
{kind=link}
{kind=link}
{kind=link}
{kind=link}
花后冠层温度对小麦产量的影响及几个关联SSR位点的效应分析
[张冬玲, 张洪娜, 郝晨阳, 王兰芬, 李甜, 张学勇*  ]
]
]
|
|


小麦生育期间的冠层温度, 尤其是开花以后的冠层温度对植株衰老、粒重和品质等有很大影响。2012—2013和2013—2014年度, 在河南新乡利用4个品种的选择导入系群体观测了冬小麦冠层温度与产量间的关系, 并通过全基因组SSR标记检测相关主效基因位点, 以解析其遗传机制。结果表明, 冠层温度与千粒重和产量呈显著负相关; 与有效穗数呈显著正相关, 并随着时间的推移, 相关性逐渐增大。在已报道的44个与千粒重或穗粒数相关联的SSR位点中, 有9个与冠层温度显著关联, 其中在6个位点上检测到优异等位变异, 携带优异等位变异材料的冠层温度显著低于携带非优异等位变异材料的冠层温度。影响冠层温度的优异等位变异间存在明显的加性效应, 这些位点同时也与千粒重和穗粒数呈现出显著关联。与冠层温度关联的SSR位点均与灌浆中后期旗叶的叶绿素含量相关联。由此表明, 在灌浆后期良好的根系功能保证了水分和营养的正常供应, 降低了冠层温度, 而较低的冠层温度对叶绿素和光合起到了良好的保护作用, 从而提高千粒重和增加籽粒产量。
The canopy temperature (CT) during wheat growing period, especially after flowering, has a major impact on plant senescence, grain weight and quality. In this study, the relationship between CT and yield was analyzed using selected introgression lines derived from more than 60 accessions of Chinese wheat mini core collection crossed and backcrossed by four wheat varieties. The CT was negatively correlated with thousand-kernel weight and kernel yield, and positively correlated with spike number per plant. The correlation coefficient gradually became larger in the process of time. Lower CT during later grain-filling protected chlorophyll and