

{kind=link}
{kind=link}
{kind=link}
豌豆品系X9002抗白粉病基因鉴定
[王仲怡1 , 付海宁1, 2 , 孙素丽1 , 段灿星1 , 武小菲1 , 杨晓明2 , 朱振东1, *  ]
]
]
|
|


白粉病是豌豆的主要病害之一, 在全球范围内引起严重经济损失。防治豌豆白粉病最有效、经济和环境友好的方法是利用抗病品种。迄今, 2个隐性抗白粉病基因 er1、 er2和一个显性抗白粉病基因 Er3已在豌豆中被鉴定, 其中 er1基因在世界上被广泛应用于抗病品种培育。 er1基因隶属 MLO基因家族, 其抗性由豌豆 PsMLO1基因座位功能丧失产生。X9002是甘肃省农业科学院培育的一个半无叶( afila)抗白粉病豌豆品系。本研究对X9002抗白粉病基因进行鉴定, 开发用于抗白粉病基因选择的分子标记。遗传分析表明X9002对白粉病抗性由1个隐性单基因控制, SSR标记将该基因定位到豌豆第VI连锁群 er1座位区域, 标记AD60和c5DNAmet与其连锁。 PsMLO1基因序列分析发现, X9002存在一个未知大小和身份的片段插入, 该类型突变也发生在含有 er1-2等位基因的豌豆品种Stratagem和Franklin, 表明X9002抗白粉病基因为 er1-2。一个鉴定 er1-2等位基因的功能标记PsMLO1-650被开发, 该标记为互引相标记, 仅在感病植株中扩增, 可以有效用于分子辅助选择。
Powdery mildew is one of the major diseases in pea, causing severe economic loss worldwide. Planting resistant cultivars is the most effective, economical and eco-friendly method for controlling the disease. To date, two recessive resistance genes ( er1, er2) and one dominant resistance gene ( Er3) have been identified in pea, and er1 has been utilized in breeding programs worldwide. Gene er1 is a member of MLOgene family, and er1 resistance is caused by the loss of function at a PsMLO1 locus in pea. X9002 with resistance to powdery mildew is an afilapea line bred by Gansu Academy of Agricultural Sciences. Here, we identified the powdery mildew resistance gene in X9002, and developed molecular marker for the gene selection. Genetic analysis for powdery mildew resistance showed that X9002 carries a recessive resistance gene. The resistance gene was mapped in a region carrying er1 locus on the pea linkage group VI using SSR markers, and was linked to SSR marker AD60 and gene marker c5DNAmet. PsMLO1 sequence analysis revealed that X9002 carries an insertion of unknown size and identity. The same mutation also existed in pea cultivars Stratagem and Franklin carrying er1-2 allele, indicating that the resistance gene is er1-2 in X9002. A functional marker PsMLO1-650 for er1-2 was developed, and the marker was a coupling-phase marker that was detected only in susceptible plants. PsMLO1-650 can be used effectively in marker-assisted selection.
