



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
长江中下游小麦品种根系改良特征及其与产量的关系
[田中伟, 樊永惠, 殷美, 王方瑞, 蔡剑, 姜东, 戴廷波*  ]
]
]
|
|


为探明小麦品种更替过程中根系改良特征对氮肥的响应及其与产量形成的关系, 从而为高产品种选育及根冠构型建成提供理论依据。本研究以长江中下游不同年代大面积推广的代表性小麦品种南大2419、扬麦1号、扬麦158和扬麦16为材料, 采用大田和盆栽试验, 研究了3个施氮水平(纯氮0、225和300 kg hm-2)下小麦根系形态与生理特性的差异及其与产量的关系。结果表明, 小麦籽粒产量随品种育成年代推进逐步增加, 现代品种对施氮的响应较早期品种大。现代小麦品种拔节至开花阶段根系干物质积累量和生长速率显著高于早期品种, 而播种至拔节期早期品种的根系生长在无氮条件下具有较强的生长优势。小麦根系总根长、表面积、根体积、0~60 cm土层根重密度、根系活力和SOD活性随品种育成年代逐步提高, 而MDA含量显著降低。增施氮肥促进了不同年代品种根系生长, 但现代品种增幅较早期品种大, 说明品种改良提高了小麦根系对氮肥的响应。籽粒产量与开花期根系总根长、表面积、根系生物量和0~60 cm根重密度呈显著正相关。因此, 增加根系与土壤接触面积和高氮适应性、提高根系生理活性、延缓根系衰老是长江中下游小麦品种演变的重要特征也是高产高效栽培调控的重要目标。
Exploring the characteristics of root growth, root physiology and their relationships to grain yield in wheat cultivars released in different decades will facilitate the development of new wheat cultivars, with stable and high yields and low nitrogen fertilizer input. Four wheat cultivars Nanda 2419, Yangmai 1, Yangmai 158, and Yangmai 16, which were bred or widely planted in the Middle-Lower Yangtze River Basin after 1950, were used in field and pot experiments in 2010-2011 with three nitrogen rates (pure N 0, 225, and 300 kg ha-1). The results showed that, grain yield was improved with the genetic improvement in wheat breeding and the sensitivity of modern cultivars to N was greater as compared with early released cultivars. The root dry matter accumulation (DMA) and growth rate from jointing to anthesis of modern cultivars were higher than these of early released cultivars. The root length, root surface area, root volume and density of root weight in 0-60 cm soil layer were enhanced with the genetic improvement in wheat breeding, while no change was found in proportion of root distribution. The root activity, MDA concentration and SOD activity in modern cultivars were improved significantly. Higher N application increased root growth and physiological performance, with larger effects in modern cultivars, indicating that the response to N and high-N tolerance were promoted with genetic improvement in wheat breeding. In addition, these were significantly positive relations of grain yield with total root length, root surface area, root biomass and density of root weight in 0-60 cm soil layer. Therefore, higher contact area of the roots with soil and high-N adaptability, enhanced root physiological activity and N assimilation capacity, and delayed senescence of root system should be the important characteristics in cultivar evolution in the Middle-Lower Yangtze River, which are the main purpose getting in high-yield and high efficiency cultivation in this region.