photosynthesis, increasing thousand-kernel weight (TKW) and yield. Among the 44 loci associated with grain number (GN) and TKW detected previously, nine were also associated with CT. Favored alleles were detected on six loci. The CTs of varieties containing favored alleles were lower than those of varieties containing other alleles. Obvious additive effect was found among favored alleles affecting CT. Several favored alleles showed pleiotropic genetic effects, such as decreasing CT, increasing chlorophyll content and kernel weight.
小麦生育期的温度, 尤其是开花以后的温度变化对植株衰老、粒重和品质等都有很大影响。由于植株对其自身温度有一定的调节作用, 因此气温与植株的冠层温度有一定差异。这种差异不仅与植株的生长状况有关, 而且与小麦的基因型密切关联。在相同背景条件下, 不同小麦基因型冠层温度存在的差异被称为冠层温度分异特性。在冬小麦上已对冠层温度的分异特性进行了较深入的研究, 不同基因型小麦材料在同一气候、土壤和栽培条件下, 冠层温度存在差异。张嵩午等[1, 2]依据灌浆期间冠层温度高低, 将小麦品种分为冷型、暖型和中间型3种类型。
伴随着光学技术、半导体技术、电子技术的不断发展, 红外测温技术也形成了完整的理论, 并广泛地应用在农业、科研、生态学等领域。随着这项技术的发展, 相应仪器的性能也不断得到改进。目前所使用的仪器一般都在1 kg以内, 为手持式、数字液晶显示、电池驱动, 携带和使用都很方便, 尤其适合于研究植物群落的冠层温度。由于冠层温度的测量具有快速、简单的特点, 因此, 很多研究者对冬小麦花后期冠层温度动态变化与产量性状之间的关系进行了研究, 发现冠层温度作为高产指标的条件取决于品种生长的环境。在灌溉条件下, 春小麦的冠层温度与产量及其构成因子间显著[3]负相关[4, 5]。Reynolds等[6]指出, 冠层温度主要与春小麦的产量和穗数显著相关, 并提出可应用冠层温度进行高产潜力品种的选择。大量研究表明, 灌浆期间的冠层温度可有效地反映小麦产量的遗传类型, 其中开花后2周内的冠层温度更能反映不同基因型春小麦的产量潜力[7, 8]。