



{kind=link}
{kind=link}
{kind=link}
{kind=link}
去除遮阴后自然强光对小偃54和8602及其杂交后代光合特性的影响
[周连杰1  , 谢彦庆
, 谢彦庆1 , 徐文1 , 郭骞欢1 , 戴双2 , 赵世杰1, * , 宋健民3 ]
, 谢彦庆, 宋健民|
|


第一作者联系方式: E-mail: lianjie_zh@163.com, Tel: 13951595720
黄淮地区冬小麦生育后期常出现阴雨寡照及连阴骤晴等不良气候, 本试验目的是阐明弱光逆境解除后自然强光对冬小麦光合特性的影响。在抽穗至开花期对小偃54和8602及其杂交后代小偃81和212遮阴处理(约自然光40%), 10 d后去除遮阴, 测定恢复过程中亲本与杂交后代旗叶叶绿素含量、光合气体交换、叶绿素荧光等参数。去除遮阴初期, 亲本与杂交后代都发生了光抑制, 净光合速率( Pn)、叶绿素 a/ b比值、胞间CO2浓度( Ci)、气孔导度( Gs)、光系统II (PSII)最大量子效率( Fv/ Fm)、PSII实际光化学效率( ΦPSII)、非光化学猝灭(NPQ)和电子传递到QA下游的概率( Ψo)都低于对照, 并且随着去除遮阴时间的延长逐渐恢复。杂交后代小偃81在去除遮阴后比其2个亲本和另一杂交后代品系212具有相对更高的 Pn、 ΦPSII和 Ψo, 表明小偃81对环境光强变化的适应性优于其亲本, 且相对较高的PSII光化学效率活性和稳定性是其在去除遮阴后拥有更高 Pn的原因之一。小偃81更能适应黄淮地区小麦生育后期光照不足和连阴骤晴的不良气候。
, XIE Yan-Qing, SONG Jian-MinShading problem often occurs at the late growing period of winter wheat in the Huang-Huai Plain of China. The objective of this study was to explore the effect of full sunlight after the shading treatment on photosynthetic characteristics in wheat. In a filed experiment with 10-day shading (40% of natural light intensity) from heading to anthesis, the photosynthetic characteristics of wheat varieties Xiaoyan 54 and 8602 and their hybrids Xiaoyan 81 and 212 were measured at 0, 1, 3, 5, 7, and 10 days after shading treatment. Photoinhibition was observed in all genotypes when the shading ended, and the photosynthetic rate ( Pn), ratio of chlorophyll a to chlorophyll b, intercellular CO2 concentration ( Ci), stomatal conductance ( Gs), maximum quantum yield of PSII ( Fv/ Fm), actually photochemical efficiency of PSII ( ΦPSII), nonphotochemical quenching (NPQ) and efficiency that an electron moves to QA downstream ( Ψo) were lower than that of the control (no shading treatment). However, such reductions were alleviated gradually with the time lasting after shading. The hybrid Xiaoyan 81 had the highest Pn, ΦPSII, and Ψo as compared with other tested genotypes, showing its good adaptability to the change of light intensity. The higher Pn in Xiaoyan 81 than in its parents after shading probably resulted from the higher photochemical efficiency of PSII in Xiaoyan 81. These results suggest that Xiaoyan 81 is a wheat variety suitable to grow in the Huang-Huai Plain with low light intensity problem at late growth period of winter wheat.
