
{kind=link}
{kind=link}
{kind=link}
绿肥还田对双季稻根际土壤产甲烷古菌群落结构的影响
[赵晓萌1, *  , 刘婧娜
, 刘婧娜2, * , 易丽霞3 , 朱波3 , 代红翠2 , 胡跃高2 , 曾昭海2, * ]
, 刘婧娜, 易丽霞]
|
|


第一作者联系方式: 赵晓萌, E-mail: zgq2091010@163.com; 刘婧娜, E-mail: liujingna199066@126.com **和#表示同等贡献(** and # indicate contributed equally to this work)
利用PCR-DGGE技术及克隆文库构建方法研究, 尿素、紫云英鲜草翻压还田、黑麦草鲜草翻压还田和不施氮肥4种处理对双季稻不同生育时期(早稻季: 分蘖期, 拔节期, 成熟期; 晚稻季: 分蘖期, 扬花期, 成熟期)稻田根际土壤中产甲烷群落结构的影响。结果表明, 双季稻不同取样时期和各处理中产甲烷古菌群落结构稳定且相似, 早稻季和晚稻季的优势群落均为甲烷微菌目(Methanomicrobiales)、Rice Cluster I (RC-I)、甲烷鬃菌科(Methanosaetaceae)、甲烷杆菌属( Methanobacterium), 但早稻季产甲烷古菌群落的Shannon-Weiner指数( H)和丰富度指数( R)整体低于晚稻季。紫云英和黑麦草鲜草翻压还田处理较尿素处理更为明显地提高了双季稻(一年)稻田根际土壤中产甲烷古菌群落的Shannon-Weiner指数和丰富度指数, 但均暂未对产甲烷群落结构产生决定性影响。
, LIU Jing-Na, YI Li-Xia
With the potential to prevent nitrogen loss through nitrate leaching and resulted non-point source pollution, green manure incorporation is widely regarded as an important soil management practice. However, massive input of organic matters from green manure supplies sufficient substrate for methanogens and may change the methanogenic archaeal community structure. To investigate the variation of methanogenic archaeal community structure as influenced by green manure incorporation, we designed four treatments of urea application, Chinese milk vetch ( Astragalus sinicusL.) incorporation, ryegrass ( Lolium multiflorumLam.) incorporation and control (without nitrogen fertilizer). Rhizosphere soil at different double-rice growing periods (tillering stage, jointing stage and mature stage for early rice; tillering stage, flowering stage and mature stage for late rice) was collected. Polymerase chain reaction-denaturing gradient gel electrophoresis technology (PCR-DGGE) and clone library method were employed to analyze methanogenic archaeal community structure. Methanogenic archaeal 16S rRNA gene was amplified using the 1106F/1378R primer set. A GC-clamp was added to the forward primers, 1106F, to enable denaturing gradient gel electrophoresis analyses. Selected Electrophoresis bands were cloned with pEASY-T3 vectors and sequenced. Relative band intensity and positions were used for principal component analysis (PCA). Methanotrophic bacterial community diversity was evaluated by the Richness index ( R) and Shannon-Weiner index ( H) from DGGE band data. The obtained sequences were analyzed with DNAMAN software and manually aligned with the sequences from GenBank database using the BLAST search program on NCBI (National Center for Biotechnology Information) web site. The phylogenetic trees were constructed by 1000-fold bootstrap analysis using a neighbor-joining method with Mega 4.0. Results indicated that methanogenic community structure was relative stable with high diversity and had small variations among sampling stages and treatments. Dominant populations were Methanomicrobiales, RC-I, Methanosaetaceae and Methanobacterium over the entire double-rice season. Methanohalophilus mahii only showed in early-rice season and fewer methanogens affiliated to Crenarchaeota in late-rice season. Methanogenic community structure in mature stages of early-rice and late-rice showed slightly low similarity with that in other growth stages of double-rice, which may caused by the decrease of organic matter content in soil. Higher Shannon-Weiner index ( H) and Richness index ( R) were observed in late-rice season by comparison with those in early-rice season. This may due to the improved activity of soil microbe and methanogens caused by increased average temperature in late-rice season. Additionally, three different fertilizer treatments increased Shannon-Weiner index and Richness index of methanogenic community structure by comparison with control, especially treatments with green manure incorporation. Increasing soil organic matter and ammonium nitrogen content, development of soil reducing conditions may contribute to this result. Whereas they didn’t have a crucial impact on methanogenic community structure in this study.