Garrity和O’ Toole[9, 10]测定了28个水稻基因型的冠层温度, 发现水分胁迫后不同品种间冠层温度具极显著差异, 严重胁迫下冠层温度高的品种在充分灌溉条件下其冠层温度也较高; 营养生长期抗旱性突出的品种扬花期冠层温度较低。Feng等[11]的研究也显示, 较低冠层温度的冷型小麦在干旱胁迫条件下, 叶功能期、叶绿素含量、超氧化物歧化酶活性、蛋白质含量、呼吸速率、以及净光合速率等生理性状都优于冠层温度较高的暖型小麦。
冠层温度已成为作物抗旱基因型选择的重要依据。春小麦产量与灌浆过程中的冠层温度呈显著负相关, 且随着灌浆推移, 相关性增大, 并可应用冠层温度选择高产潜力品种。利用冠层温度预测冬小麦产量潜力的最佳时期是育种家最关注的目标, 但前人报道相对较少且研究结果缺乏一致性。因此, 本研究利用遗传背景基本相似的选择导入系, 以消除抽穗、灌浆时期差异过大带来的误差, 找到冠层温度测定的最佳时期, 并与分子标记相结合, 揭示其遗传的基本规律。
以郑麦366、周麦18、邯郸6172和偃展4110作母本、以中国普通小麦微核心种质[12]中的60余份材料作父本配制杂交组合, 再以这4份材料为轮回亲本回交2~3代, 自交4~5代, 通过表型选择, 形成4个亲本的优异性状的选择导入系群体, 分别定名为ZM366-ILs (70份)、ZM18-ILs (136份)、HD6172- ILs (59份)和YZ4110-ILs (24份), 共计289个品系。于2012年和2013年秋, 在中国农业科学院河南新乡试验基地按小区种植所有品系, 2次重复。小区面积8.0 m × 1.2 m = 9.6 m2, 行距25 cm; 小区播种量为0.25 kg, 均采用Wintersteriger Plotseed TC (奥地利)播种。
1.2.1 冠层温度 自开花期开始, 选择晴朗无风少云的天气, 于13:00至15:00, 采用雷泰ST20型(美国)手持式红外测温仪测定冠层温度, 每7 d测定一次。光谱通带为8~14 m, 其灰度值为0.95。为消除太阳方位角和种植方向对观测值的影响, 测定时仪器探棒沿小区种植走向, 在高于小麦冠层20 cm与冠层呈30° 夹角处测量长势均匀的部位, 避免红外线照射裸露地面。每小区测3次, 取平均值作为该次测定的冠层温度值。
1.2.2 旗叶叶绿素含量 在测定冠层温度的同时, 采用SPAD-502叶绿素仪(日本)测定289份导入系的旗叶叶绿素含量(相对值), 选取旗叶中部(避开叶脉)测量, 测量面积为2 mm × 3 mm, 厚度不超过1.2 mm, 深度滑块(depth stop)可以使被测叶片放入的深度保持一致。每份材料测10次, 取平均值。
1.2.3 产量及其相关性状 成熟期调查单位面积穗数、株高, 每个小区随机取20个单株考察穗粒数。用Wintersteriger小区收割机(奥地利)分小区收获计产, 折合成公顷产量。从每小区随机取500粒称量, 重复4次, 计算千粒重。
