
{kind=link}
{kind=link}
{kind=link}
{kind=link}
以谷蛋白GluA-2信号肽增强外源蛋白在转基因水稻胚乳中的表达与积累
[王红梅1 , 张昌泉1 , 李钱峰1 , 辛世文2 , 刘巧泉1, *  , 徐明良
, 徐明良1, 3 ]
, 徐明良|
|


提高外源蛋白在特定目标组织器官中的表达量是转基因植物研究与开发的核心技术之一。谷蛋白是水稻种子中最主要的贮藏蛋白, 其表达具有严格的时空特异性。为进一步研究谷蛋白信号肽序列在指导基因表达中的作用, 本研究克隆了水稻谷蛋白 GluA-2基因的启动子及其信号肽编码序列, 并与 GUS报告基因编码区融合, 构建了分别含有和不含有信号肽的表达载体p13GG和p13GSG; 经农杆菌介导法分别转入同一水稻品种中, 获得了20多个独立转化子, PCR证明外源基因都已整合进了水稻基因组中。Northern杂交结果表明, 融合GluA-2信号肽编码序列可显著提高 GUS基因在水稻胚乳中的转录; 利用GUS特异的抗体进行Western杂交分析, 显示该信号肽序列可显著提高外源蛋白在转基因水稻胚乳中的积累, 但是其所指导表达的GUS蛋白在水稻胚乳中并没有表现出相应的活性, 其机制有待进一步深入解析。相关结果对于水稻品质改良基因工程研究以及以水稻种子作为生物反应器高效表达外源蛋白具有重要的指导作用。
, XU Ming-LiangIt is one of the key important techniques to enhance the expression of foreign proteins in target tissue/organ of transgenic plants. Glutelin is the major component of storage proteins in rice seeds, and its expression was tightly temporal and tissue- specific, which is controlled by several mechanisms. To further reveal the function of the Glutelin signal peptide on expression of target gene, in present study, we isolated the promoter and signal peptide-coding sequences of the glutelin GluA-2 gene, and fused them transcriptionally to the GUS coding sequences. Beside, the construct without the GluA-2 signal peptide-coding sequences was also generated as a control. Both constructs with the GUS chimeric genes, named as p13GSG and p13GG, were introduced into the same rice variety by Agrobacterium-mediated transformation. More than twenty independent transgenic lines were generated for each construct, and the integration of the GUS chimeric gene was confirmed by PCR technique. The results from Northern blot analysis showed that, after fusing the GluA-2 signal peptide coding sequences between the GluA-2 promoter and the GUS coding sequence, the transcription of GUS chimeric gene could be dramatically increased. Then, Western blot was carried out by using the GUS-specific antibody, and the results obviously revealed that the accumulation of foreign proteins was significantly enhanced in the endosperm of transgenic rice with the signal peptide. However, there was no or very low GUS activity in the endosperm of transgenic rice plants with the signal peptide. These results were very useful to improve the grain quality of rice via genetic engineering, especially produce foreign proteins in the seeds of rice as bioreactor.