冬小麦生育后期常出现阴雨寡照及连阴骤晴等不良气候条件, 而光照强度的大幅度变化会明显地影响植物叶片的光合作用能力、植物对营养物质的吸收及其在植物体内的重新分配等一系列生理生化过程, 造成减产。研究表明, 小麦开花期, 随着遮阴强度的增加, 旗叶实际光化学效率(Φ PSII)升高, 净光合速率(Pn)下降[1]; 弱光下叶绿体结构被破坏, 光系统II (PSII)活性受抑[2]; 遮阴使小麦旗叶叶绿素a和叶绿素b含量增加, 但叶绿素a/b的比值降低[3]; 从拔节期开始对小麦遮阴处理, 随着遮阴强度的加大, 蒸腾速率(Tr)显著降低, 同时胞间CO2浓度(Ci)显著升高[4]。透光率为57.3%的遮光处理导致小麦物候期推迟2~4 d, 前期生物量增加, 但从抽穗期开始由于光照严重不足, 春季分蘖和下部叶片大量死亡, 最终导致地上生物量和籽粒产量下降[5]。杂交水稻抽穗期弱光胁迫10 d后恢复自然光照, 旗叶Pn随着时间的推移与对照的差异减小[6]。小麦遮阴处理15 d后, 叶片光能吸收能力增加, 但光能利用能力及过剩光能耗散能力降低[7]。上述研究主要侧重弱光胁迫对农作物光合特性的影响, 而对于由弱光环境转入自然光照后光合作用的适应和恢复过程研究较少, 忽略了前期弱光胁迫对恢复光照后农作物生长发育的持续影响。因此, 研究弱光胁迫及胁迫解除后自然强光对冬小麦光合特性的影响具有重要意义。
本研究旨在阐明短时间遮阴处理去除遮阴后自然强光对不同冬小麦品种及其杂交后代光合特性的影响, 以期为小麦高光效高产育种和高产抗逆栽培提供理论依据。
供试材料中, 小偃54是中矮秆、抗逆性强的优质小麦品种, 水肥利用效率高; 8602茎秆较粗壮, 旗叶直立转披, 光化学效率较高, 丰产性好。以这2个品种为亲本(小偃54× 8602)选育杂交后代, 获得高代(F7)稳定品系212及育成品种小偃81, 而且小偃81已经推广种植。
2012— 2013生长季, 将两亲本和2个杂交后代播种于山东农业大学实验基地, 每个基因型6个小区, 其中3个被遮阴处理, 另外3个重复为自然光照(对照)。田间随机区组排列, 小区面积3 m × 4 m, 行距25 cm, 播种量11.5 g m-2, 均匀撒播。生育期内按高产田常规栽培措施管理。
由于在冬小麦生育后期天气转阴时多数时间的光照强度是自然光照的40%左右, 故本试验选用透光率为40%的黑色遮阴网制成可拆卸遮阴棚进行抽穗期至开花期的遮阴处理。遮阴10 d后去除遮阴网, 恢复自然光照直至小麦成熟。遮阴网高度距离小麦冠层顶部50 cm左右, 以保证冠层通风良好。分别在去除遮阴后的第0、第1、第3、第5、第7和第10天测定旗叶的各项生理指标。
根据Arnon[8]的方法, 用80%丙酮黑暗浸提48 h, 其间摇动数次, 叶片色素经完全提取后用UV-1601紫外可见分光光度计(岛津, 日本)测定其叶绿素含量。
选择晴天上午8:30至10:30, 用CIRAS-2光合作用测定系统(PP Systems, 美国)测定旗叶的Pn、Gs、Ci, LED光强为1200 μ mol m-2 s-1, CO2浓度为380 μ mol mol-1, 采用大气湿度和温度。