选取与穗粒数和千粒重平均值显著关联的44个SSR位点[13, 14], 参考Wang等[13]描述的方法进行PCR扩增及基因型数据读取。
为了分析材料间遗传协方差程度, 用SPAGeDi软件[15]分析基因型数据, 构建材料间亲缘关系矩阵。根据Loiselle等[16]提出的方法, 采用10 000次排列测验来计算材料间的共祖系数。如果2个材料间的值为0, 说明这2份材料无关[17]。为了降低关联分析的假阳性比率, 首先采用STRUCTURE3.0软件[18, 19]分析所有材料的群体结构。选取分布于小麦21条染色体上的42对不连锁的SSR标记, 估计全部材料的群体结构。参数burn-in period为50 000次迭代次数, Markov Chain Monte Carlo (MCMC)为500 000次重复[13]。
采用TASSEL 2.1软件中同时考虑群体结构和亲缘关系(Q+K)的混合线性模型(MLM), 对选择导入系进行组关联分析, 并计算优异等位变异的相对贡献率[13, 14]。
冬小麦冠层温度在基因型间存在显著差异, 同一材料重复间差异显著, 说明冠层温度受基因型和生长环境共同影响, 并且两者存在一定程度的互作(表1)。
| 表1 冠层温度的方差分析 Table 1 Analysis of variance on canopy temperature (CT) |
4套选择导入系的冠层温度存在一定差异, 其中ZM18-ILs的冠层温度无论是在灌浆前期、还是中后期都显著低于其他3个导入系; YZ4110-ILs的冠层温度在灌浆前期及后期显著高于其余各系; ZM366-ILs和HD6172-ILs的冠层温度介于ZM18- ILs和YZ4110-ILs之间(表2)。同样, 4套选择导入系的叶绿素含量也存在差异, HD6172-ILs的叶绿素含量在灌浆前期显著低于其余各系; Z18-ILs的叶绿素含量较高, 尤其是在灌浆后期, 显著高于其余各系。比较产量及其构成因子, YZ4110-ILs的穗粒数显著低于其余各系; Z18-ILs的千粒重显著高于其余各系, 由于千粒重对产量具有重要作用, 因此, 使得Z18-ILs的单产显著高于其余各系(表2)。
4个时期的冠层温度与产量都呈负相关(表3)。灌浆初期, 冠层温度与产量呈不显著的负相关, 随后呈显著或极显著负相关, 并且相关系数随灌浆进程逐渐增大。冠层温度与千粒重也呈负相关关系, 并且变化趋势同冠层温与产量的关系, 但是冠层温度与千粒重的相关程度更高。与此相反, 冠层温度与单株有效穗数表现正相关, 灌浆初期相关性不显著, 随着时间的推移, 两者间的相关系数逐渐增大, 并达到显著或极显著水平。冠层温度与穗粒数之间相关不显著。
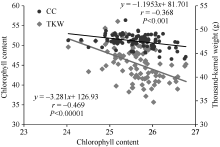