甲烷(CH4)具有很强的红外线吸收潜力, 单位质量CH4的GWP (全球增温潜势)为CO2的25倍(100年尺度内), 对全球变暖的贡献率达15%[1]。稻田是大气CH4的主要排放源, 年排放量达30 Tg (20~40 Tg)[2, 3]。我国是水稻生产大国, 种植面积居世界第二, 产量居世界第一。双季稻种植区是水稻生产的主体, 种植面积占全国水稻种植面积的66%, 产量为全国水稻产量的61.3%[4]。稻田甲烷由产甲烷古菌在严格厌氧条件下产生, 土壤中有机质及植物残体、根系分泌物等作为产甲烷古菌的营养基质, 经过代谢可产生CH4和CO2。稻田中产甲烷古菌种类分布较多, 目前从水稻土壤中分离得到7个属, 分别是甲烷杆菌属(Methanobacterium)、甲烷八叠球菌属(Methanosarcina)、甲烷短杆菌(Methanobrevibacter)、甲烷囊菌属(Methanoculleus)、产甲烷古菌属(Methanogenium)、甲烷髦毛菌属(Methanosaeta)和甲烷螺菌属(Methanospirillum)[5, 6, 7]。
紫云英(Astragalus sinicusL.)和黑麦草(Lolium multiflorum Lam.)是我国稻区两种主要绿肥, 将其翻压还田可改善土壤理化性质, 并可部分替代氮肥, 减少过量氮肥投入带来的面源污染等生态问题[8, 9]。紫云英黑麦草翻压还田增加了稻田土壤中有机质含量, 影响水稻土壤环境, 使甲烷排放有所增加[1, 8]。本文研究紫云英和黑麦草鲜草翻压还田对双季稻不同生育时期稻田土壤中产甲烷古菌群落结构动态变化, 了解稻田甲烷排放变化机制, 为减少稻田温室气体排放提供理论支撑和参考依据。
湖南省岳阳市华容县万庾镇吴家桥村七组(29° 52′ N, 112° 55′ E)地处洞庭湖平原, 属亚热带季风湿润气候, 年均气温17℃, ≥ 10℃积温5273℃, 无霜期261 d, 年降雨量1232 mm。试验土壤类型为紫潮泥。经测定, 试验前土壤 pH 6.0, 含有机质5.07%, 土壤全氮0.18%, 碱解氮125.0 mg kg-1, 有效磷12.5 mg kg-1, 速效钾11.6 mg kg-1, 土壤容重1.18 g cm-3。
设置施用尿素(CF)、紫云英翻压还田(MV)、黑麦草翻压还田(RG)和不施氮肥(CK) 4个处理。小区面积5 m × 6 m=30 m2, 完全随机区组排列, 3次重复。小区间修筑0.5 m宽的田埂, 区组间用1 m宽的灌水沟隔开。各处理均以早稻和晚稻移栽前施用过磷酸钙(75 kg hm-2P2O5)和氯化钾(100 kg hm-2K2O)为基肥。CF施氮量为当地农民惯用施肥量(早晚稻各施200 kg N hm-2), MV和RG为紫云英和黑麦草按照田间直接生长情况在早稻移栽前1周翻压还田, 早稻和晚稻两季均不施用其他氮肥。