利用FMS-2便携式叶绿素荧光测定系统(Hansatech, 英国)测定旗叶PSII的Φ PSII和非光化学猝灭(NPQ)。于上午9:00-10:30, 经暗适应30 min后, 利用Handy-PEA植物效率分析仪(Hansatech, 英国)测定旗叶叶绿素荧光动力学曲线并计算相关的荧光参数[9, 10]。
用DPS 3.01软件统计分析数据, 用Microsoft Excel 2003绘图。
抽穗期遮阴处理使冬小麦旗叶的叶绿素总量上升, 各品种上升幅度由大到小依次为212、8602、小偃81和小偃54 (图1-A, B), 表明弱光胁迫下植株通过提高叶片叶绿素含量增加光能吸收, 从而提高光合能力。去除遮阴后, 随着转入自然强光下时间的延长, 4个基因型叶片的叶绿素总量逐渐降低, 其中品种小偃54和小偃81在第3天即下降到对照水平, 而品系212在第10天才下降到对照水平。在遮阴及去除遮阴第1天, 叶绿素a/b比值下降幅度较大, 显著低于对照水平, 但亲本与杂交后代间差异不显著, 且都在去除遮阴后第3天即恢复至对照(图1-C, D)。
 | 图1 去除遮阴后不同冬小麦品种(系)叶绿素总量及叶绿素a/b比值的变化. A和C: 去除遮阴; B和D: 无遮阴对照。小偃54 (XY54)和8602分别为杂交母本和父本; 小偃81 (XY81)和212为杂交后代。Fig. 1 Changes in total content of chlorophyll and ratio of chlorophyll a to chlorophyll b in different wheat genotypes after shading treatment. A and C: after shading treatment; B and D: control without shading treatment. Xiaoyan 54 (XY54) and 8602 are female and male parents, respectively; Xiaoyan 81 (XY81) and 212 are hybrids. |
遮阴处理10 d后, 4个冬小麦品种(系)小偃54、小偃81、212和8602旗叶的Pn均显著低于对照(图2-A, B), 分别为对照的36.8%、40.9%、34.5%和38.0%, 且在去除遮阴后的第5天杂交后代品种小偃81和亲本8602的Pn即已恢复至对照水平, 而小偃54和212的Pn在去除遮阴后的第10天才恢复至对照, 表明小偃81和8602两个品种更能适应由弱光到强光的转变。