对4个时期所测得的冠层温度和叶绿素取平均值后, 进一步分析了冠层温度、叶绿素含量和千粒重三者间的关系, 回归分析显示, 冠层温度与叶绿素含量及千粒重均呈极显著负相关。即品种的冠层温度越低, 该品种的叶绿素含量则相对较高, 最终品种的千粒重也较高(图1)。说明较低的冠层温度对叶绿素及光合系统起着保护作用, 从而保证后期的光合作用, 保障灌浆中后期籽粒的充实。
| 表2 4套选择导入系(ZM366-Ils、ZM18-Ils、HD6172-ILs和YZ4110-ILs)间表型变异的比较 Table 2 Phenotypic comparison among SIL lines in genetic background of Zhenmai 366 (ZM366-ILs), Zhoumai 18 (ZM18-ILs), Handan 6172 (HD6172-ILs), and Yanzhan 4110 (YZ4110-ILs) |
| 表3 冠层温度与产量及构成因子间的关系 Table 3 Correlation coefficients between canopy temperature (CT) and yield and its component factors |
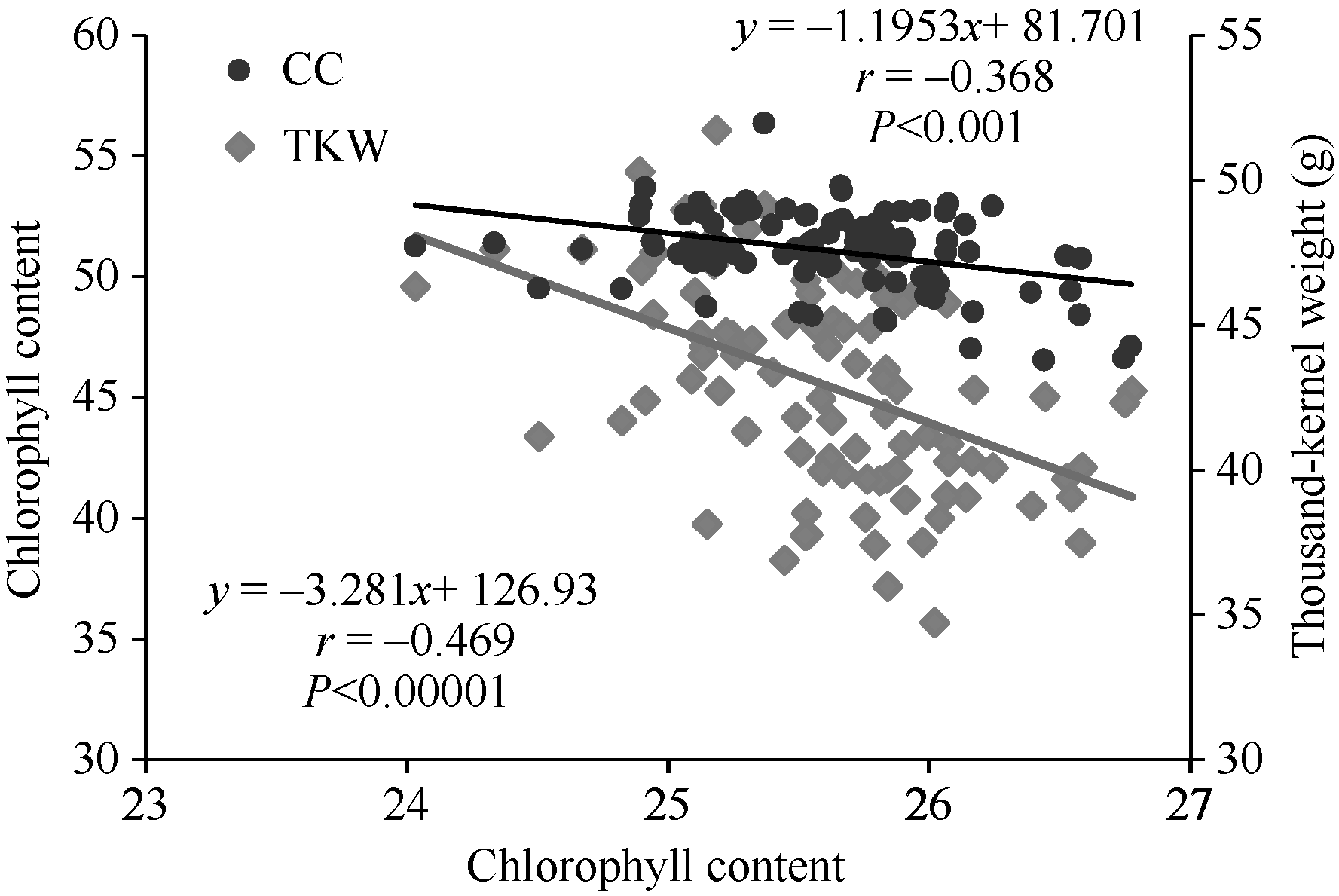 | 图1 冠层温度与叶绿素含量(CC)、千粒重(TKW)间的关系(2013)Fig. 1 Correlations between chlorophyll content (CC), thousand-kernel weight (TKW), and canopy temperature (CT) in 2013 |
利用44个与穗粒数或千粒重相关联的SSR位点对全套材料扫描, 共检测到9个SSR位点与冠层温度显著关联(P< 0.01)。其中, 有4个位点在3个时期都能被检测到关联, 占关联位点的44.5%; 有3个位点在2个时期被检测到关联, 占关联位点的33.3%; 另2个位点仅在1个时期被检测到关联, 占关联位点的22.2%。这9个位点中, 5个位点同时与穗粒数和千粒重相关联, 另2个位点与千粒重显著关联, 占关联位点的77.8% (表4)。在9个关联位点中, 6个位点上检测到优异等位变异, 携带优异等位变异材料的冠层温度显著低于携带非优异等位变异材料的冠层温度(图2)。
 | 图2 关联位点上优异等位变异的表型效应 * 表示携带优异等位变异材料的冠层温度显著低于携带非优等位变异材料的冠层温度。Fig. 2 Phenotypic effect of favored alleles at loci associated with canopy temperature * Canopy temperatures of accessions contained favored alleles were significantly lower than those contained unfavored alleles. |