紫云英施用量21 t hm-2 (鲜重), 含水量89.5%, C/N 14.5, 总有机碳含量514 g kg-1 (干重); 黑麦草施用量24 t hm-2 (鲜重), 含水量79.6%, C/N 62.3, 总有机碳含量553 g kg-1 (干重)。水稻移栽后进行喷施除草剂、灌水、中期晒田等常规管理措施, 晚稻移栽前进行翻耕和整地。黑麦草供试品种黑麦草特高(播种量30 kg hm-2), 紫云英供试品种湘肥3号(播种量37.5 kg hm-2)。早稻供试品种株两优819, 2011年5月1日移栽, 移栽密度每公顷200 000蔸, 7月20日收获。晚稻供试品种丰源优299, 移栽密度每公顷200 000蔸, 7月24日移栽, 10月26日收获。
分别于早稻分蘖期、拔节期、成熟期及晚稻分蘖期、扬花期、成熟期取水稻根际土(将水稻从水稻田中连根拔起, 多次抖动根部去除松散附着于根表的土壤, 收集仍然黏附于根表的土壤, 约距根为0~4 mm), 从每个处理取5株水稻根际土土样, 均匀混合, 后将土壤用锡箔纸包裹, 置液氮罐中迅速冷冻带回北京中国农业大学农学与生物技术学院, -20℃保存备用。
采用华越洋新型快速土壤DNA提取试剂盒(北京, 华越洋生物科技有限公司), 各处理均称取0.3 g土壤, 按试剂盒的试验步骤提取土壤微生物总DNA, 以微量紫外分光光度计ND-1000 (美国)检测其纯度和浓度。
以提取的土壤总DNA为模板, 利用产甲烷古菌的16S rRNA特异性引物1106F (5′ -TTWAGTCAGG CAACGAGC-3′ )和1378R (5′ -TGTGCAAGGAGCAG GGAC-3′ )进行扩增, 其中前引物带GC-clamp (5′ -C GCCCGCCGCGCGCGGCGGGCGGGGCGGGGGCACGGGGGG-3′ )。反应程序为94℃预变性90 s; 94℃变性30 s, 56℃退火30 s, 72℃延伸90 s, 33个循环; 72℃延伸3 min。PCR体系50 μ L, 含5 μ L 10× PCR buffer (含Mg2+), 4 μ L 2.5 mmol L-1 dNTP, 1 μ L 20 μ mol L-1 1106F-GC clamp, 1 μ L 20 μ mol L-1 1378R, 0.2 μ L 5 U μ L-1Taq DNA聚合酶, 1 μ L模板DNA, 37.8 μ L超纯水。用2.0%琼脂糖凝胶电泳检测PCR 300 bp左右的特异性扩增片段。
应用DNA片段纯化试剂盒(北京三博远志生物技术有限公司)按其说明书将产甲烷古16S rRNA的PCR产物浓缩至约200 ng μ L-1, 取10 μ L该样品加入5 μ L loading buffer, 应用Bio-Rad公司DCode 突变检测系统(Universal Mutation System), 选取16 cm× 16 cm胶板, 分离PCR产物。采取聚丙烯酰胺和变性剂双梯度凝胶, 变性梯度设为32%~62% (7 mol L-1尿素、40%去离子甲酰胺时的变性浓度为100%), 聚丙烯酰胺浓度为6%~12%。60℃, 0.5× TAE (DGGE专用)、100 V条件下电泳12 h。将DGGE胶采用SYBR Green I (1∶ 10000)染色30 min后, 用Alpha Innotech Chemilmage 5500 CCD成像仪照相。