豌豆(Pisum sativum L.)是世界上第三大豆类作物, 在全世界有近90个国家种植[1], 是人类和动物的主要蛋白质来源之一。豌豆在中国已有2000多年的栽培历史, 作为蔬菜、粮食和饲料等原料广泛栽培于全国各地[2]。据联合国粮农组织(FAO)统计, 2013年我国豌豆干籽粒产量138万吨, 居世界第2位, 仅次于加拿大; 此外, 我国是世界上最大的菜用豌豆生产国[3]。豌豆白粉病是制约豌豆生产的最重要病害之一。该病由专性寄生真菌豌豆白粉菌(Erysiphe pisi)引起, 在温带及亚热带地区普遍发生, 特别是在白天温暖、夜间冷凉的气候条件下为害严重, 一般发生年份可造成25%~50%的产量损失, 严重流行时, 高度感病品种的产量损失可达80%以上[4, 5]。白粉病还加速豌豆植株的成熟, 导致青豌豆的嫩度值快速提高。豆荚被严重侵染可导致籽粒变色和苦涩, 品质下降[5]。
化学防治是控制豌豆白粉病常用的方法, 但是杀菌剂的防治效果受喷药时期、次数、环境条件、病原菌抗药性等影响。此外, 使用杀菌剂不但增加生产成本, 而且导致环境污染、残留、食品安全、病原菌抗药性等问题[5]。迄今, 在所有病害防治方法中, 利用寄主自然抗性是最有效、经济和环境友好的方法。1943年Harland[6]从秘鲁的一个豌豆地方品种中发现了一些对白粉病免疫的植株并对这些植株的抗性遗传进行分析, 发现豌豆白粉病抗性受一个隐性基因er控制。1969年2个独立遗传的隐性抗白粉病基因er1和er2被鉴定[7]。随后的遗传分析表明, 大量的豌豆抗性资源或品系含有er1基因[8, 9, 10, 11, 12], er2只在少数几个品种中被鉴定[6, 8]。在细胞学水平上, er1抵抗白粉菌侵入表皮细胞, 表现为免疫或高抗; er2阻止已入侵病原菌在组织中的扩展, 但抗性只在叶片上表现, 且抗性水平受温度、叶龄等条件的影响[13]。最近, 西班牙科学家在豌豆野生种Pisum fulvum中鉴定一个新的显性抗白粉病基因Er3[14]。随着分子标记技术的应用, er1被定位到豌豆遗传图谱的第VI连锁群[15], er2被定位到第III连锁群[16]。由于大量的抗性豌豆资源或品种被鉴定含有er1基因, 许多研究者开展了er1基因分子作图, 不同类型的连锁标记被开发[17, 18, 19, 20, 21, 22], 这些连锁的分子标记为豌豆抗白粉病分子辅助选择育种奠定了基础。er1基因的隐性性质及完全、广谱和持久抗性的特点与大麦抗白粉病Mlo基因相似[23], 也能够通过人工诱变获得[24]。最近, 研究发现豌豆er1基因座位是1个MLO同源基因PsMLO1, PsMLO1功能丧失导致豌豆抗白粉病表现型[25, 26]。由于突变位点和方式的不同, 4个er1等位基因(er1-1、er1-2、er1-3、er1-4)分别在不同的豌豆资源中被鉴定[25]。Santo等[27]通过化学诱变获得一个新的er1突变体er1-5。基于不同分子标记技术, Pavan等[28]建立了5个er1等位基因的功能标记, 但这些标记的有效性尚未被验证。