小麦是世界上重要的粮食作物, 提高小麦单产潜力是保障我国乃至世界口粮安全的重要途径。品种改良是小麦产量提高的重要推动力, 20世纪50年代以来我国主要冬麦区共经历5~6次大的品种变革, 每次品种更替均大幅度提高了小麦产量潜力。前人关于小麦品种改良过程中农艺性状、光合生产、干物质积累转运和生理特性的演变规律及其与产量的关系已有较深入的研究。多数研究认为, 小麦品种更替中产量提高主要由于叶片增大、光合能力增强、生理代谢旺盛、物质生产能力提高进而引起单穗产量和收获指数增加[1, 2, 3, 4]。我国近60年的育种和生产实践表明, 以往育种家在选育品种过程中多以地上部综合性状作为选育高产品种的主要依据。由于小麦根系生长在地下, 其形态和生理特性观测难度较大, 因此一直被国内外育种家所忽略[5]。但小麦根系作为水分和养分的主要吸收器官和多种激素和氨基酸合成的重要场所, 其形态和生理特征与地上部的生长发育、产量和品质形成及养分的高效吸收利用均有密切的关系[6]。
近年来, 小麦根系已成为小麦高产、高效、优质栽培的一个较为活跃的研究领域。前人对小麦根系形态与分布特征[7]、根系发育建成规律[8]、根系衰老[9]、根冠平衡[10]、根系活力[11]、根系吸收动力学[12]、根系分泌特性[13]等都有较深入的研究, Mailk等[14]通过研究19个不同小麦品种根系性状与产量的关系发现, 小麦根系生长存在显著的基因型差异, 籽粒产量高的品种根系干重较大, 但根系氮含量较低。Chochois等[5]研究发现禾本科短柄草(Brachypodium)的根系构型和根系生长特征与小麦根系高度相似, 可通过分子生物学手段将短柄草目的基因转入小麦中从而改善小麦根系构型、提高小麦产量。但纵观国内外研究, 前人多侧重于不同栽培条件下根系形态生理的变化, 而小麦品种更替过程中根系生长、分布和根系生理特性的演变规律及其与产量的关系至今并不明确。探明小麦品种改良过程中与产量形成密切相关的根、冠、生理特征的变化及氮肥效应, 对选育高产高效品种和制定未来小麦高产育种目标均具有重要意义。因此, 本研究选用长江中下游不同年代主推品种通过大田和盆栽试验, 分析不同施氮条件下不同年代小麦品种根系生长、分布和根系衰老特性的差异, 以期明确小麦品种改良过程中根系生长与生理特性的演变规律及氮肥效应, 为小麦高产高效品种选育和栽培管理提供理论依据。
2010— 2011年度在南京农业大学江浦和牌楼试验站分别进行大田和盆栽试验, 均采用20世纪50年代以来长江流域育成的代表性品种, 包括南大2419 (1950年后在长江流域推广, 最大面积500万公顷)、扬麦1号(1967年审定, 上世纪70年代大面积推广)、扬麦158 (1993年审定, 累计推广面积近1亿公顷)和扬麦16 (2004年审定, 目前长江中下游麦区推广面积最大的品种)。均设全生育期施纯氮0 (N0)、225 (N225)和300 kg hm-2(N300) 3个氮肥处理。50%氮肥和全部磷(P2O5 150 kg hm-2)、钾肥(K2O 150 kg hm-2)作基肥, 另外50%氮肥拔节期追施。