分析并标记DGGE图谱中主要条带, 回收条带至1.5 mL离心管中。每个离心管中加入400 μ L无水乙醇, 10~15 min待胶变为白色, 真空吸走上清液, 再加入200 μ L diffusion buffer。待胶由白色变为透明后, 4℃过夜。最后用纯化试剂盒(北京, 华越洋生物科技有限公司)回收DNA, 置-20℃冰箱待用。
将电泳优势带回收后转化至大肠杆菌DH5α 感受态细胞, 连接pEASY-T3载体, 筛选蓝白斑, 阳性克隆菌液送北京三博远志公司测序。
利用Quantity one-4.6.2 (Bio-Rad)软件分析DGGE凝胶, 用得到的条带强度及位置信息做主成分分析(PCA), 并通过公式(1)、(2)计算Shannon-Weiner指数、丰富度指数(R):

其中, S为某一泳道的条带总数, ni为第i条条带的强度, N为所有条带总强度, pi为泳道中某一条带的强度与同泳道中所有条带强度的比值。
测序结果先经过DNAMAN软件编辑, 再登陆NCBI, 利用Blast搜索GenBank, 获取公开的相近菌株的16S rRNA基因序列, 利用Mega 4.0软件中N-J算法(Neighbor-Joining)建立系统发育树。
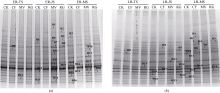
在早稻季, 产甲烷古菌16S rRNA基因DGGE图谱见图1-a, 不同处理不同时期的泳道中条带数相近(表1), 代表其菌种丰富程度相近。处理MV和RG在各时期的条带数略大于CF和CK。不同处理不同时期条带分离程度相似, 如ER-B16、ER-B12和ER-B22在所有处理及时期中均占优势, 说明该类产甲烷古菌受环境变化影响较小, 分布广泛; 个别条带为某时期所特有, 如ER-B5仅在早稻拔节期出现, 而ER-B20仅在早稻拔节期末出现。
 | 图1 早稻(a)和晚稻(b)不同生育时期稻田根际土壤中产甲烷古菌16S rRNA基因的DGGE图谱. CK: 不施氮肥; CF: 施用尿素; MV: 紫云英鲜草翻压还田; RG: 黑麦草鲜草翻压还田。ER-TS: 早稻分蘖期; ER-JS: 早稻拔节期; ER-MS: 早稻成熟期。LR-TS: 晚稻分蘖期; LR-FS: 晚稻扬花期; LR-MS: 晚稻成熟期。