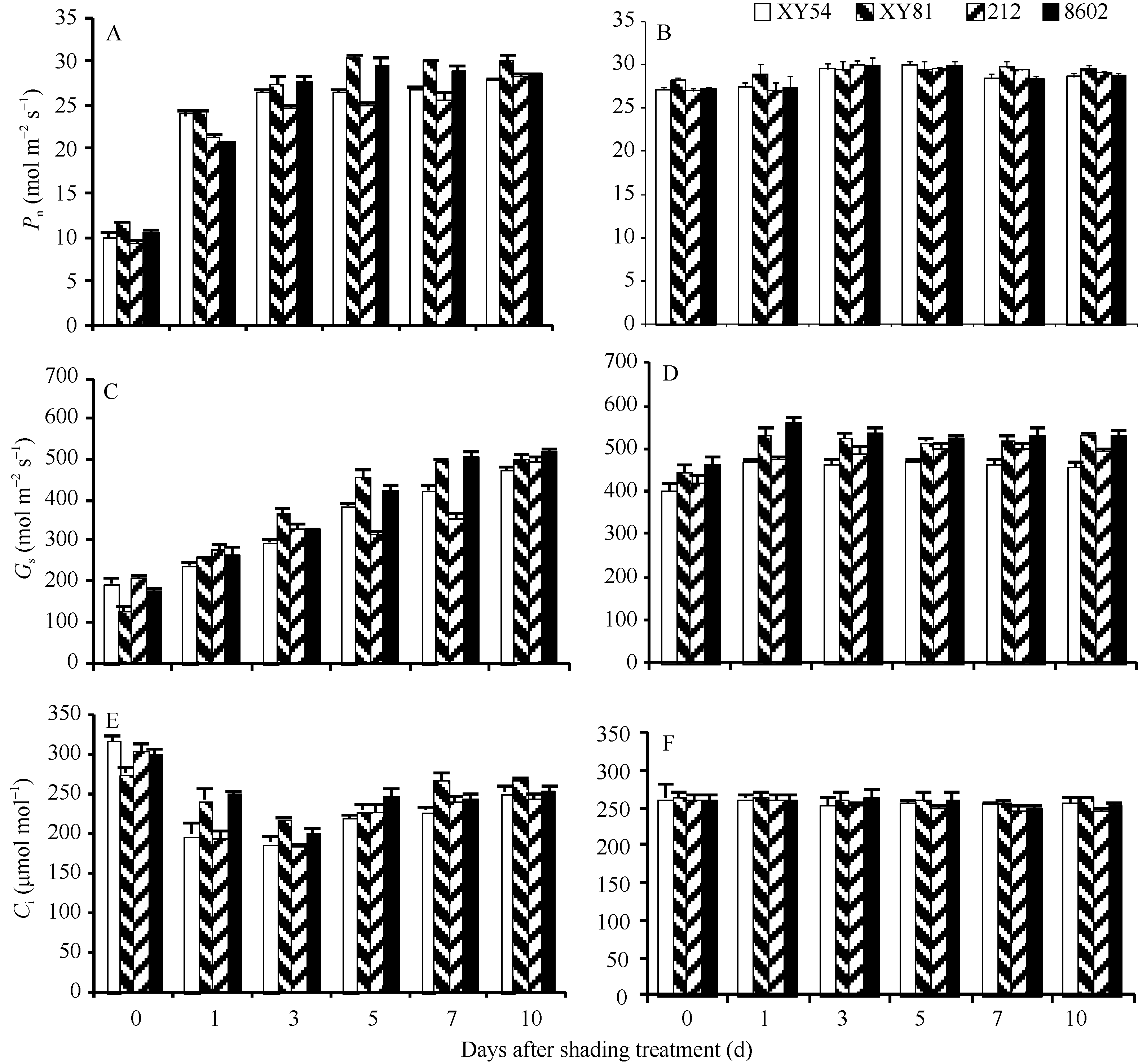 | 图2 去除遮阴后不同品种(系)冬小麦光合速率、气孔导度及胞间CO2浓度的变化. A、C和E: 去除遮阴; B、D和F: 无遮阴对照。小偃54 (XY54)和8602分别为杂交母本和父本; 小偃81 (XY81)和212为杂交后代。Fig. 2 Changes in photosynthetic rate, stomatal conductance, and intercellular CO2 concentration in wheat flag leaves after shading treatment. A, C and E: after shading treatment; B, D and F: control without shading treatment. Xiaoyan 54 (XY54) and 8602 are female and male parents, respectively; Xiaoyan 81 (XY81) and 212 are hybrids. |
在遮阴及去除遮阴初期, 4个基因型的Gs均低于对照(图2-C, D), 小偃54、小偃81、212和8602分别为对照的48.4%、27.8%、48.4%和37.4%, 以小偃81下降幅度最大。随着恢复时间的延长, Gs逐渐上升, 4个基因型间差异显著, 其中小偃54和212去除遮阴后第10天才恢复至对照水平, 而小偃81和8602在转入自然光第7天即恢复至对照水平。
去除遮阴前(第0天), 4个基因型的旗叶Ci高于对照(图2-E, F), 但此后迅速下降, 并低于对照, 至第5天开始逐渐升高, 至第7天恢复到对照水平。在去除遮阴后的第1天, 小偃54和212的Ci下降幅度较大, 分别比对照下降24.2%和25.4%, 且在恢复过程中始终低于小偃81。
冬小麦Fv/Fm在遮阴时升高, 去除遮阴后迅速下降(图3-A, B), 去除遮阴后第1天亲本和杂交后代都出现出不同程度的光抑制, 其中亲本8602光抑制最严重。随着恢复时间延长, Fv/Fm升高至对照水平, 光抑制解除。弱光下亲本和杂交后代的Φ PSII都高于对照(图3-C, D), 去除遮阴后Φ PSII迅速下降, 杂交后代小偃81下降幅度最小; 小偃81和8602在第5天已恢复至对照水平, 而小偃54和212则在第7天才恢复。遮阴使冬小麦NPQ降低(图3-E, F), 其中杂交后代小偃81降低幅度最小, 仅为对照的66.5%, 表明在弱光下小偃81用于耗散的能量相对较少。去除遮阴后, 亲本和杂交后代的NPQ均逐渐恢复至对照水平, 随着时间延长, 除亲本8602外, 其余基因型的NPQ无显著差异, 在去除遮阴后第5天即恢复至对照。
 | 图3 去除遮阴后不同冬小麦品种(系) Fv/Fm、Φ PSII和NPQ的变化. A、C和E: 去除遮阴; B、D和F: 无遮阴对照。小偃54 (XY54)和8602分别为杂交母本和父本; 小偃81 (XY81)和212为杂交后代。Fig. 3 Changes in Fv/Fm, Φ PSII, and NPQ in wheat flag leaves after shading treatment. A, C, and E: after shading treatment; B, D, and F: control without shading treatment. Xiaoyan 54 (XY54) and 8602 are female and male parents, respectively; Xiaoyan 81 (XY81) and 212 are hybrids. |