大田试验播种前0~25 cm土层土壤含有机质21.62 g kg-1、全氮1.12 g kg-1、速效氮14.39 mg kg-1、速效磷17.40 mg kg-1、速效钾115.52 mg kg-1。试验小区面积为3 m × 3 m = 9 m2, 行距25 cm, 随机区组排列, 3次重复。11月1日播种, 三叶期定苗, 基本苗1.8 × 106 hm-2。采用20 cm × 20 cm孔径的防倒网防止倒伏, 其余管理措施(病虫草害防治等)同大田高产栽培。
盆栽试验用土取自高产田耕层土壤, 含有机质10.95 g kg-1、全氮0.79 g kg-1、速效氮26 mg kg-1、速效磷25.6 mg kg-1、速效钾72.3 mg kg-1。自然风干过筛后装盆, 播种前与肥料充分混匀, 装入直径30 cm、高40 cm的聚乙烯塑料桶, 每桶装风干土12.0 kg。11月20日播种, 三叶期定苗, 每盆留苗10株, 随机区组排列, 3次重复。
因育成年代不同, 供试品种抽穗期有差异, 其中扬麦158与扬麦16相当, 南大2419和扬麦1号分别较扬麦16晚4 d和2~3 d。
1.2.1 根系形态 分别于拔节期、孕穗期、开花期和成熟期取盆栽小麦根系, 每重复取样1盆(10株), 在清水中反复洗净后, 应用根系分析仪(WinRhizo LA 1600型)扫描完整根系, 记录总根长、表面积、根体积等形态数据。将扫描后根系于105℃烘箱杀青30 min, 80℃烘干至恒重, 称重获取根系生物量。根系生长速率(g d-1) = 根系生物量/生长天数。
1.2.2 根系时空分布 采用冯福学等[7]描述的方法, 于小麦孕穗期和开花期, 使用钻头直径为4 cm, 长100 cm的根钻, 每处理取3钻, 钻的中心分别在种植行上、行间距1/2处和行边缘(钻与种植行相切), 3钻混合作为一个样品, 并按土层分样, 每20 cm为一个层次。将根样放于烘箱内105℃杀青30 min, 然后过100目筛, 用镊子去除杂质和杂根, 用自来水冲洗干净; 再将根样品在80℃烘箱内烘干, 用万分之一天平称重。根干重密度(× 10-4 g cm-3) = 10 000 × 根干重/土体体积; 土体体积V= π r2h, 式中r为钻头半径(2 cm), h为取样深度(20 cm)。
1.2.3 根系活力和抗氧化特性 在拔节期、孕穗期和开花期取盆栽小麦根系, 每重复取1盆, 将根系在清水中洗净后, 从各处理取相同部位(根尖1 cm处)迅速测定根系活力。同时, 取根系样品于液氮中速冻保存, 用于丙二醛(MDA)含量和超氧化物歧化酶(SOD)活性的测定。采用三苯基氯化四氮唑(TTC)还原法[15]测定根系活力, 按Tan等[16]描述的方法测定根系MDA含量和SOD活性。
于大田试验成熟期取1 m2样段, 人工收割, 人工脱粒, 自然晒干后称重, 实收计产。于盆栽试验成熟期从每处理随机收获3盆, 人工脱粒, 自然晒干后称重计产。
使用Microsoft Excel 2007处理数据, 采用SPSS17.0 (SPSS Inc., Chicago, IL, USA)软件进行方差分析(ANOVA)和显著性测验(Duncan’ s), 用SigmaPlot10.0软件绘图。