Fig. 1 DGGE profiles of amplified methanogenic archaeal 16S rRNA gene obtained from the rhizosphere soil of double-rice at different growing stages. CK: without nitrogen fertilizer; CF: with urea application; MV: with Chinese milk vetch incorporation; RG: with ryegrass incorporation; ER-TS: early-rice tillering stage; ER-JS: early-rice jointing stage; ER-MS: early-rice mature stage; LR-TS: late-rice tillering stage; LR-FS: late-rice flowering stage; LR-MS: late-rice mature stage. |
| 表1 双季稻不同生育期产甲烷古菌Shannon-Weiner指数和丰富度指数 Table 1 Shannon-Weiner (H) and Richness (R) indexes of methanogens derived from the rhizosphere soil of double-rice at different growing periods |
晚稻三生育期产甲烷古菌DGGE图谱如图1-b。在不同时期各处理间均存在优势条带(LR-B6、LR-B7、LR-B12、LR-B15、LR-B17、LR-B20、LR-B23), 但其强度差异微小。比较早稻晚稻相同生育期(分蘖期和成熟期), 晚稻各处理泳道的条带数明显多于早稻各处理, 表明晚稻季产甲烷古菌种的丰富度高于早稻季。
图2-a表明, 主成分因子1 (PC1)的贡献率为42.4%, 主成分因子2 (PC2)的贡献率为19.1%。PC1将样品分为2个部分, 早稻分蘖期和早稻拔节期的样品主要分布在图的左边, 早稻成熟期的样品主要分布在图的右边。在早稻分蘖期样品中, PC2将CK、CF处理的样品和MV、RG处理的样品分开; 早稻拔节期4个处理的样品分布非常接近; 早稻成熟期样品中, PC2将CF、RG处理的样品和MV、CK处理的样品分开。
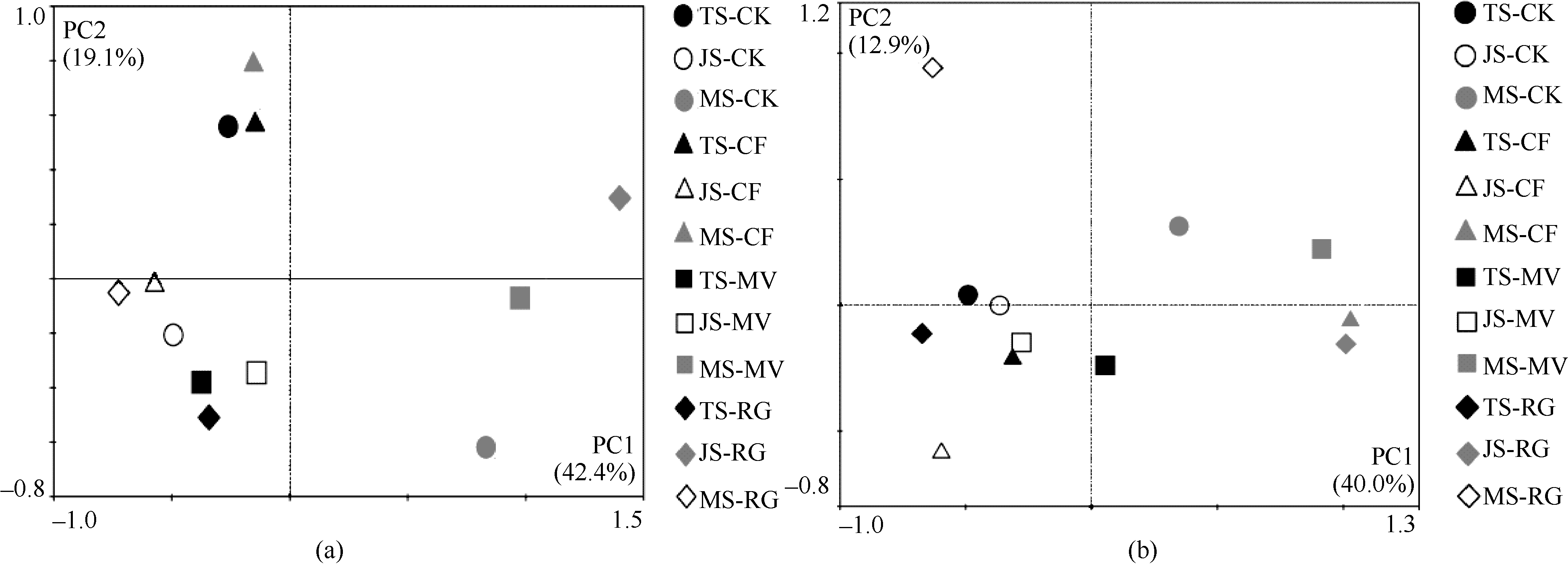 | 图2 早稻(a)和晚稻(b)PCR-DGGE图谱的主成分分析. 缩写同图1。Fig. 2 Principal component analysis of DGGE profiles (a: early-rice, b: late rice). Abbreviations are the same as those given in Figure 1. |