er1基因在欧洲、北美洲和澳大利亚被广泛地应用到商业育种中, 大量抗病品种的育成和推广, 成功地控制了豌豆白粉病在这些地区的流行[5, 8, 12, 29]。自20世纪80年代以来, 我国相关研究人员开展了豌豆抗白粉病资源的筛选, 鉴定了一批抗性地方豌豆资源和国外引进资源[30, 31, 32]。然而, 迄今豌豆白粉病的抗性在我国还没有被利用, 抗性资源的抗病基因没有被鉴定。豌豆品系X9002是甘肃省农业科学院选育的半无叶皱粒型豌豆。我们前期的温室接种鉴定和多年田间鉴定发现该品系对白粉病表现稳定的抗性[30]。本研究对X9002抗白粉病基因进行鉴定, 开发用于分子辅助选择育种的功能标记, 为有效利用该抗病基因、培育优良的抗白粉病豌豆品种奠定基础。
豌豆品系X9002由甘肃省农业科学院作物研究所杨晓明研究员提供。豌豆白粉病感病品种坝豌6号由河北省张家口市农业科学院徐东旭副研究员提供。以感病的坝豌6号为母本、抗病的X9002为父本配制杂交组合, F1代群体自交产生F2代群体, F2代自交衍生F2:3家系。F1代和F2代群体均种植于本实验室繁殖温室中。
豌豆白粉菌分离物EPBJ由本实验室分离, 用坝豌6号活体保存于10℃光照培养箱。接种体的繁殖采用分生孢子抖落法接种坝豌6号幼苗, 接种后置18~22℃温室培养10~12 d。
1.2.1 亲本抗性鉴定 将抗病亲本X9002和感病亲本坝豌6号分别播种于以粗蛭石为基质的250 mL的纸杯中, 每份材料播种30粒, 每杯播种5粒, 播后置18~26℃的温室培养, 待豌豆苗第3或第4茎节叶片展开时, 采用分生孢子抖落法接种, 其后于18~22℃温室培养。采用0~4级的病情分级标准, 0级为无病; 1级为病斑上有淡薄菌丝层, 可见绿色叶面, 不产生孢子; 2级为菌丝层较厚, 不透绿, 产生一定量孢子; 3级为菌丝层厚, 产孢量较多; 4级为产孢量多, 病斑上的菌丝层全被孢子覆盖[9, 33]。接种10 d后调查发病情况。抗性评价以0~2级为抗病(R), 3~4级为感病(S)。
1.2.2 F1和F2代抗性鉴定 采用离体叶片方法鉴定F1和F2植株对白粉病的抗性。将F1和F2代播种于繁殖温室内, 苗期在每株相同位置取一片复叶, 将叶柄斜插入培养皿内2%水琼脂培养基中, 用分生孢子抖落法接种叶片, 盖上培养皿后用封口膜密封, 置20℃光照培养室培养, 每天14 h光照。10 d后调查各单株叶片的发病情况, 病情分级同前。对表现为免疫和抗病的单株进行重复鉴定。用χ 2测验F2群体抗、感植株分离比。
1.2.3 F2:3家系抗性鉴定 从每个F2:3家系中随机取24粒种子, 分别播种于以粗蛭石为基质的250 mL纸杯中, 每杯播6粒, 播后置18~26℃的温室培养, 待豌豆苗第三或第四茎节叶片展开时, 采用分生孢子抖落法接种, 接种后置18~22℃温室培养, 10 d后调查各家系的发病情况。病情分级同前。对鉴定为纯合抗病和抗性分离的家系进行重复鉴定。对F2:3群体抗性分离比进行χ 2测验。
取坝豌6号、X9002及F2群体单株嫩叶用CTAB 法提取基因组DNA。在F2群体中, 根据抗性鉴定结果随机挑选5个抗病单株和5个感病单株基因组DNA, 将浓度调至为40 ng μ L-1, 分别取等量抗单株DNA或等量感单株DNA混合, 建立抗病基因池和感病基因池[34]。