在水稻种子中贮藏蛋白是仅次于淀粉的第二大类贮藏成分, 绝大多数贮藏蛋白基因的表达具有严格的组织与时空特异性, 并且还具有特定的蛋白靶向元件[1, 2, 3]。贮藏蛋白基因在翻译成蛋白前体时, 其氨基末端都含有一段信号肽(signal peptide)序列, 长度随种类不同而异; 这些信号肽在贮藏蛋白及其mRNA的定位与运输中起重要作用。在贮藏蛋白基因转录出成熟的mRNA后, 首先翻译成含信号肽的前体蛋白, 由信号肽序列牵引定向地转移到粗糙型内质网腔内, 随后这一信号肽序列从前体蛋白上被剪除下来, 以便贮藏蛋白能正确地加工[4]。除了在目标蛋白的正确定向中具有决定作用外, 信号肽序列在调控基因表达水平上也具有重要的作用[5]。信号肽序列的上述特性已在转基因植物研究中获得了很好的应用。例如, 利用菜豆肌动蛋白内质网特异的信号肽序列可明显增强大肠杆菌辅酶Q在转基因烟草中的转录与随后的表达量。由此可见, 贮藏蛋白的信号肽序列对转基因植物中外源蛋白的表达与沉积是有积极作用的。
谷蛋白是水稻胚乳中最主要的贮藏蛋白, 它一般在种子发育的中后期大量合成并积累[6, 7, 8]。虽然它与豆科植物中的11S球蛋白的溶解性能完全不同, 但它们在氨基酸组成、蛋白质结构及其合成途径上有许多相似之处[9, 10], 因此有时将水稻谷蛋白也归为植物的球蛋白大家族中。水稻种子中含有3种不同分子量的谷蛋白, 分别为57 kD的前体谷蛋白、37~39 kD的酸性亚基(具酸性等电点)和20~22 kD的碱性亚基(具碱性等电点), 其中的酸性亚基和碱性亚基是由谷蛋白前体经翻译后加工剪切而成[6]。成熟的谷蛋白都是异质性的, 如谷蛋白酸性亚基至少由12个多肽组成, 而碱性亚基也至少含有9个多肽[10]。根据谷蛋白的氨基酸序列, 可将其分为A和B两个大的亚家族, 分别称为GluA和GluB, 每一个亚家族都由多基因控制; GluA或GluB内各多肽间氨基酸序列的同源性在80%~88%之间, 而两类不同亚家族间多肽的氨基酸序列同源性却不到65%[11]。从谷蛋白基因或其cDNA推导的氨基酸序列分析, 在谷蛋白前体的N-端有一个很典型的由24个氨基酸残基组成的信号肽, 该信号肽序列在引导谷蛋白前体定位于内质网膜, 进而转运到内质网膜腔内的过程中起重要作用[11, 12]。
目前, 多数谷蛋白基因已被分离克隆[11, 12, 13, 14, 15]。在GluA亚家族中至少有3种不同的编码基因, Takaiwa和Oono[15]将它们分别命名为GluA-1、GluA-2和GluA-3, 它们在每个水稻单倍性基因组上都含有5~8个拷贝[12]。另外还有一个基因称为GluA-4, 属于假基因[15]。GluA亚家族内各成员间在基因组织结构及一些特异序列上都很保守[12]。GluA-1与GluA-2的编码区核苷酸序列同源性高达95%, 而GluA-1 (或GluA-2)与GluA-3的同源性为81%。GluA-2是水稻谷蛋白家族的成员之一, 其前体蛋白的氨基末端含有由24个氨基酸组成的信号肽序列, 其编码基因只在发育的水稻胚乳中特异性地表达[12, 13, 14]。