不同年代小麦品种不同生育阶段根系干物质积累存在显著的基因型差异, 生长速率在拔节至开花阶段达到最大值(表1)。在播种至拔节阶段, 在N0处理条件下, 品种间的干物质积累和生长速率均以早期品种最大, 且随年代进程显著降低, 在N225和N300处理下, 现代品种的积累量和生长速率明显高于早期品种; 在拔节至开花阶段, 现代品种的干物质积累和生长速率显著大于早期品种。不同年代小麦孕穗期和开花期根冠比均存在显著的基因型差异, 施氮显著降低了小麦的根冠比。早期品种孕穗期和开花期根冠比在N0处理下大于现代品种, 而在N300处理小于现代品种。说明早期品种在低氮条件下根系具有较强的生长优势, 而高氮条件更适宜现代品种根系生长。
| 表1 盆栽试验3种施氮水平下不同年代小麦品种单株根系生物量(RB)、生长速率(RGR)和根冠比(RSR) Table 1 Root biomass (RB), growth rate (RGR), root-shoot ratio (RSR) of a single plant under three nitrogen levels in wheat cultivars released in different years in pot experiment |
随着生育时期的推进, 小麦根系总根长、表面积和根体积均显著增大, 且现代小麦品种显著大于早期品种, 品种间差异在开花期达到最大, 在不同氮肥处理下均表现为扬麦16> 扬麦158> 扬麦1号> 南大2419 (表2)。说明品种改良提高了小麦根系与土壤的接触面积, 增加了对水分、养分的吸收面积, 为提高产量奠定了基础。施氮显著提高了不同年代小麦品种根系总根长、表面积和根体积, 但不同年代品种在不同生育时期对氮肥的响应不同。拔节期早期品种南大2419根系总根长、表面积和根体积随施氮量的增加呈先升高后降低的趋势, 而现代品种随施氮量的增加而增加; 在开花期早期品种均在N225处理达到最大值, 而扬麦16在N300处理最大, 说明品种对氮肥的响应和对高氮的耐性随品种改良逐步增强。
| 表2 盆栽试验3种氮肥水平下不同年代小麦品种单株总根长(RL)、根表面积(RSA)和根体积(RV) Table 2 Root length (RL), surface area (RSA), and volume (RV) of a single plant under three nitrogen levels in wheat cultivars released in different years in pot experiment |
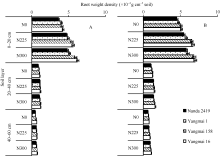