用于多态性筛选的SSR标记来源于Loridon等[35]绘制的豌豆SSR连锁图谱。从每个连锁群均匀选取10个SSR标记, 在抗、感亲本以及抗、感池之间筛选。用抗、感亲本和抗、感池之间均具有多态性标记鉴定所有F2代单株基因型。根据SSR标记对抗病基因初步定位结果, 在豌豆SSR连锁图谱和豌豆一致性功能图谱(consensus functional map)的相应连锁群上选择分子标记进行多态性鉴定[36], 随后用多态性标记鉴定所有F2代单株基因型。所用引物均由生工生物工程(上海)有限公司合成。SSR标记及基因标记的PCR体系为25 μ L, 含4 ng μ L-1基因组DNA、2 mmol L-1 buffer Mg2+、160 μ mol L-1 dNTPs、0.2 μ mol L-1 M引物对、1.25 U TaqDNA聚合酶。采用Touch Down PCR扩增程序, 94℃预变性3 min; 94℃变性30 s, 60℃退火30 s, 72℃延伸2 min, 循环15次, 每个循环退火温度降低1℃; 接着94℃变性30 s, 45℃复性30 s, 72℃延伸2 min, 循环25次; 最后72℃延伸5.5 min。PCR扩增产物经8%聚丙烯酰胺凝胶电泳检测, 硝酸银染色后观察记录。
用RNAprep植物总RNA提取试剂盒(离心柱型)(天根生化)提取X9002、坝豌6号和奇珍76的总RNA。用BioRT逆转录扩增(RT-PCR)试剂盒(两步法)合成mRNA第1条cDNA链, 用PsMLO1特异性引物对PsMLO1F/PsMLO1R进行PCR扩增[26]。纯化PCR产物并克隆到pZeroBack载体或pMG-T载体(天根生化), 送北京六合华大基因科技股份有限公司公司测序, 利用ClustalX2软件与PsMLO1序列(FJ463618)比对分析获得的序列[25]。
根据X9002和坝豌6号的PsMLO1候选基因序列及PsMLO1序列比对结果, 用Primer Premier 5.0设计功能性标记。由生工生物工程(上海)有限公司合成引物。用X9002和坝豌6号基因组DNA为模板进行PCR 扩增, 检测引物有效性和多态性, 用抗、感亲本间多态性引物鉴定F2群体单株的基因型。PCR体系、程序及产物检测同1.4。
接种豌豆白粉菌分离物EPBJ后, 感病亲本坝豌6号叶片及卷须均被大量分生孢子覆盖, 发病严重度为4级, 表现为感病; 抗病亲本X9002没有出现感病症状, 病级为0, 表现为抗病。以离体叶片接种鉴定9个F1代单株, 结果均为感病。选择含137个植株的F2群体鉴定, 109个植株为感病, 28个植株为抗病, 抗、感植株比经χ 2适合性检验符合1∶ 3的分离比; 在137个F2植株衍生的F2:3家系中, 28个家系为纯合抗病, 72个家系表现为抗、感分离, 37个家系为纯合感病, 经χ 2适合性检验, 纯合抗病、杂合和纯合感病F2:3家系比符合1∶ 2∶ 1的分离比(表1)。抗性遗传分析结果表明, X9002的白粉病抗性由1个隐性基因控制。
| 表1 坝豌6号× X9002杂交组合对豌豆白粉菌分离物EPBJ抗性分离分析 Table 1 Segregation analysis of resistance to isolate EPBJ of Erysiphe pisiin a cross of Bawan 6 × X9002 |