本研究克隆了水稻GluA-2基因的启动子及其信号肽编码序列, 并与GUS报告基因编码区融合, 经农杆菌介导转入水稻中, 以研究利用该基因信号肽序列指导外源基因在转基因水稻中表达的作用。
高产粳稻(Oryza sativa L. subsp. japonica)品种武香粳9号为转化受体品种; 另一粳稻品种武运粳8号基因组DNA作为模板用于克隆GluA-2基因启动子和信号肽编码序列。
载体pGEM-T购自Promega公司, 农杆菌菌株EHA105和双元载体pCAMBIA1301由澳大利亚Jefferson教授提供, 双元载体pC1300/GN由本实验室自行购建[16], 大肠杆菌DH5α 为本实验室保存。限制性内切酶、T4 DNA连接酶等购自TaKaRa公司。
根据Takaiwa等[13, 14]和Okita等[12]发表的谷蛋白基因序列, 在水稻谷蛋白GluA-2前体的N-端含有由24个氨基酸组成的信号肽序列。根据已发表的水稻谷蛋白GluA-2基因的核苷酸序列(GenBank登录号为Y00687), 设计引物Gt1P6(5′ -CAGGATCCAA TGGCATCCATAAATCGCCCC-3′ )和Gt1P10(5′ -CA CCATGGCTAGGGAGCCATCGCACAA-3′ ), 在引物5′ 末端分别加上BamH I和NcoI酶切位点(下画线表示); 以水稻品种武运粳8号基因组DNA为模板进行PCR扩增, 用于克隆GluA-2基因ATG下游+1 ~ +72位碱基之间的信号肽编码序列(称为GluA-2_SP片段, 图1-a)。设计引物Gt1P1(5′ -GGCAAGCTTCA CTGCTACCTTTAAGTAAC-3′ )和Gt1P3(5′ -CGAGG ATCCGTTGTTGTAGGACTAATGAA-3′ ), 从水稻总DNA中扩增GluA-2基因翻译起始密码子ATG上游1.3 kb长的启动子区。在2个引物的5′ 末端分别加上了Hind III和Bam HI限制性内切酶位点(下画线表示)。扩增的启动子片段称为GluA-2_Pro片段。PCR扩增程序为: 95℃、5 min; 95℃、1 min, 50℃、1 min, 72℃、1 min, 30个循环; 72℃、10 min。PCR产物经克隆入质粒pGEM-T后进行序列分析。
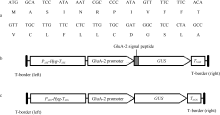
将GluA-2_Pro和GluA-2_SP片段连接在一起, 形成GluA-2_Pro+SP片段。然后, 将该片段经Hind III和NcoI双酶切, 替代双元载体pCAMBIA1301中GUS编码区上游的CaMV35S启动子, 即构建成双元载体p13GSG (图1-b)。该载体中, 在GluA-2_ Pro启动子与GUS基因编码区(保留有自己的ATG序列)之间即含有GluA-2基因的信号肽编码序列, 信号肽编码序列与GUS编码区序列组成一个融合基因, 经测序证明两者的读码框都没有发生改变。