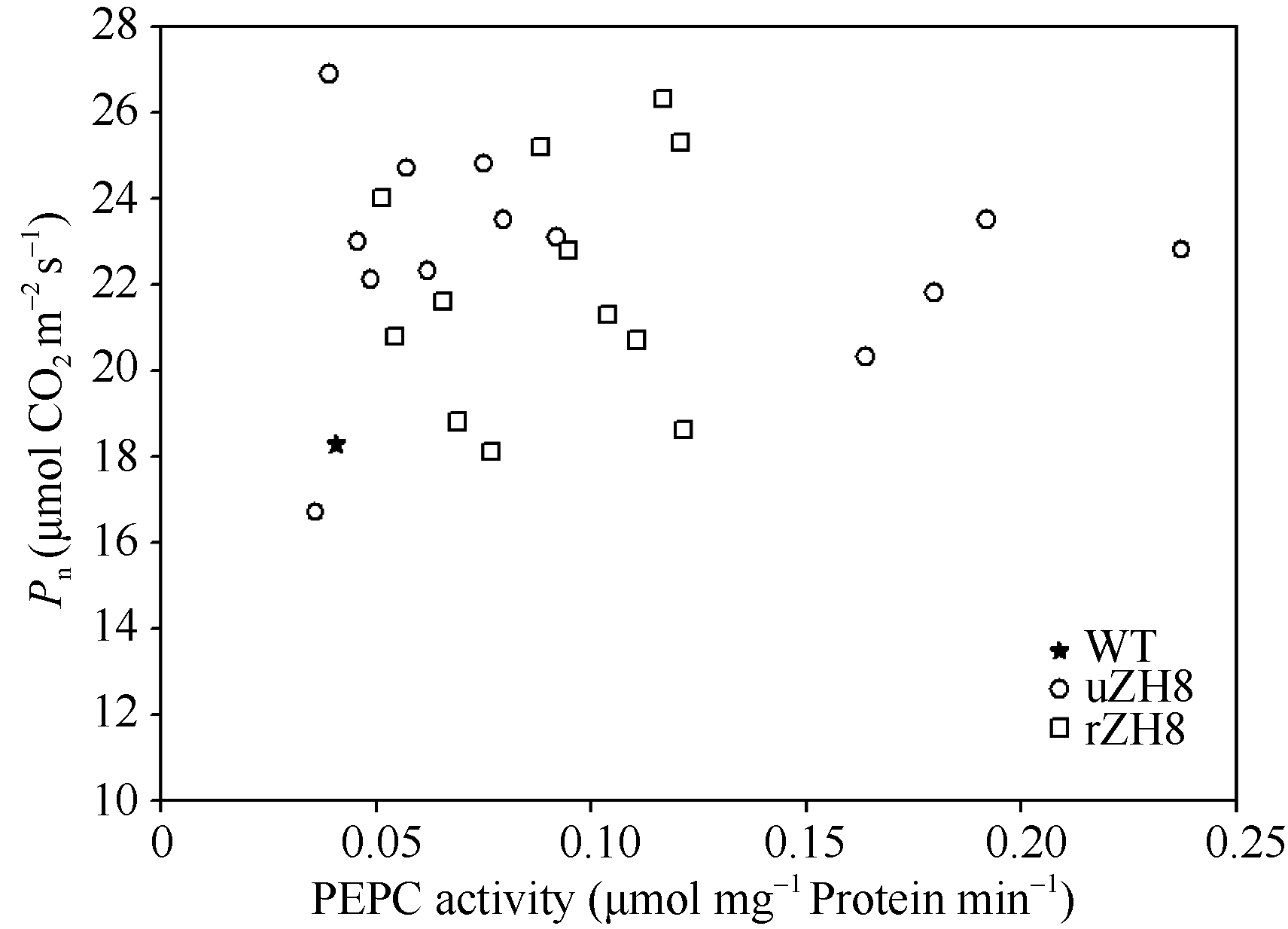 | 图7 PEPC活性与净光合速率的关系 缩写同图6。Fig. 7 Relationship of PEPC activities and Pn Abbreviations as the same as those given in Table 6. |
经过逐代选育, 测定了10株T6代转基因水稻的PEPC活性(表2), 发现10株的PEPC活性均高于WT, PEPC活性最大的1株达到了对照的6.676倍。相应编号T6代花后20 d植株的气体交换参数测定结果见表2, 10株中只有1株(rZH8-12)的Pn低于对照, 其他9株的Pn均高于对照, 其中Pn最高的植株rZH8-4 (17.9 μ mol m-2 s-1)比对照提高了38.76%左右, 此结果证明转稗草根型ppc基因水稻在高代仍保持较高的光合速率。
植物的PEPC由多基因家族编码。植物的不同组织如根、茎、叶、果实、种子等器官[11, 12, 13]都存在着不同类型的PEPC同工酶, 并通过组织特异性表达调节着细胞的各种生理过程。C4光合型ppc基因主要在叶肉细胞中表达, 受光照的调控[14], 在光合组织中主要执行原初CO2固定的功能; 而非光合类型同工酶还具有为硝酸盐同化后的合成氨基酸提供碳骨架、维持细胞质pH和离子渗透平衡、参与种子形成与萌发、果实发育过程中物质代谢、调节气孔运动、为根瘤共生固氮过程提供碳骨架等多种功能[7, 15, 16, 17, 18, 19]。
由于栽培作物玉米和谷子的C4光合型ppc基因导入水稻后, 能使转基因水稻的光合效率不同程度地改善[1, 2, 3, 4, 5, 6], 而稗草是稻田中最常见的C4型野生植物, 与玉米、谷子等栽培作物相比, 其更能适应水、旱2种生存环境条件, 具有更强的生长优势, 因此, 本研究选定稗草PEPC基因对水稻进行转化。根型PEPC同工酶主要在根组织表达, 根型PEPC同工酶活性的增加可提高氨的同化能力[8]。本研究将稗草根型ppc基因分别由强组成型表达的启动子pUbi和受强光调控在叶组织特异性表达的启动子pRbcS调控下, 在转基因水稻叶组织中的表达研究发现: T0代92%的转基因水稻Pn高于对照, 比对照提高30%以上的植株占28%; 通过逐代选育, 到T6代检测的大部分植株Pn仍高于对照, Pn最高的植株比对照提高了38.76%左右, PEPC活性也保持高于对照。