在豌豆分子连锁图谱上均匀选取了148个SSR标记扩增亲本X9002和坝豌6号基因组DNA, 结果表明有68个标记具有多态性。将这些标记用于抗池和感池间多态性标记的筛选, 只有位于第VI连锁群的SSR标记AD60具有多态性, 推测该标记与抗病基因连锁。用AD60鉴定136个F2代植株的基因型, 有36株为抗病基因型, 38株为感病基因型, 62株为杂合基因型, 经χ 2适合性检验符合1∶ 2∶ 1的分离比, 表明AD60为共显性标记。进一步连锁分析表明, AD60与抗病基因连锁。由于AD60位于豌豆遗传图谱第VI连锁群且与er1座位连锁[20, 35], 表明X9002抗白粉病基因可能是一个er1座位的等位基因。
为进一步鉴定X9002抗白粉病基因连锁标记, 将已报道与er1座位连锁的SCAR标记和豌豆一致性功能图谱第VI连锁群SSR标记AD60附近的标记在亲本及抗感池之间进行多态性筛选, 发现5-胞嘧啶DNA甲基转移酶基因(cytosine-5 DNA-methyltransferase)标记c5DNAmet在坝豌6号和X9002间及抗、感池间存在多态性。进一步用F2群体单株进行标记c5DNAmet与抗病基因的连锁验证表明, 该标记与抗病基因连锁, 并在群体中呈现出1∶ 2∶ 1的分离比, 为共显性标记。
对5个坝豌6号PsMLO1同源cDNA克隆的测序表明, 获得的序列与野生型PsMLO1cDNA序列完全相同, 长度为1725 bp, 说明坝豌6号含有白粉病感病基因Er1。10个X9002的PsMLO1同源cDNA克隆的测序表明, 获得了3种明显不同的cDNA序列。与野生型序列比对发现, cDNA1有一个大片段插入和缺失突变, 导致一个155 bp片段的增加; cDNA2序列存在一个129 bp片段缺失; cDNA3除缺失一个129 bp片外, 还缺失一个5 bp的片段(图1)。X9002的PsMLO1同源基因序列与抗白粉病豌豆品种Stratagem (JI 2302)和Franklin相同, 这两品种被鉴定含有豌豆抗白粉病等位基因er1-2[25, 26], 表明X9002抗白粉病基因为er1-2, 其抗性是一个未知大小和身份的片段插入导致PsMLO1基因功能丧失而产生。
根据X9002、坝豌6号PsMLO1同源cDNA序列的差异设计7对引物用于X9002抗白粉病基因鉴定。PCR扩增X9002和坝豌6号基因组DNA, 引物对MLO1-5F/MLO1-5 R (5° -TTTCAGAATGCCTTTC AACT-3° / 5° -CTAGAGCATAAAGAGGCAAA-3° )在坝豌6号中扩增出约650 bp的片段, 在抗病品种X9002中不扩增。进一步用该引物对扩增F2群体单株基因组DNA, 结果全部感病植株扩增一致的约650 bp条带, 抗病植株中没有条带扩增(图2), 表明感病品种或单株中扩增的条带是一个与X9002抗白粉病基因互引相共分离的显性标记, 我们将其命名为PsMLO1-650。该标记可以用于等位基因er1-2的鉴定。
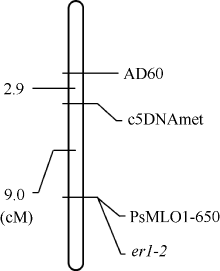
用MapMaker 3.0分析X9002抗白粉病等位基因er1-2和连锁标记之间的遗传距离, 用MapDraw构建遗传连锁图。在构建的连锁图中, 标记AD60和c5DNAmet均位于er1-2的一侧, 遗传距离分别为11.9 cM和9.0 cM, 功能标记PsMLO1-650与er1-2共分离(图3)。