同时, 为比较GluA-2基因信号肽编码序列对GUS基因表达的影响, 又构建了只含有GluA-2基因启动子(GluA-2_Pro)的GUS嵌合基因。用Hind III和Bam HI双酶切GluA-2_Pro片段, 连接入质粒pC1300/GN [16]的相应酶切位点中, 构建成含GluA-2_Pro启动子与GUS报告基因相融合的质粒p13GG (图1-c)。构建的载体经冻融法导入农杆菌EHA105中, 并用于水稻转化。
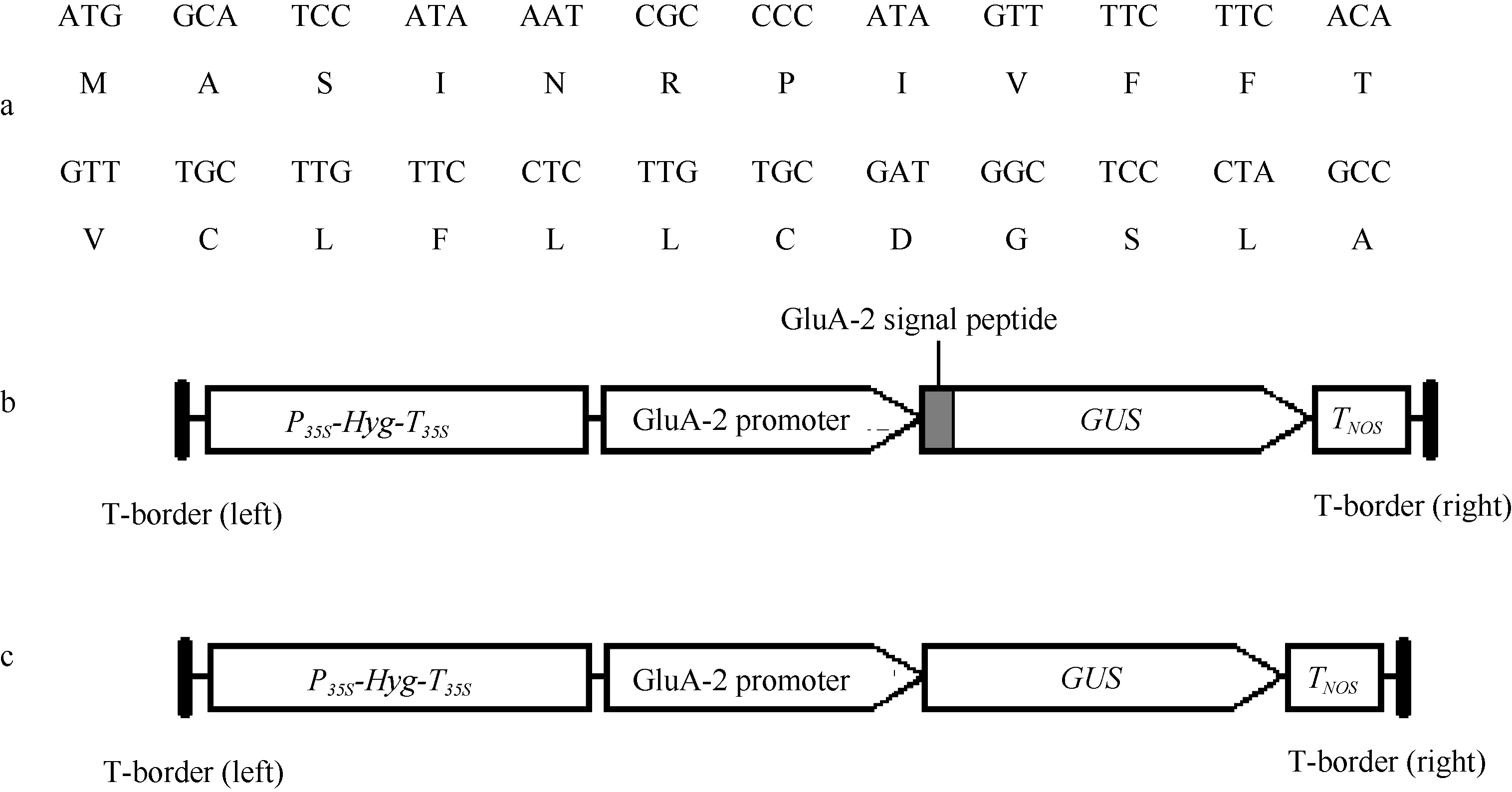 | 图1 谷蛋白GluA-2信号肽序列(a)及双元载体p13GSG (b)和p13GG (c)中T-DNA区的结构 P35S和T35S分别代表CaMV35S启动子和终止子, Hyg代表潮霉素抗性基因, TNOS代表NOS终止子。Fig. 1 Sequences of rice GluA-2 signal peptide (a), and the T-DNA structure of binary vectors p13GSG (b) and p13GG (c) P35S and T35S represent the promoter and terminator of CaMV35S gene, respectively; Hyg means the hygromycin resistance gene, TNOS means the terminator of NOS gene. |
取武香粳9号开花后12 d左右的未成熟胚诱导初生愈伤组织, 作为农杆菌介导转化的受体愈伤组织。按刘巧泉等[17]的方法进行水稻组织培养及其农杆菌介导的转化。按CTAB法[18]从转基因水稻叶片中提取总DNA; 参照刘巧泉等[17]的方法进行GUS编码区片段的PCR扩增, 预期PCR产物片段长为500 bp左右。
按郑霏琴等[19]的冷酚法从水稻开花后12~15 d的未成熟种子中提取总RNA。取5~10 μ g总RNA, 经55℃变性20 min在1.0%琼脂糖/甲醛凝胶分离后, 按Sambrook等[20]和Roche公司推荐的方法将RNA转移至尼龙膜(Roche)上, 在紫外交联仪(Bio-Rad)上于254 nm处将RNA固定到膜上, 再以地高辛(Digoxigenin, Roche)标记的GUS基因编码区序列作为探针进行杂交。
在转基因水稻成熟后, 收取成熟种子磨成米粉, 从其淀粉胚乳中分别提取可溶性蛋白和总蛋白。按每1 mg米粉10 μ L抽提液加入可溶性种子总蛋白抽提缓冲液(50 mmol L-1 Na3PO4, pH 7.0, 500 mmol L-1