表明转稗草根型ppc基因水稻在高代大多数植株仍能保持高的光合速率, 此结果也证明C4植物根型PEPC与叶组织表达的C4型PEPC一样可以对C3植物光合作用具有调节作用。国内学者凌丽俐等利用从美国华盛顿州立大学引进的转C4光合酶基因的水稻, 经过世代繁殖、检测、选择, 得到第8代稳定的种质, 其PEPC活性经测定与第6代种质的PEPC活性相近, 他们的结果也说明PEPC基因可以稳定遗传并高表达[20]。可见在改善C3作物光合作用方面, 与转PPDK、NADP-ME等酶基因水稻植株的光合特性相比, PEPC起到了很关键的作用[21], 而转PEPC+ PPDK双基因水稻植株也未表现比转PEPC基因[22, 23]水稻植株有明显的光合优势[24, 25]。我们转入水稻的是稗草根型PEPC基因, 我们将继续关注根型PEPC对水稻光合生理特性的改善以及在其他代谢方面的调控功能。
由气体交换参数计算得到的WUE可以反映叶片的生理抗旱能力。本研究获得的25株T0代转基因水稻的WUE均高于对照。在玉米中导入高粱C4型ppc基因的研究也证实WUE和抗旱能力得到了提高[26]。我们前面的研究也已发现导入玉米和谷子C4型ppc基因的水稻抗旱能力得到了增强[6, 27, 28, 29]。本研究导入了C4植物的根型(非光合型) ppc基因, 同样也可能提高水稻的抗旱能力, 由表1数据可知, Pn高于对照的转基因水稻中, 78.3%植株的E反而低于对照, 因而提高了WUE, 推断稗草根型PEPC同时也可能调节转基因水稻的羧化能力与气孔运动。但转基因植株的叶面积、绿叶面积的持续期及植株的干重等方面与对照没有明显差异。
转基因水稻的PEPC 活性增加的同时的确也提高了水稻的Pn, 但分析结果显示Pn增加的幅度与PEPC活性增加的程度并没有显著的相关性, 表明PEPC活性的提高被限制在一定水平内可同时提高Pn, PEPC活性的过高或过低反而对Pn的增加或降低影响不明显, 可能是与细胞质内苹果酸的代谢和运转[30]以及苹果酸对PEPC的反馈调节作用有关[31]。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|