迄今, 除发现一份豌豆近缘种P. fulvum资源对白粉病抗性为显性单基因控制外[14], 几乎所有豌豆抗白粉病遗传分析都表明, 豌豆对白粉病的抗性由一个隐性基因控制[4, 6, 7, 8, 9, 10, 11, 12]。1969年Heringa等[7]在豌豆中鉴定2个独立遗传的隐性抗白粉病基因er1和er2, 其中SVP950和Mexique 4同时含有er1和er2。然而, 随后的研究表明, SVP950和Mexique 4对白粉病的抗性仅由单个隐性基因er1控制[8, 9]。除已证明SVP951、SVP952和JI 2480中含有抗白粉病基因er2外, 大量不同地理起源的抗白粉病豌豆资源及近缘种P. sativumssp. arvense和P. fulvum的抗性均由er1控制[8, 9, 10, 11, 12, 39]。er1基因是大麦感白粉病基因MLO家族成员, 其抗性由感病的野生型PsMLO1功能丧失获得。自然选择或化学诱变导致豌豆er1座位发生碱基缺失、插入、代换、颠换等, 从而产生不同的er1等位基因。迄今已在er1座位鉴定5个等位基因, 即er1-1、er1-2、er1-3、er1-4和er1-5[25, 26, 27]。X9002是甘肃省农业科学院最近选育的豌豆品系, 高抗白粉病。本研究表明, X9002对白粉病的抗性也是由一个隐性基因控制, SSR标记分析将该基因定位到豌豆第VI连锁群上抗白粉病基因er1区域, P00sMLO1候选基因分析和功能标记鉴定确定该基因为er1座位的等位基因er1-2。法国皱粒型豌豆Stratagem是世界上最早鉴定的抗白粉病品种, 作为抗源在欧洲、北美洲、印度等地区和国家被广泛利用, 大量的抗白粉病商业皱粒豌豆品种被育成[29, 40, 41]。1969年Heringa等[7]将Stratagem抗白粉病基因定名为er1。2011年Humphry等[25]通过基因克隆技术进一步确定Stratagem含有er1-2等位基因, 一个未知大小和身份的大片段插入感白粉病基因PsMLO1(Er1)序列导致严重变异的转录子产生, 从而获得对白粉病抗性。X9002为半无叶皱粒型豌豆, 其亲本之一可能来自法国的品系90-PE-10。90-PE-10为半无叶皱粒型豌豆, 抗褐斑病、白粉病等[42, 43], 因此, 我们推测X9002的er1-2等位基因可能来源于Stratagem。
 | 图1 部分野生型PsMLO1编码序列与豌豆品系X9002中鉴定的3个PsMLO1 cDNA同源区域的多重比对Fig. 1 Multiple alignment among partial wild-type PsMLO1coding sequence and homologous regions of three PsMLO1 cDNA sequences identified in line X9002 |
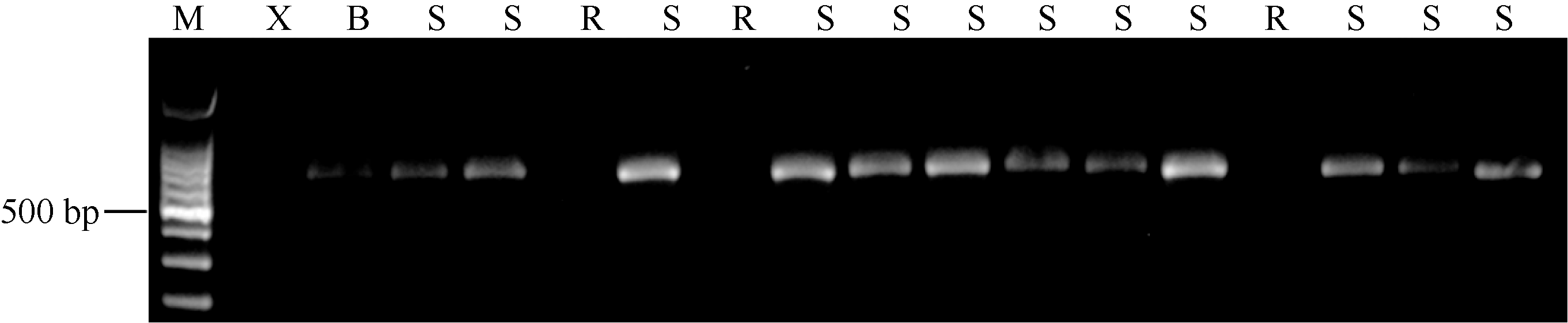 | 图2 功能标记PsMLO1-650对部分坝豌6号× X9002 F2代单株基因型鉴定结果 M: 100 bp DNA marker; X: X9002; B: 坝豌6号; S: 感病单株; R: 抗病单株。Fig. 2 Genotyping partial F2 plants derived from the cross of Bawan 6 × X9002 by functional marker PsMLO1-650 M: 100 bp DNA marker; X: X9002; B: Bawan 6; S: susceptible plants; R: resistant plants. |