NaCl), 在37℃振摇3 h; 然后在15 000转min-1、4℃离心20 min, 吸取上清液至另一新的离心管中, 保存在4℃备用。按Bradford法[21]测定蛋白浓度。按Yamagata等[6]方法提取种子总蛋白。按Sambrook等[20]的SDS-PAGE程序, 取适量种子总蛋白样品与2× 样品缓冲液混合, 99℃变性处理10 min后, 在12%分离胶和5%积层胶组成的聚丙烯酰胺凝胶上于电泳分离; 经SDS-PAGE分离后, 将蛋白质转移到Nitrocellulose膜上。按AURORA Western blot chemiluminescent detection system (AURORA)推荐的程序进行Western杂交, 所用的抗体为GUS特异的兔免疫抗血清(购自Abcam公司), 按1∶ 10 000稀释。
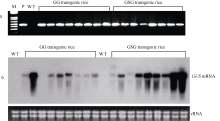
将构建的双元载体p13GSG和p13GG经冻融法分别导入根癌农杆菌菌株EHA105感受态细胞中。按本实验室已建立的农杆菌介导法转化水稻的程序[17], 将所构建的2个GUS融合基因分别导入水稻品种武香粳9号中, 再生的小苗经生根壮苗后移入网室或人工气候室盆栽。两个融合基因构建中, 各获得20个以上的独立转化子。含GluA-2_Pro+GUS融合基因(载体p13GG)的转基因植株称为GG1、GG2和GG20等, 而含GluA-2_Pro_SP+GUS融合基因(载体p13GSG)的转基因植株称为GSG1、GSG2和GSG20等。每个转化子各含有2~6个转基因水稻植株。转基因水稻植株移栽入大田后, 绝大多数能正常生长、开花并结实。经PCR分析证明GUS融合基因已整合进转基因水稻的基因组中(图2-a)。
 | 图2 转基因水稻的PCR (a)和未成熟种子总RNA的Northern杂交分析(b) 图a为转基因水稻植株总DNA的PCR分析, 所用引物为GUS基因特异引物。图b上图为Northern杂交结果, 下图显示总RNA中的28S核糖体RNA。总RNA分别抽提自GG和GSG类转基因水稻植株或未转化植株(标记为WT)的未成熟种子。杂交所用的探针为地高辛标记的反义GUS编码区片段。Fig. 2 PCR (a) and Northern blot (b) analyses of transgenic rice Panels a and b show the PCR analysis using GUS specific primers of total leaf DNA and Northern blot analysis of total RNA from developing seeds of rice plants, respectively. WT means wild type plant, GG and GSG mean the transgenic plants derived from the GG and GSG constructs, respectively. The probe for Northern blot is DIG-labeled anti-GUSDNA fragment. |