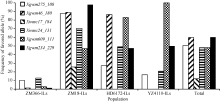
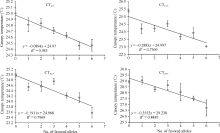
优异等位变异在导入系中的分布, 以Xgwm 234_229和Xgwm46_180的频率最高, 其次是Xgwm 275_108、Xgwm609_111和Xwmc24_131, Xwmc17_ 184的频率较低(图3)。4个导入系中, 以ZM8-ILs包含的优异等位变异数量最多, 并且各个优异等位变异在该选择导入系中的频率也都较高; HD6172- ILs包含的优异等位变异数量其次; YZ4110-ILs再次; 而ZM366-ILs携带的优异等位变异数最少, 且各个优异等位变异在该导入系中的频率也较低(图3)。同时发现, 在4个检测时期, 随着导入系材料包含优异等位变异数量的增加, 其冠层温度呈降低趋势(图4)。
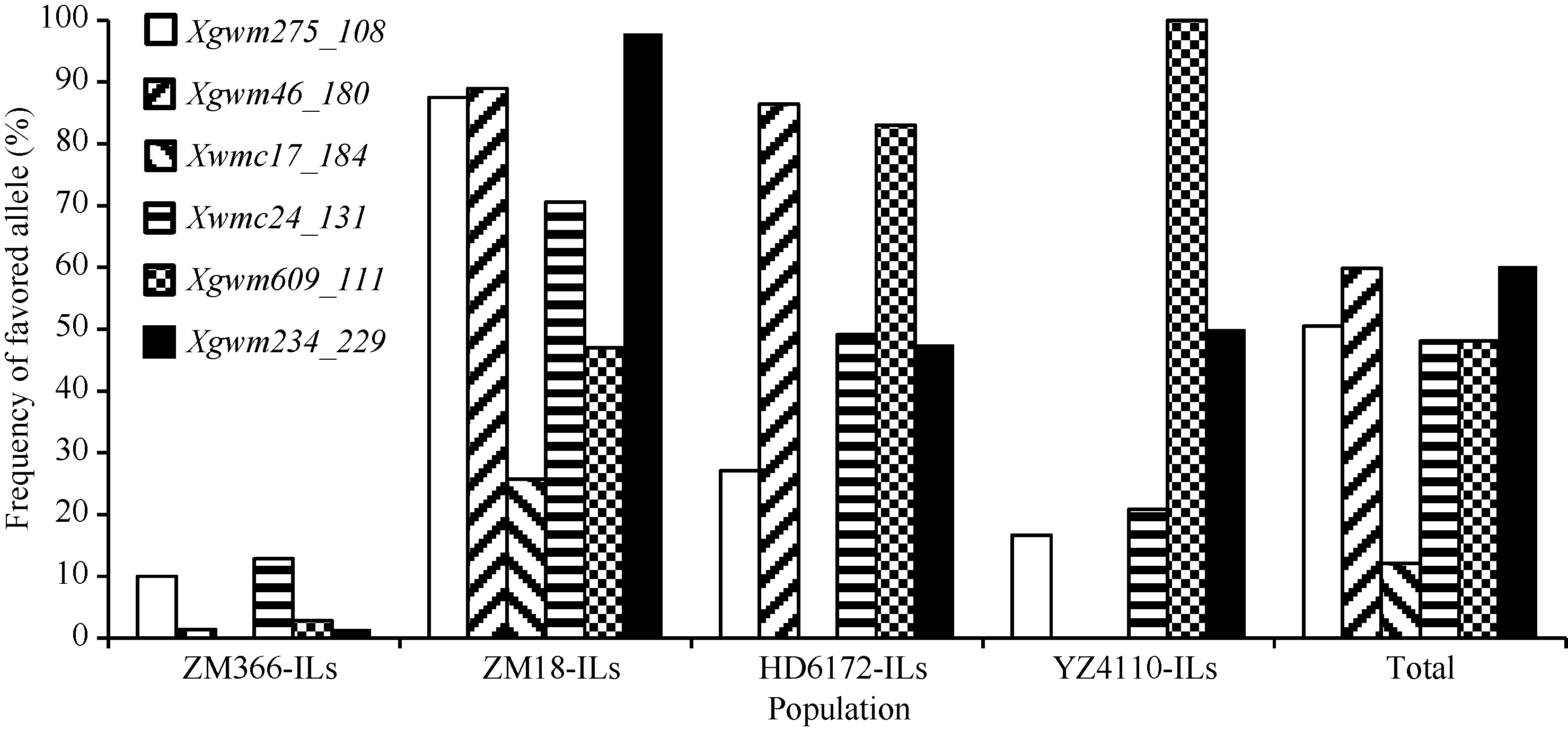 | 图3 优异等位变异在选择导入系中的频率分布Fig. 3 Distribution of favored alleles in selected introgression lines |
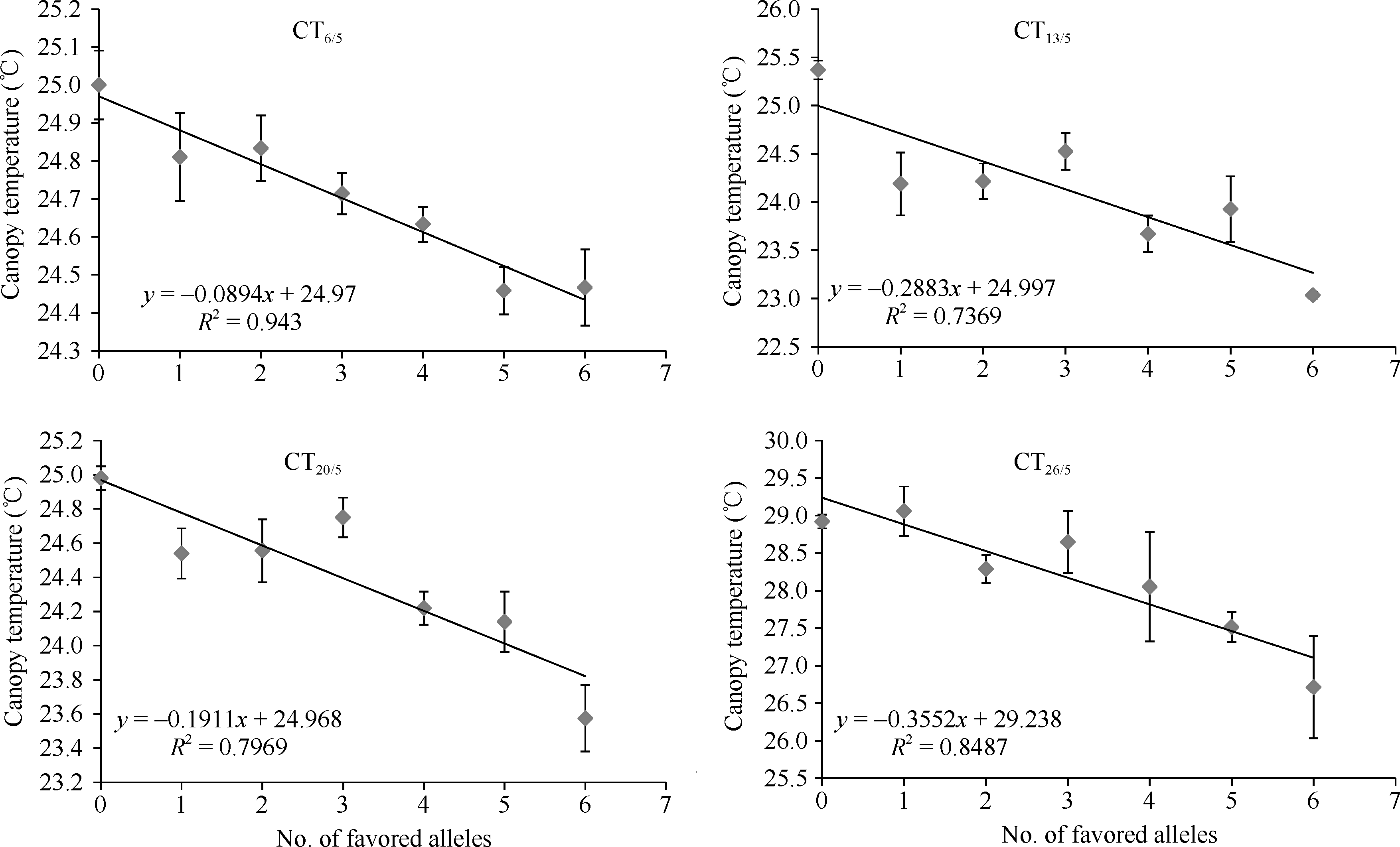 | 图4 冠层温度与优异等位变异数之间的关系Fig. 4 Correlations between canopy temperature and number of favored alleles at different stage |
4套导入系的叶绿素含量存在明显差异, 尤其是后期的叶绿素含量(表2)。用上述44个与穗粒数或千粒重相关的SSR位点与叶绿素含量的关联分析发现, 20个位点与其呈现不同程度的关联(表5)。非常有趣的是与冠层温度关联的9个位点均在其中, 并多与灌浆后期的叶绿素含量关联(表4和表5)。
| 表4 与不同灌浆时期冠层温度显著关联的SSR位点 Table 4 Loci associated with canopy temperatures at different grain filling stages |
| 表5 与不同灌浆时期叶绿素含量显著关联的SSR位点 Table 5 Loci associated with chlorophyll content at different grain filling stages |