不同年代小麦品种根系时空分布存在显著的基因型差异, 各土层根重密度随着生育时期推进显著增加(图1)。品种改良显著提高了小麦孕穗期和开花期0~60 cm各土层根重密度, 降低了孕穗期和开花期N0处理下0~20 cm土层根重比例, 提高了N225处理下0~20 cm土层根重比例, 但差异未达到显著水平。说明品种根系改良主要是提高了各土层根系干重, 而根系在各层次的分布比例保持相对稳定。施氮显著提高了各土层根系密度。氮肥对各生育时期、各土层根重密度调控效应均达到显著水平, 0~ 20 cm根系根重密度的品种和氮肥的互作效应达到极显著水平。说明氮肥对调控根系形态和分布具有重要的作用, 且氮肥的调控效应显著大于品种效应。
 | 图1 大田试验3种氮肥水平下不同年代小麦品种孕穗期(A)和开花期(B) 0~60 cm土层根重密度Fig. 1 Root weight density at booting (A) and anthesis (B) from 0 to 60 cm soil layer under three nitrogen levels in wheat cultivars released in different years in field experiment |
不同年代品种在不同施氮条件下随生育时期推进其根系活力变化不同, 早期品种南大2419在N0和N225处理随生育时期推进逐步提高, 在N300处理呈先降低后升高的趋势; 而现代品种扬麦16在低氮处理下随生育时期推进呈先升高后降低的趋势, 在高氮处理下逐步提高(图2)。早期品种根系活力在拔节期低氮处理下显著高于现代品种, 而在N225和N300处理下显著低于现代品种, 增施氮肥显著提高了小麦品种根系活力, 并在N225处理达到最大值, 增施氮肥对现代品种根系活力增幅显著大于早期品种。孕穗期和开花期小麦根系活力在高氮处理下均随品种育成年代推进逐步提高。进一步表明低氮条件更适宜早期品种根系生长, 而现代品种对氮肥的响应和高氮的耐性增强。
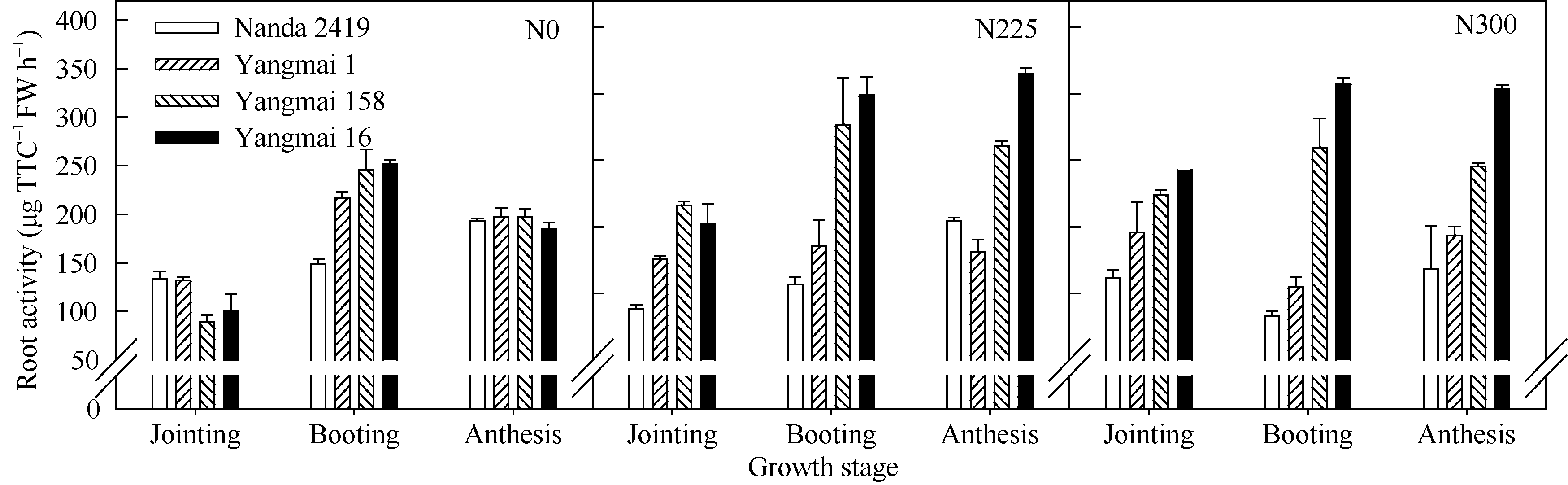 | 图2 盆栽试验3种氮肥水平下不同年代小麦品种不同生育时期根系活力的变化Fig. 2 Changes of root activity at different growth stages under three nitrogen levels in wheat cultivars released in different years in pot experiment |
小麦根系丙二醛(MDA)含量随品种育成年代推进呈逐步下降的趋势(图3)。施氮量在0~225 kg hm-2范围内, 增施氮肥显著降低了小麦根系MDA含量。小麦旗叶超氧化物歧化酶(SOD)活性随着育种年代推进呈逐步升高的趋势, 且随着施氮量的增加而提高, 但在225~300 kg hm-2范围内增施氮肥对SOD活性影响不显著。说明品种改良和适量施氮降低了根系膜酯化氧化程度, 增强了小麦清除超氧阴离子自由基(O2܋)的能力, 从而延缓了根系衰老。