在转基因水稻开花后12 d左右, 从每一植株上收取40粒左右未成熟种子, 提取总RNA, 并用与GUS基因编码区序列特异的反义DNA探针进行Northern杂交分析, 结果显示在两类转基因水稻植株未成熟种子中都有较高的GUS基因的转录本(图2-b)。从杂交信号的强弱判断, 虽然在不同转化子间的表达量有较大的差异, 但从总体情况分析, 在GSG转基因水稻中, 与GluA-2信号肽序列融合的GUS嵌合基因的表达水平明显高于GG类转基因水稻种子中GUS嵌合基因(即不含信号肽序列、只含GluA-2启动子序列的GUS嵌合基因)的转录水平, 说明有GluA-2信号肽序列的存在, 可提高目标基因在转基因水稻胚乳中的转录水平(图2-b)。
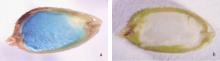
在转基因水稻植株开花后15 d左右, 从已检测为GUS基因阳性的转基因植株上分别取叶片、茎和未成熟种子进行GUS组织化学染色分析。结果显示, 对于GG类转基因水稻植株, 在未成熟种子胚乳中能观察到很强的GUS活性(图3-a); 而在叶片、茎和胚等组织中, 只有极少数转基因植株有GUS活性, 并且较弱, 而绝大多数观察不到GUS蓝色。显示GluA-2启动子可指导外源基因在水稻胚乳中特异性表达, 这与之前的研究相一致[12]。但是, 在GSG转基因水稻不同组织中都没有检测到GUS活性(图3-b), 暗示与GluA-2信号肽序列融合的GUS嵌合基因虽然可以在转基因水稻种子中高效转录, 但是检测不到外源蛋白的活性表达。
 | 图3 转基因水稻植株GG (a)和GSG (b)发育种子中GUS活性分析Fig. 3 GUS activity analysis of developing seeds of GG (a) and GSG (b) transgenic rice |
在转基因水稻成熟后, 收取成熟种子, 从其胚乳中提取种子总蛋白, 用抗GUS的特异抗体进行Western杂交分析, 结果显示在两类转基因水稻成熟种子中都有GUS蛋白的积累(图4-a)。在不同转基因水稻种子中GUS蛋白的表达量有所不同, 这与Northern杂交的结果相类似。但从总体趋势上分析, 在GSG类转基因水稻胚乳中GUS蛋白的积累量明显高于GG类转基因水稻中的(图4-a)。进一步先从转基因水稻成熟种子中提取可溶性蛋白, 然后再从沉淀物中分离出总蛋白, 分别经SDS-PAGE分离后进行Western杂交分析。结果显示, 在GG类转基因水稻成熟种子可溶性蛋白中可检测到GUS蛋白的存在, 而在GSG类转基因水稻种子中可溶性蛋白中却检测不到GUS蛋白(图4-b)。在已不含可溶性蛋白的总蛋白提取物中, 两类转基因水稻中都有较强的GUS蛋白积累, 但是GSG类转基因水稻种子中GUS蛋白的积累量要显著高于前者(图4-c), 这与种子中总蛋白的分析结果相一致。