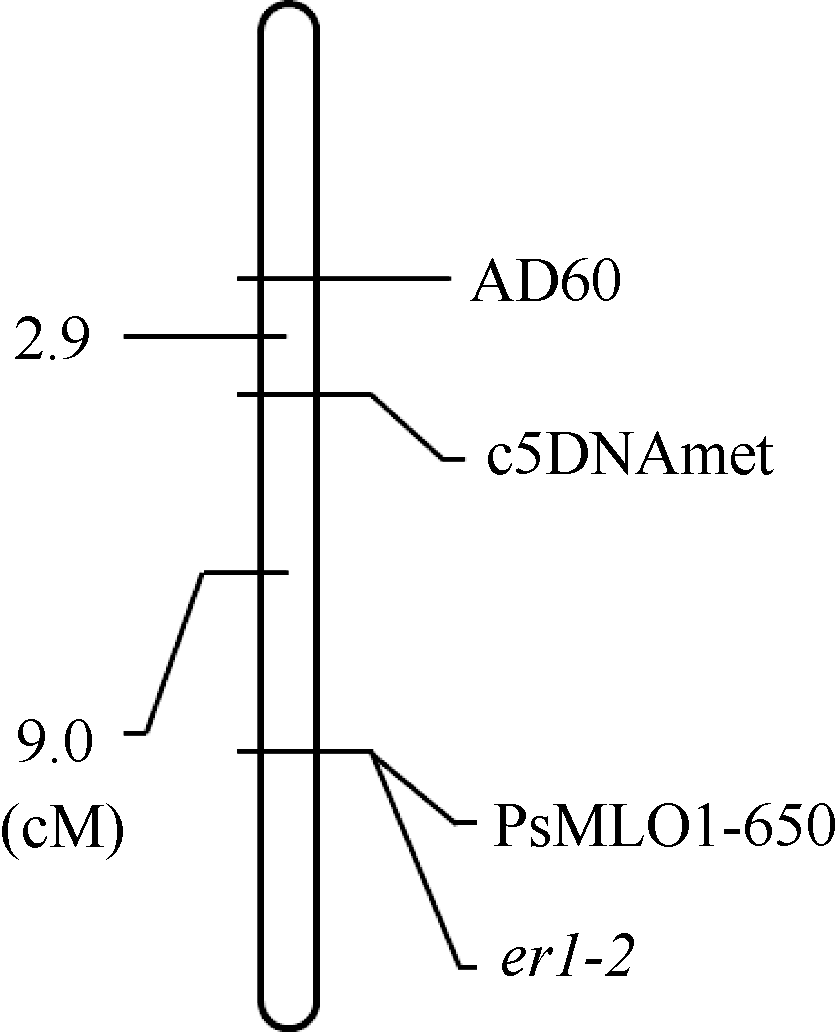 | 图3 豌豆品系X9002抗白粉病基因er1-2和分子标记连锁图Fig. 3 Linkage map of powdery mildew resistance gene er1-2 in pea line X9002 with molecular markers |
Marx[44]发现豌豆抗白粉病基因er与豌豆第III连锁群上的形态学标记Gty具有明显的连锁关系, 据此首先将er基因指派到第III连锁群。1990年Wolko和Weeden[45]将标记Gty放在了豌豆第VI连锁群, 因此er基因也应该位于第VI连锁群。随着分子标记技术发展, 不同类型的分子标记被应用到与er基因的连锁分析和作图。Dirlewanger等[15]利用RFLP标记构建了er1基因第1张分子连锁图, 证明了该基因位于第VI连锁群。随后许多研究者对er1基因进行了分子作图和连锁标记的开发, 但不同研究者获得的er1基因连锁标记不尽相同, 且相同连锁标记与er1基因的遗传距离也存在较大差异[4]。Timmerman等[17]开发了与er1共分离的SCAR标记ScOPD10650, 但Tiwari等[18]发现该标记不能有效地鉴定加拿大豌豆种质。用RAPD标记和Highlight (R)× Radley(S)组合衍生的F3群体作图, Tiwari等[18]获得了3个与er1连锁的RAPD标记, 其中标记OPO181200与er1基因互引相连锁且共分离, OPE161600和OPL61900与er1基因互斥相连锁, 遗传距离分别为4± 2 cM和2± 2 cM。随后, 他们将OPO181200和OPE161600转换为SCAR标记ScOPO181200和ScOPE161600并成功用于豌豆资源的鉴定。Janila和Sharma[19]用ScOPD10650、ScOPO181200和ScOPE161600鉴定印度起源的豌豆种质, 发现ScOPD10650只有80%的可靠性, 而ScOPO181200和ScOPE161600不能区分抗、感品系, 连锁作图表明ScOPD10650与er1基因的遗传距离为3.4 cM。Ek等[20]以Majoret(S)× 955180(R)组合F2群体为材料, 获得了5个与er1基因连锁的SSR标记, 其中SSR标记AD60与er1基因的距离最近, 为10.4 cM。Pereira等[21]利用ISSR、RADPs和AFLPs技术鉴定化学诱变等位基因er1mut2的连锁分子标记, 所用标记还
包括3个SCAR标记ScOPD10650、ScOPO181200、ScOPE161600和5个SSR标记A5、AA369、AA374、AD51和AD60, 利用近等基因系和BSA方法共获得16个连锁分子标记, 但3个SCAR标记和5个SSR标记中只有ScOPE161600和A5与er1mut2连锁, 且距离都大于15 cM, 而其他标记不具多态性或不能有效扩增。