冠层温度的测定具有快速、简单的特点, 已成为高通量表型鉴定的一个重要性状指标。很多研究者对冬小麦籽粒花后期冠层温度动态变化与产量性状之间的关系进行了研究, 发现冠层温度作为高产指标的条件取决于品种生长的环境[3]。在灌溉条件下, 春小麦的冠层温度与产量及其构成因子间呈显著负相关[4, 5]; 冠层温度主要与春小麦的产量和穗数呈显著相关, 并提出可应用冠层温度进行高产潜力品种的选择[6]。大量研究表明, 灌浆期间的冠层温度可有效地反映小麦产量的遗传类型, 其中开花后两周内的冠层温度更能反映不同基因型春小麦的产量潜力[7, 8]。李向阳等[20]对小麦冠温和产量的关系探讨发现, 在河南省的生态条件下, 小麦灌浆期间的冠层温度与小麦产量及其主要构成因素大部分呈负相关。徐银萍等[21]报道, 冬小麦产量与灌浆过程中的冠层温度呈显著负相关, 且随着灌浆进程, 相关性增大。由此可见, 冠层温度可以作为产量潜力的间接选择指标[22]。多数研究认为, 不同基因型小麦的冠气温差和产量显著相关[23, 24], 如李向阳等[25]研究发现, 无论是强势粒还是弱势粒的灌浆速率, 冷尾小麦豫麦50均大于暖尾小麦豫麦34和豫麦70。然而, 也有研究发现, 在水分胁迫条件下, 当土壤含水量是小麦产量的主要限制因子时, 冠层温度与产量相关不显著[26, 27]。本研究利用289份选择导入系, 连续两年度考察自开花至成熟4个时期的冠层温度与产量及其构成因子间的关系, 发现冠层温度与产量、千粒重、穗粒数和单株有效穗数显著相关, 且随着灌浆进程, 相关性逐渐增大, 获得与前人研究一致的结论: 高产基因型无论是在水分充足的条件下, 还是在水分胁迫条件下, 均表现较低冠层温度, 小麦冠层温度因基因型而异, 尤其是在小麦抽穗至成熟期间更为明显。本课题组多年系统观察表明, 不同类型小麦冠层的高、低温特性相当稳定, 不因年份的更替和天气的变化而变化, 因此认为, 冠层温度可以作为优异种质筛选的简易指标, 本研究结果进一步表明, 灌浆中后期或末期的冠层温度更适合作为优异种质筛选的指标之一。
本研究结果显示, 叶绿素含量与千粒重和产量显著正相关, 随着灌浆进程, 两者间的相关性逐渐增大, 高产(超高产)品种在生育后期都具有较高的叶绿素含量和光合速率、以及较大的光合叶面积指数和较高的PSII最大光化学效率, 并且旗叶可溶性糖和可溶性蛋白含量不仅高而且下降缓慢, 使得叶片的光合功能期延长[28, 29]。李永攀等[30]研究表明, 持绿型小麦品种抗衰老, 光合作用时间长, 籽粒充实, 千粒重提高。本研究中周麦18选择导入系群体的产量显著高于其他3套导入系, 其在在灌浆末期, 表现出显著低的冠层温度和显著高的叶绿素含量, 表明在灌浆期能维持较高的光合生产能力, 通过热量耗散的增加来保护叶片不受强光的伤害, 就能延迟叶片的衰老, 使叶片维持较高的叶面积系数和叶绿素含量。