综合上述结果, 当在GUS基因编码区前加接上GluA-2基因的信号肽编码序列时, 不仅可明显促进GUS融合基因的转录效率, 而且可使最终的蛋白产物表达量也明显增多。对Northern和Western杂交中各杂交信号进行扫描定量比较分析, 可估算在GSG类转基因水稻胚乳中GUS蛋白的表达量比GG类转基因水稻中的要高5~10倍。
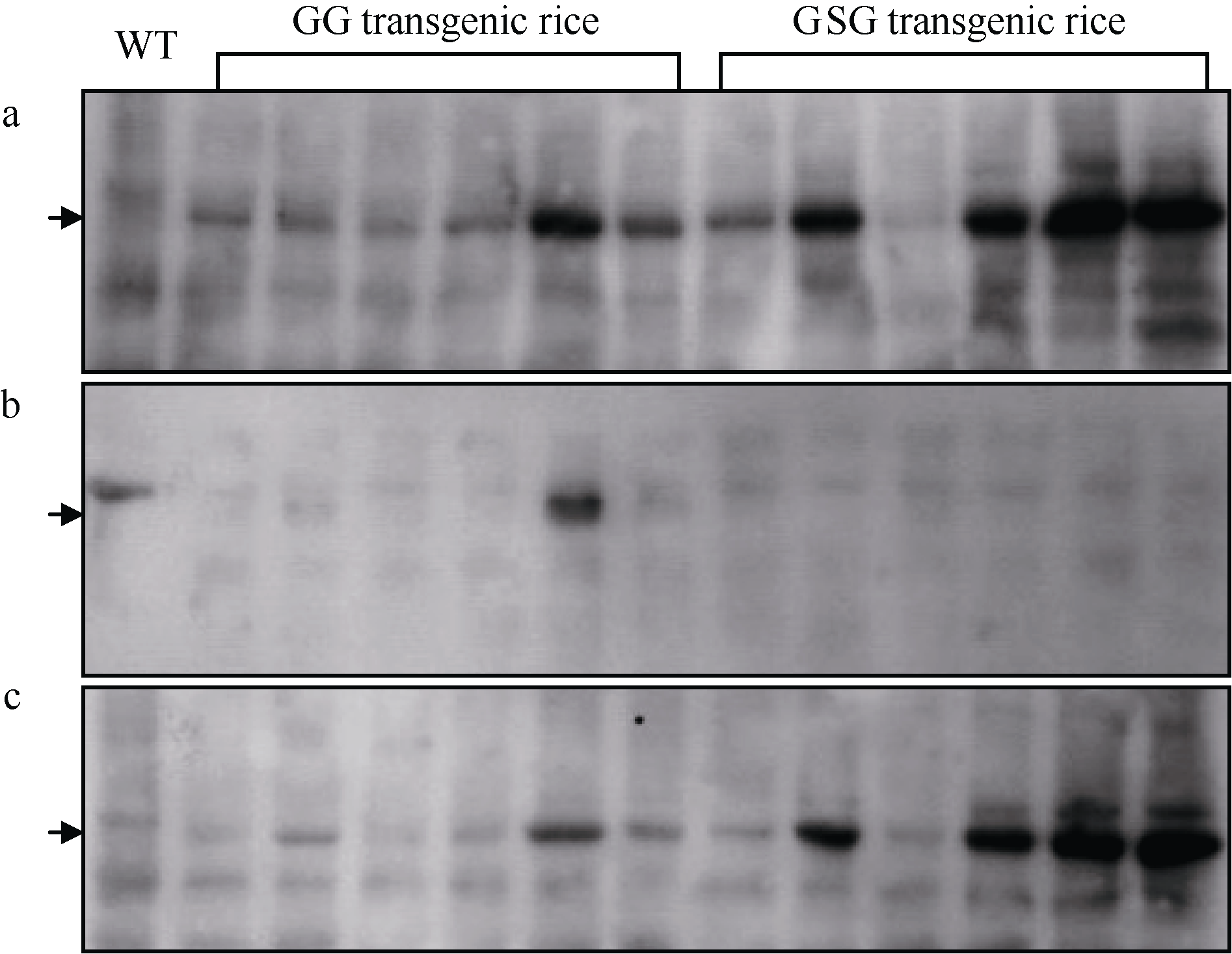 | 图4 转基因水稻植株成熟种子蛋白的Western杂交分析 种子蛋白分别抽提自GG和GSG类转基因水稻植株或未转化植株(WT)的成熟种子, 其中a为种子总蛋白, b为种子可溶性蛋白, c为去除可溶性蛋白后的其他蛋白。每个泳道所加的蛋白量一致。图左侧箭头所指为GUS蛋白。Fig. 4 Western blot analyses of proteins from mature seeds of transgenic rice Proteins were extracted from the mature seeds of GG and GSG transgenic plants or their wild type (WT), respectively. Panels a, b, and c mean the total proteins, soluble proteins and the remaining total proteins after extraction of soluble proteins, respectively. The amount of proteins for each lane is same. The arrow at left indicates the expressed GUS protein. |
植物种子贮藏蛋白基因在翻译成前体蛋白时, 其N端都含有一段信号肽序列, 长度随物种和贮藏蛋白种类的不同而异, 它们在贮藏蛋白及其mRNA分子的定位及运输中起重要作用[4, 23]。水稻醇溶蛋白mRNA上与信号肽对应的序列在将该mRNA定位到特定的内质网膜上的过程中起着决定作用[24]。本研究将谷蛋白GluA-2的信号肽编码序列融合在GUS编码区的N-端, 并与GluA-2启动子组成融合基因, 在转基因水稻未成熟种子中的GUS转录本要明显高于不含此信号肽编码序列的转基因水稻(图2-b), 说明这一序列的存在可增强外源基因的转录。Boehm等[25]的研究指出, 菜豆肌动蛋白的内质网特异的信号肽序列可增强大肠杆菌辅酶Q在转基因烟草中的转录与随后的表达量。Wright等[26]利用水稻谷蛋白Gt3的信号肽序列将人细胞巨化病毒的糖蛋白B成功地定位在转基因烟草种子中的蛋白贮藏囊泡中。由此可见, 包括谷蛋白GluA-2等在内的多数贮藏蛋白的信号肽序列对于在转基因植物中外源蛋白的表达与沉积是有积极作用的。