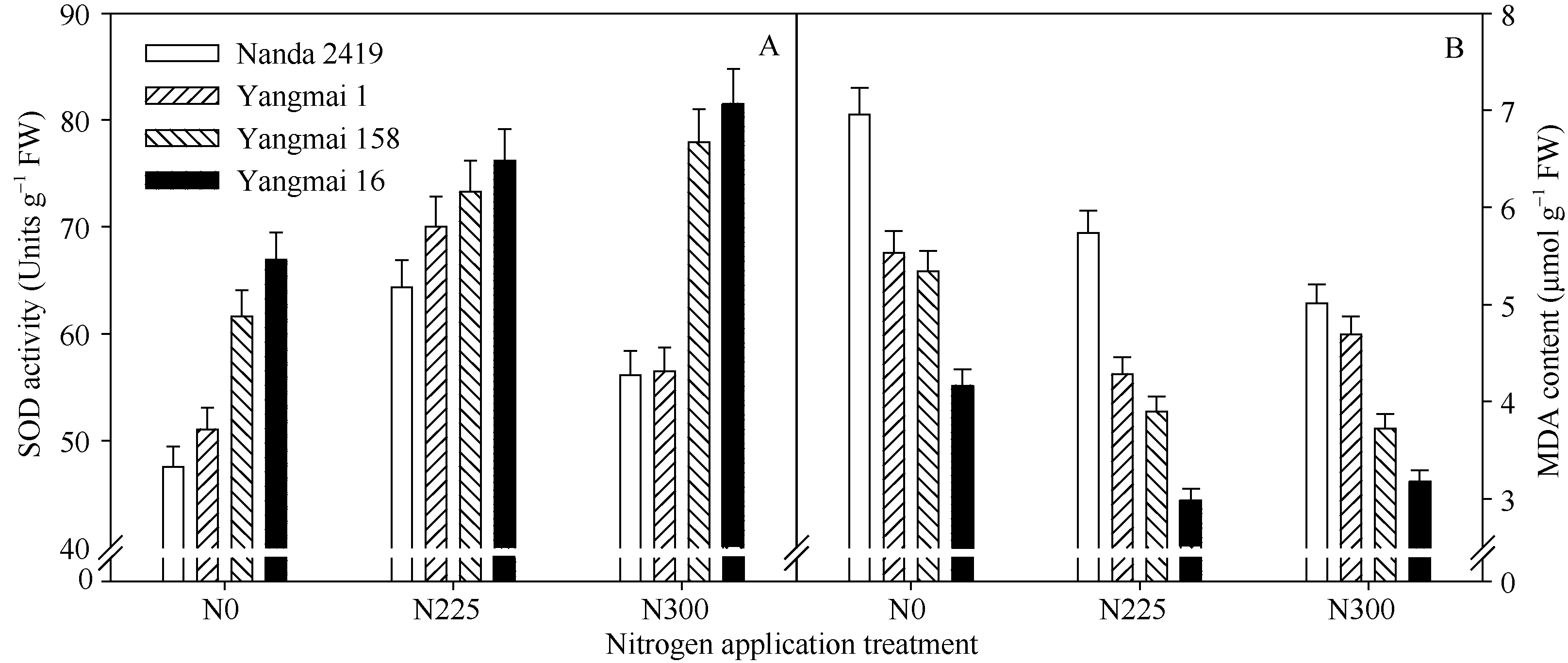 | 图3 盆栽试验不同年代小麦品种开花期根系SOD活性(A)和MDA含量(B)变化特征及其对氮肥的响应Fig. 3 Changes of root SOD activity (A) and MDA content (B) at anthesis in wheat cultivars released in different years and their response to nitrogen fertilizer in pot experiment |
小麦籽粒产量随着品种育成年代推进呈增加趋势, 在纯氮0~225 kg hm-2范围内, 施氮显著提高了不同年代小麦籽粒产量, 且氮肥对现代品种的增产效应显著大于早期品种(图4)。当施氮量超过225 kg hm-2时, 早期品种产量降低幅度显著高于现代品种。说明现代品种对氮肥的响应和对高氮的耐性增强。
 | 图4 不同年代小麦品种的籽粒产量及其对氮肥的响应 A: 大田试验; B: 盆栽试验。Fig. 4 Grain yield of wheat cultivars released in different years and their response to nitrogen fertilizer A: field experiment; B: pot experiment. |
籽粒产量与开花期根系总根长、表面积和根系生物量呈极显著正相关(图5), 说明小麦品种改良过程中籽粒产量提高与根系形态的改良密切相关。根系形态对籽粒产量的贡献表现为根系生物量> 总根长> 表面积。籽粒产量与根重密度呈显著正相关, 且下层根的分布对籽粒产量贡献更大。
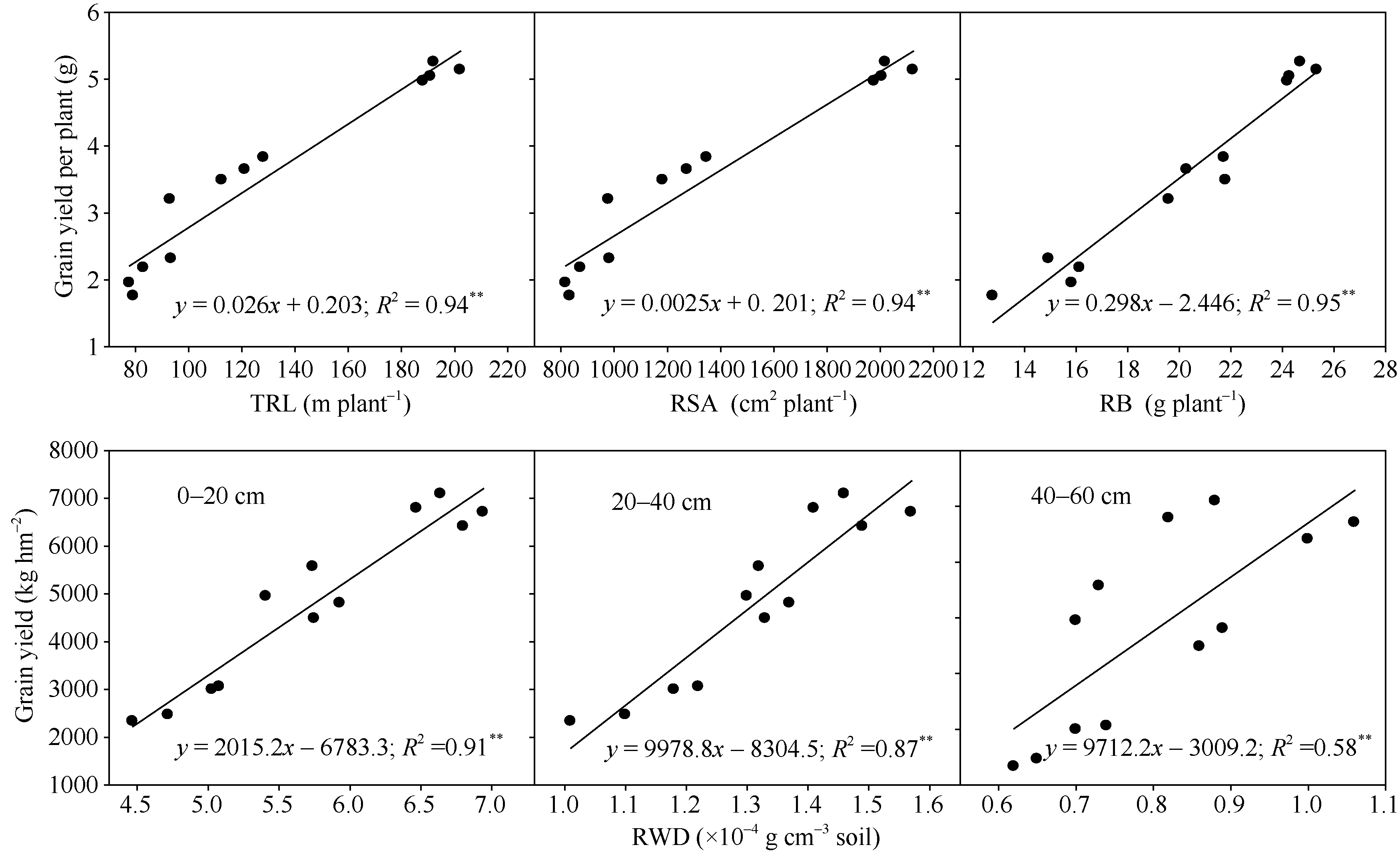 | 图5 开花期根系总根长(TRL)、表面积(RSA)、生物量(RB)及根重密度(RWD)与籽粒产量的关系Fig. 5 Relationships between grain yield and total root length (TRL), root surface area (RSA), root biomass (RB), and root weight density (RWD) at anthesis |
根系是植物重要的营养器官, 与地上部生长发育和养分的吸收利用关系密切。对根直径、干重、根长和根数等作物根系形态性状的遗传学研究发现, 这些形态性状具有较高的遗传力[17]。因此, 可通过改良作物根系性状和根系生理活性, 选育根系和地上部关系协调的品种从而提高产量。在小麦品种选育过程中, 根系改良一直是小麦育种工作的热点和难点, 其核心问题是高产小麦应该具有怎样的根系构型和生理特征。Xue等[18]研究认为, 高产品种根系发达、发根力强、根数多、扎根深, 后期根系活力高、吸收面积广、吸收能力强、衰老较晚且缓慢。但苗果园等[19]研究认为, 并非根系越发达地上部生长就越好、产量就越高, 在一定范围内, 根系的差异并不一定引起地上部和产量的差异。本研究发现长江中下游小麦品种更替中, 根系总根长、表面积和根体积显著增大, 说明品种改良提高了小麦根系与土壤的接触面积, 增加了对水分、养分的吸收面积, 为提高产量和养分吸收量奠定了基础。同时, 我们也观察到根系形态和根系生物量与籽粒产量具有较好的相关性, 且现代品种在整个生育期均显著高于早期品种。