图2-b表明, 主成分因子1 (PC1)的贡献率为40.0%, 主成分因子2 (PC2)的贡献率为12.9%。PC1将样品分为2个部分, 晚稻分蘖期和晚稻扬花期的样品主要分布在图的左边, 晚稻成熟期的样品主要分布在图的右边。晚稻分蘖期4个处理的样品分布非常接近, 晚稻扬花期样品中, PC2将CK、MV和CF处理的样品同RG处理的样品分开; 晚稻成熟期样品中, PC2将CF、RG处理的样品和MV、CK处理的样品分开。
将早稻不同生育期产甲烷古菌DGGE图谱(图1-a)中所标示的条带切割, 回收测序, 序列经NCBI-Blast比对后, 得到最相近的菌种相关信息, 如表2所示。ER-B3序列比对得到最相近的菌为Uncultured clone SS033 (EU329770.1), 相似性仅为88%, ER-B22与Methanohalophilus mahii DSM 5219 (马氏甲烷嗜盐菌)菌种相似度为91%, 其他所有条带序列与NCBI数据库中菌种相似度均高于94%。
| 表2 早稻不同生育期产甲烷古菌DGGE条带序列比对结果 Table 2 Result of sequence alignment of methanogenic archaea at different growth stages of early rice |
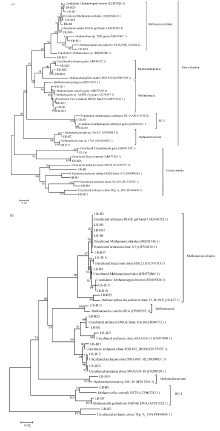
根据克隆测序和序列比对结果构建16S rRNA基因序列系统发育树, 如图3-a所示。早稻不同生育期各处理根际土中产甲烷古菌有甲烷微菌目(Methanomicrobiales)、Rice Cluster I (RC-I)、甲烷鬃菌科(Methanosaetaceae)、甲烷杆菌属(Methanobacterium), 还有较特殊的一类菌群是马氏甲烷嗜盐菌。
晚稻不同生育期产甲烷古菌DGGE凝胶条带回收、克隆测序结果经NCBI数据库Blast比对(表3)表明11条带测序结果比对后得到的是未能培养的菌种, 仅个别条带序列与已鉴定出的菌种有着高相似度, LR-B2、LR-B10、LR-B1序列比对后得到的结果均与Candidatus Methanoregula boonei strain SN19 (EU887826.1)相似度高, 分别为98%、99%和100%;
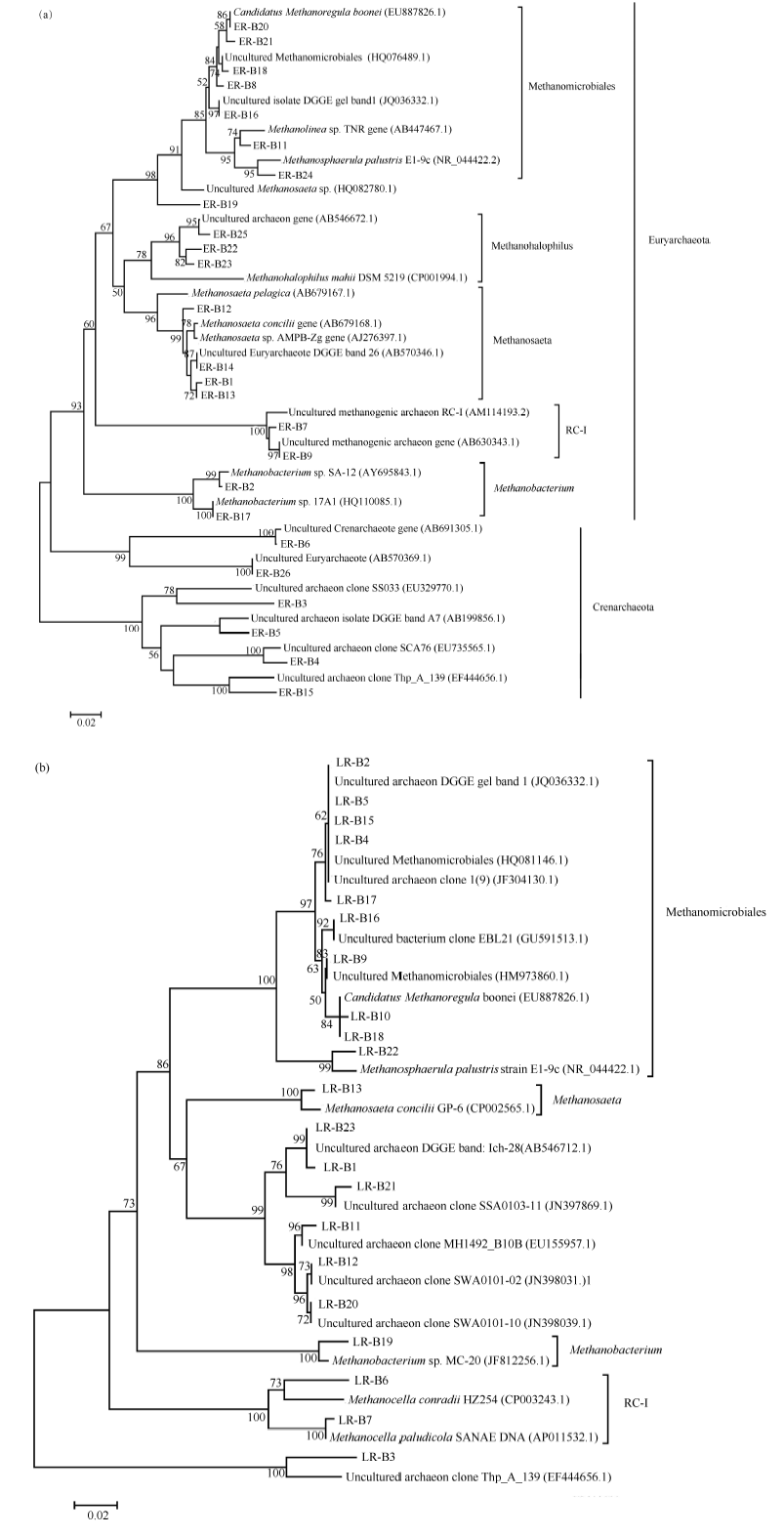 | 图3 早稻(a)和晚稻(b)不同生育时期产甲烷古菌16S rRNA基因序列系统发育树Fig. 3 Phylogenetic tree analysis based on methanogenic 16S rRNA gene sequences at different growth stages of early rice (a) and late rice (b) |
LR-B6和LR-B7为Methanocella(RC-I)属, LR-B13与Methanosaeta concilii GP-6 (甲烷鬃菌属)相似度为99%, LR-B19与Methanobacteriumsp. MC-20 (甲烷杆菌属)相似度为98%, LR-B22与Methanosphaerula palustris strain E1-9c相似度为98%。早稻和晚稻大部分条带序列比对得到较多相同菌种, 唯有相似度有些差异(表2和表3)。