但在本研究获得的含GluA-2信号肽编码序列的转基因水稻植株中, 却检测不到或只检测到极微弱的GUS活性, 可能与以下两点有关: (1)含信号肽编码序列的GUS转录本在翻译后形成了仍含信号肽序列的GUS融合蛋白, 但该信号肽序列不能被正常剪除下来, 因而影响了GUS蛋白在随后的加工过程(如糖基化等), 进而影响了它的酶活性。Coleman等[4]的研究发现在玉米的fleury-2突变体中, 信号肽序列不能从α -玉米醇溶蛋白(α -zein, 与水稻醇溶蛋白具有极高的同源性, 属于同一类植物贮藏蛋白)的前体上剪切下来, 从而影响了它的加工过程, 因此在该突变体中的α -玉米醇溶蛋白的量明显减少, 与其相应的蛋白体的量也减少。(2)在GUS前体蛋白形成后, 由于有信号肽序列的存在, 使GUS融合蛋白进入内质网的内腔中, 并在这里进行加工, 最后像醇溶蛋白一样沉积到蛋白体中, 从而影响了其酶活性的发挥。Choi等[3]的研究指出, 将水稻醇溶蛋白基因中的信号肽编码序列除去, 可有正常的醇溶蛋白转录本产生, 但却检测不到最后的蛋白。因此, 他们认为信号肽序列对于醇溶蛋白的稳定及其定位是非常重要的。由此, 我们可推测, 不含信号肽序列的GUS蛋白可能没有进入内质网腔内, 而仍留在细胞质中, 从而能被检测出来; 而当加上信号肽序列后, 它进入内质网腔中, 就很难被检测出来。此外, 本实验室利用相似的表达策略, 利用GluA-2信号肽指导一些分子量较小的外源蛋白在水稻胚乳中表达, 结果显示确可显著提高外源蛋白的表达量, 而且所表达出的外源蛋白仍具有预期的活性(结果待发表)。但是, 究竟是什么原因导致这种表达现象, 尚需进一步试验证实。
利用转基因技术提高植物抗逆性、增加产量和改善品质等已成为现代农业生物技术研究开发的重点与热点[27, 28, 29]。尤其是近年来, 利用植物这一廉价生产系统作为生物反应器, 来大量生产重组的编码具重要功能的异源蛋白如医用活性多肽或疫苗、抗体等, 是当前植物基因工程发展的一个重要领域之一[30, 31, 32, 33, 34]。但是, 在植物生物反应器研究中存在一个较为突出的问题, 即目标蛋白的表达量极低, 某些蛋白还不能稳定积累下来[30, 31]。因此, 建立高效稳定的外源蛋白表达技术体系历来是该领域研究的重点与难点。细胞代谢合成的蛋白质在植物与动物中贮存的方式有所不同, 要实现外源重组蛋白在转基因植物细胞中高效表达与稳定积累, 需要遵循与植物本身蛋白一致的表达与贮存规律。其中关键技术之一便是要应用受体植物来源的基因表达调控元件和目标蛋白靶向序列。因此, 在以水稻种子或胚乳为目标器官表达外源蛋白时, 使用像谷蛋白这样的胚乳特异性表达基因的启动子及其N-端信号肽编码序列, 是实现目标蛋白在水稻种子组织中高效表达的有效途径。
克隆了水稻谷蛋白GluA-2基因的启动子及其信号肽编码序列。结果表明, 融合GluA-2信号肽编码序列可显著提高GUS基因在水稻胚乳中的转录, 进一步可显著提高外源蛋白在转基因水稻胚乳中的积累。相关结果对于水稻品质改良基因工程研究以及以水稻种子作为生物反应器高效表达外源蛋白具有重要的指导作用。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|