如前所述, 与冠层温度关联的位点均与灌浆中后期旗叶的叶绿素含量明显相关, 且对两性状而言, 优异等位变异存在高度的一致性, 它们又是与高千粒重关联的优异等位变异[13, 14], 说明灌浆后期, 良好的根系功能保证了水分和营养的正常供应, 降低了冠层温度, 对叶绿素和光合起到了良好的保护作用, 最终体现在千粒重和产量的增加。我们对4个轮回亲本(周麦18、偃展4110、郑麦366和邯郸6172)根系的观察证明了上述推测。因此灌浆中后期冠层温度可作为小麦根系大小、活力及产量潜力的重要选择指标应用于育种。
尽管小麦冠层温度无论在灌溉条件下, 还是在逆境胁迫条件下, 都显示出与产量存在显著关系, 但关于冠层温度的遗传机制研究相对较少。Reynolds等[31]认为冠层温度是小麦基因型在特定环境下的遗传表现, 主要受光合有效辐射、气孔导度、空气温度和VPD等一系列生理生态因子的影响。Lopes等[32]选取288份来自于世界各地的春小麦种质, 将其分别种植于北非、西亚、南亚等12个环境中, 发现基因型与环境间存在明显互作; 性状间的关系也与环境密切相关; 冠层温度表现为中等遗传力, 且在所有环境中均表现与产量负相关。申国安等[33]于1997— 1999年用6个不同的材料配制成完全双列杂交, 研究表明, 小麦冠层温度有较高的遗传力, 比较稳定, 小麦冠层温度的遗传符合加性-显性遗传模型, 以加性效应为主, 呈部分显性, 低温型为显性或部分显性。Saint等[34]也认为小麦冠层温度的遗传以加性效应为主, 但同时也存在显性和共显性的遗传模型, 且基因型与环境间存在明显互作。Rebetzke等[35]研究显示, 冠层温度的遗传力约为0.75, 在3种环境中共检测到16个影响冠层温度的QTL, 其中部分QTL与控制株高和植株形态发育等的QTL相重叠。本研究以高世代选择导入系群体为试验材料, 利用关联分析法一次性检测到比前人更多的影响冠层温度的位点, 与前人已定位到的QTL相比, 一部分定位结果相一致或位于其区间附近, 此外, 还检测到新的QTL。并且本研究所检测到的关联位点中, 有近一半在整个小麦灌浆期都能被检测到关联, 因此, 我们推断这些位点可能对叶片的蒸腾作用及叶片的衰老起重要作用。同时, 这9个位点全部与穗粒数和千粒重同时显著关联[13, 14]。另外, 这9个与冠层温度显著关联的位点中, 有8个还与叶绿素含量同时显著关联, 推测一因多效或基因间的紧密连锁可能是同一个标记位点与多个性状相关联或性状相关的遗传基础。
冠层温度与产量和千粒重显著负相关, 而与有效穗数显著正相关, 并随灌浆进程, 相关性逐渐增强。发掘出9个与冠层温度显著关联的位点, 并在6个位点上检测到对育种有显著正效应的优异等位变异。这些优异等位变异间存在明显的加性效应, 同时这些位点还与千粒重和穗粒数也呈现显著关联。
致谢: 中国农业科学院作物科学研究所科研助理付丽娜、田维、宋丽宁参与了冠层温度及田间农艺性状数据的调查和收集, 在此一并致谢。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|