| 表3 晚稻不同生育期产甲烷古菌DGGE条带序列比对结果 Table 3 Result of sequence alignment of methanogenic archaea at different growth stages of late rice |
如图3-b所示, 晚稻不同生育期根际土中主要群落与早稻相似, 优势群落主要有甲烷微菌、Rice Cluster I、甲烷鬃毛菌、甲烷杆菌属。晚稻季属于泉古菌门的古菌类群明显少于早稻季, 但因晚稻季有11条序列对比结果为不可培养的菌种, 未可确定其是否属于泉古菌门。
如表1所示, CK和CF在早稻季不同生育期Shannon-Weiner指数变化不大, MV和RG的Shannon-Weiner指数在各时期都较接近; 在早稻季3个时期Shannon-Weiner指数为MV> RG> CF> CK。
晚稻季中, MV的Shannon-Weiner指数变化比较有规律, 趋势是分蘖期> 扬花期> 成熟期; 同时期不同处理间差异较大, 分蘖期MV> RG> CK、CF; 扬花期CK的Shannon-Weiner指数较高, 成熟期除CK外, 其他处理Shannon-Weiner指数较接近。总体来讲, 晚稻季的Shannon-Weiner指数高于早稻季。
产甲烷古菌群落受取样时间变化小, 多数生育时期双季稻根际土中菌群落结构相似度高, 这与Krü ger等[10]对意大利水稻田中产甲烷古菌群落研究结果类似。Watanabe等[11]利用DGGE技术, 对稻-麦轮作体系产甲烷古菌研究中, 同样发现产甲烷古菌群落结构较稳定。但不论是早稻季还是晚稻季, PCA分析都将成熟期产甲烷古菌群落结构与其他时期分开, 可能原因在于成熟期稻田有机质含量低于其他时期。稻田土壤中有机质分解后产物可作为产甲烷的前体。紫云英和黑麦草还田后有机质迅速腐解, 早稻成熟期稻田有机质含量降至较低水平[12], 晚稻季中, 早稻根系和残茬为稻田中主要有机质来源, 其在晚稻成熟期也会降至较低水平[1]。并且, 水稻成熟期光合作用减弱, 根际分泌物减少[13]。总体来看, 成熟期稻田土壤中有机物质含量降低, 产甲烷前体减少, 可能使成熟期产甲烷古菌群落结构异于其他时期。晚稻季产甲烷古菌的Shannon-Weiner指数(H)和丰富度指数(R)均略高于早稻季, 可能原因在于晚稻季土壤温度较高, 刺激产甲烷古菌、土壤高分子水解菌和发酵菌的活性, 加剧稻田中有机物的分解[14, 15]。
此外, RC-I产甲烷古菌较丰富且稳定分布于双季稻各个生育时期, 很多研究结果已表明RC-I在根际微区中较其他类型产甲烷古菌活跃[4, 6, 17]。