最近, Srivastava等[22]开发了一个与er1基因紧密连锁的互引相SCAR标记ScOPX04880, 该标记和SCAR标记ScOPD10650分别位于er1基因两侧, 遗传距离为0.2 cM和2.2 cM。在本研究中, 首先用SSR标记鉴定X9002抗白粉病基因连锁标记, 基于连锁的SSR标记AD60将抗病基因定位到第VI连锁群。由于SSR标记AD60已被Ek等[20]鉴定与er1连锁, 遗传距离为10.4 cM, 而本研究获得AD60与X9002抗白粉病基因遗传距离为11.9 cM, 因此推测X9002抗白粉病基因为er1座位等位基因。之后, 用前人开发的4个SCAR标记ScOPD10650、ScOPO181200、ScOPE161600和ScOPX04880对亲本进行鉴定, 但4个标记不能有效扩增或在亲本间扩增单一条带。最后, 选择豌豆一致性功能连锁图谱第VI连锁群上SSR标记AD60附近区域的功能基因标记进行X9002抗白粉病基因连锁标记鉴定, 发现5-胞嘧啶DNA甲基转移酶基因(cytosine-5 DNA-methyltransferase)标记c5DNAmet为抗病基因连锁标记。遗传作图表明, SSR标记AD60和基因标记c5DNAmet位于X9002抗病基因一侧, 遗传距离分别为11.9 cM和9.0 cM。不同研究者开发的与er1基因连锁标记的通用性很差, 其中可能的原因是研究材料的遗传背景存在较大差异。
基于er1座位等位基因间的序列差异和不同分子标记技术, Pavan等[28]开发了用于5个er1等位基因鉴定功能标记, 其中er1-2的功能标记er1-2/MGB是基于基因内部一个未知大小和身份的插入片段设计的STS标记, 该标记在含er1-2豌豆品种Franklin中扩增大约2700 bp片段, 在感病品种Sprinter中不扩增。我们用功能标记er1-2/MGB鉴定X9002和坝豌6号, 发现该标记在这2个品种中不能有效扩增, 其原因可能是X9002与Franklin在基因组水平上存在较大的遗传差异。Pavan等[28]用er1-2/MGB仅鉴定了Franklin一个品种, 并没有在其他含有er1-2品种或作图群体中进行验证。基于PsMLO1cDNA序列差异, 我们重新设计了7对引物开发er1-2功能标记, 获得一个能够在坝豌6号基因组DNA有效扩增而在X9002基因组DNA不能扩增的引物对PsMLO1-5F/ PsMLO1-5R。PsMLO1-5F序列为PsMLO1基因组DNA序列(KC466597)第13外显子和第13内含子的一部分, PsMLO1-5R为PsMLO1基因组DNA序列第14外显子一部分, 预期扩增607 bp, 而在坝豌6号中实际扩增650 bp左右, 表明坝豌6号PsMLO1同源基因组DNA序列与KC466597存在差异。功能标记PsMLO1-650能够准确区分F2群体抗感植株, 可用于以X9002为抗源的豌豆抗白粉病分子辅助选择育种。
豌豆品种X9002对白粉病的抗性是由单隐性基因控制, 遗传作图将该抗病基因定位于豌豆连锁图谱第VI连锁群, SSR标记AD60和5-胞嘧啶DNA甲基转移酶基因标记c5DNAmet与抗病基因连锁。X9002抗白粉病基因为er1座位等位基因er1-2。功能标记PsMLO1-650能够有效区分F2群体抗病和感病植株。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|