因此, 现代品种发达的根系为地上部的生长提供了充足的养分、水分和植物激素, 优化了地上部结构, 改善了地上部生长, 是品种改良过程中产量提高的重要原因。但是, 根系不仅可以吸收水分和养分, 同时也需要消耗地上部产生的光合产物以维持根系的生长。有研究表明, 用于生产单位根干物质量的能量是地上部的2倍, 根量过大会消耗大量的光合产物, 并与穗部竞争光合产物, 从而不利于高产的形成[20, 21]。因此, 在品种改良过程中构建适宜的根系群体和根冠比尤为重要。
小麦产量与水分和养分的供应及吸收利用密切相关, 根系形态是影响水分和养分吸收的重要因素, 其中根系构型在水分、养分高效利用中起着决定性作用[22]。Pang等[23]研究指出, 早期吸氮能力较强的品种一般具有较高的氮素利用效率和光合效率, 同时具有根系生长量、分布密度、有效吸收面积大等特征。Erenoglu等[24]研究发现, 根系质量、根长、总吸收表面积等与氮素吸收速率显著正相关, 且根系形态及分布对籽粒产量有直接或间接的影响。程建峰等[25]研究认为, 拔节期较高的根密度和根系吸收面积是氮高效水稻的重要特征, 可作为水稻氮素高效管理和遗传改良的可靠指标。本研究表明, 现代小麦品种根系总根长、表面积、根体积均显著高于早期品种, 说明品种改良提高了根系与土壤的接触面积, 从而为养分的高效利用奠定了基础。同时也说明增加根长、根系吸收面积是长江中下游地区品种改良的重要特征, 也可作为评价氮高效品种的重要指标。而张荣等[26]研究表明, 黄土高原半干旱地区春小麦品种在根系生长上存在冗余, 并认为以产量为目标的品种选育过程是根系减小的过程, 与本文研究结果不同。这主要与当地不同的生态条件有关, 半干旱地区水分是小麦生长的限制因素, 理想株型应是对水分竞争能力较弱的小根系品种, 小根系品种更能适应群体理想株型。因此, 应根据不同生态条件综合考虑根系的适宜大小。
叶片生长发育和光合作用所需要的无机养分大部分是由根系提供的, 根系所需的营养主要是由叶片光合作用所产生的。因此, 叶片的功能与根系生理活性是相互影响、相互制约的。我们前期研究发现, 现代品种具有较高的光合速率和叶面积指数[1], 保证了充足的碳水化合物被运送至根部, 进而促进根系更好地生长。因此, 现代品种强大的根系与地上部有着良好的互作效应, 也是现代品种获得高产的重要原因。褚光等[21]研究表明, 具有良好的根系形态和较强的根系活力是超级稻获得高产的主要原因, 也是氮高效型水稻品种的重要特征[27]。氮高效利用型小麦品种根具有较强的NO3-和NH4+的吸收能力和较强的生理代谢能力[28]。Sgherri等[29]研究认为, 通过品种改良或栽培技术手段延缓根系衰老、提高根系生理活性可有效提高小麦产量。本研究发现, 现代小麦品种根系活力较早期品种显著提高, 同时根系SOD活性显著提高, 而MDA含量显著降低, 表明品种改良提高了根系抗氧化能力, 延缓了根系衰老。因此, 通过遗传改良改善小麦根系质量, 增加根长和根直径, 促进根系深扎, 延缓根系衰老速度, 保持花后根系较强的生理活性和较大吸收面积, 是小麦品种改良和栽培调控的重要目标。