与CK相比, 3个施肥的处理在双季稻不同生育期提高了产甲烷古菌的丰富度和多样性指数, 在施放有机肥的MV和RG中表现更明显。
产甲烷古菌的繁殖与生长需要充足的底物[17], 紫云英或黑麦草还田从两方面增加土壤中产甲烷古菌的底物, 一是其本身为稻田注入大量有机质, Zhu等[12]研究表明, 80%的有机干物质将在还田后的1个月内迅速腐解, 为产甲烷古菌提供大量产甲烷前体; 二是紫云英和黑麦草还田可促进水稻植株的生长, 水稻根系脱落物、分泌物及植株残体增加[8, 13], 进而增加土壤产甲烷前体数量。普通施放氮肥处理也可通过促进水稻植株的生长来实现产甲烷古菌底物的增加, 但无大量有机质注入, 对产甲烷古菌的影响不如紫云英或黑麦草还田。
产甲烷古菌需要严格的厌氧环境, 在氧气或其他氧化态无机化合物存在的情况下, 产甲烷古菌将会丧失活性, 逐渐死亡[17]; 而在严格厌氧和还原条件下(氧化还原电位Eh ≤ -200 mV), 产甲烷古菌将保持活性, 将产甲烷前体转化为甲烷[18]。紫云英和黑麦草有机质分解时, 土壤兼性细菌和厌氧细菌将消耗土壤中大量氧气和氧化态无机化合物(NO3-、Mn4+、Fe3+、SO42-)并产生还原物质, 使得稻田氧化还原电位快速下降, 为产甲烷古菌活动提供必要环境[13, 19]。
除了必备的碳源(有机质分解产生的产甲烷前体)和土壤还原条件外, 产甲烷古菌的生存还需氮源, 而所有产甲烷古菌都可将NH4+作为氮源, 即使肽和氨基酸存在, NH4+仍为产甲烷古菌所必需[17]。紫云英或黑麦草还田、施放铵态氮肥尿素, 均可为产甲烷古菌的生长提供必需的NH4+。Zhu等[12]研究表明, 紫云英或黑麦草还田对产甲烷古菌群落影响期较施用尿素处理长, 可能更有利于产甲烷古菌的繁殖生长。
紫云英或黑麦草还田不论是在碳源、氮源供给, 土壤环境等方面均明显比施用尿素更有利于产甲烷古菌的生长, 但文中结果显示其暂未对产甲烷古菌群落结构产生决定性影响, 表明仍需长期定位试验来进一步明确紫云英或黑麦草还田对产甲烷古菌群落的影响。
早稻季和晚稻季各取样时期, 产甲烷古菌种类变化不大, 优势条带类似, 优势群落主要有广古菌门的甲烷微菌、Rice Cluster I、甲烷鬃菌、甲烷杆菌属。早稻季中还存在特殊的一类菌群, 马氏甲烷嗜盐菌。双季稻季节内取样时期和处理对产甲烷古菌群落结构总体影响不大, 早稻分蘖期和拔节期根际土中菌群落结构相似度较高, 晚稻分蘖期和扬花期群落相似度较高, 早稻和晚稻成熟期产甲烷古菌群落结构与其他时期相似度较低。晚稻季产甲烷古菌群落的Shannon-Weiner指数(H)和丰富度指数(R)均略高于早稻季。施肥处理(施用尿素、紫云英鲜草翻压还田和黑麦草鲜草翻压还田提高了双季稻不同生育时期稻田根际土壤中产甲烷古菌群落的丰富度和多样性指数, 紫云英和黑麦草还田的处理较尿素处理提高的更为明显。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|