根系的分布与小麦产量和氮素吸收利用具有密切的联系, 根重密度与作物产量有较好的相关性[30]。Herrera等[31]研究表明, 增施氮肥或土壤基础肥力高均可促进小麦根系的生长发育。Gan等[32]研究指出, 施氮仅使离施肥最近区域根重增加, 且增加施氮使根重在表土层(0~20 cm)分布比例增加, 而亚表土层(20~40 cm)分布比例则降低。本研究结果显示, 现代品种根系在0~60 cm各土层的根重密度和上层根(0~20 cm)的分布比例显著增加, 且随着施氮量的增加, 根重密度和上层根的比例增加。因此, 增加根重密度和上层根比例是品种改良的重要特征, 也是评价氮高效品种的重要指标。
Passioura[33]认为, 从生存的角度考虑, 野生种较大的根冠比是必要的, 而在栽培种上, 根冠比则应适当降低。Xiong等[34]认为小麦进化过程中根冠比减小是人工选择使得物质分配流向改变的结果, 小麦地上部物质运输占据更多的优势, 这是植物对波动环境的一种生态适应。本研究结果显示, 不同年代品种根冠比随着生育时期推进逐步下降, 施氮降低了不同年代小麦品种根冠比。但在不同施氮水平下品种间差异表现不同, 在低氮条件下现代小麦根冠比较小, 而在高氮条件下早期品种相对较小。同时, 施氮显著提高了小麦籽粒产量、根系总根长、表面积、根体积和根重密度, 增强了根系的抗氧化能力, 但现代品种的增幅较早期品种大, 说明不同年代品种对氮肥的响应不同, 且随着品种改良小麦对氮肥的响应和对高氮的耐性逐步增强。品种的不同氮肥响应可能与品种的产量潜力和选育条件不同有关。早期品种产量潜力小, 少量施氮即可满足其生长需求, 增施氮肥会造成碳氮代谢失衡、产量降低; 而现代品种产量潜力大, 对肥料的需求较高, 因此对施氮的响应也较强。另外, 小麦生产中氮肥用量逐年增加, 长江中下游麦区从20世纪80年代开始大量施用化肥, 施氮量从80年代的87 kg hm-2增加到目前的230 kg hm-2左右[1]。早期品种多是在低氮条件下选育而成, 对低氮具有较强的适应性, 而现代品种选育多是在高氮环境下进行, 能够较好地适应高氮环境。而且, 随着小麦产量潜力的进一步提高需要投入更多的氮肥来保障小麦高产。因此, 选育高氮适应型品种和高氮条件下适宜根冠构型在未来品种选育中应重点考虑。
小麦籽粒产量随品种育成年代逐步提高, 现代小麦品种拔节到开花期根系干物质积累量和生长速率显著高于早期品种, 根系总根长、根系表面积、根体积、根重密度及根系生理活性随品种育成年代逐步提高, 早期品种在低氮条件下根系生长优势较强, 而现代品种对氮肥响应和对高氮的耐性增强。表明小麦品种更替过程中通过改良拔节到开花期根系生长、改善生理活性、延缓花后衰老, 从而促进了产量增加。因此, 改善小麦根系与土壤接触面积和高氮适应性、提高根系生理活性、延缓根系衰老是长江中下游小麦品种演变的重要特征, 也是小麦高产高效栽培调控的重要目标。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|