遮阴及去除遮阴后, 4个基因型的K点可变荧光(Fk)占FJ-FO荧光振幅的比率(Wk)均高于对照, 但杂交后代与亲本间差异不显著, 并且都在转入自然强光后的第10天恢复到对照水平(图4-A, B)。在去除遮阴后的第1天, 小偃54、小偃81、212和8602电子传递到QA下游的概率(Ψ o)分别降至对照的94.8%、96.8%、92.2%和89.3%, 以小偃81下降幅度最小。在随后的恢复过程中, 4个基因型的Ψ o均表现为先下降后上升趋势, 在转入自然强光后的第7天恢复至对照水平(图4-C, D)。
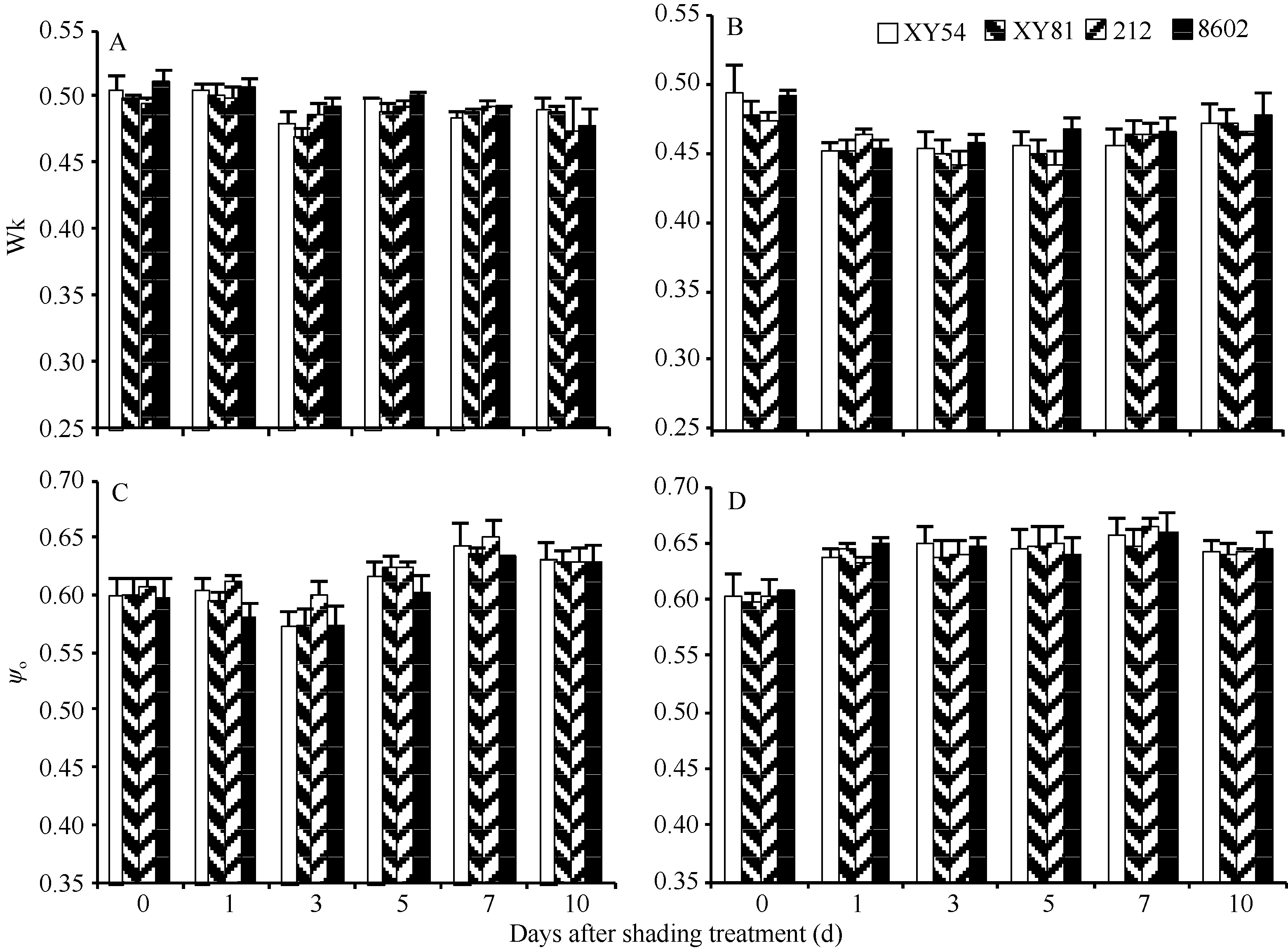 | 图4 去除遮阴后不同冬小麦品种(系)Wk和Ψ o的变化. A和C: 去除遮阴; B和D: 无遮阴对照。小偃54 (XY54)和8602分别为杂交母本和父本; 小偃81 (XY81)和212为杂交后代。Fig. 4 Changes in Wk and Ψ o in wheat flag leaves after shading treatment. A and C: after shading treatment; B and D: control without shading treatment. Xiaoyan 54 (XY54) and 8602 are female and male parents, respectively; Xiaoyan 81 (XY81) and 212 are hybrids. |
植物通过光合色素捕获光能进行光合作用, 光照的强弱会影响色素的形成和分布, 进而影响光合作用。弱光胁迫下单位面积叶片的叶绿体数目减少, 但叶绿体变大, 叶绿素含量增加[2, 11], 叶绿素a/b比值降低[3, 12]。但亦有相反的报道, 如遮阴可以缓解夏玉米生育后期叶绿素a的降解, 使叶绿素a/b比值升高, 且在夏玉米穗期遮阴后叶绿素含量不能恢复至对照水平[13], 这可能与不同试验、不同作物、不同基因型和不同处理时期等有关。本试验结果表明, 抽穗期约40%透光率遮阴10 d后, 冬小麦叶片叶绿素含量升高(图1-A, B), 叶绿素a/b比值降低(图1-C, D)。与对照比, 杂交后代212的叶绿素总量上升幅度最大, 亲本8602其次, 亲本与杂交后代间叶绿素a/b比值无显著差异。去除遮阴后, 叶绿素逐渐降解并最终达对照水平, 说明光照条件变化时植株通过动态调节叶绿素含量以适应不同的光照条件。
光照强度是影响植物光合作用效率的重要因素之一, 在光饱和点以下, 随着光照强度减弱, 叶片Pn降低[3, 6, 14, 15, 16, 17]。遮阴使冬小麦叶片光合速率和气孔导度降低(图2-A~D), 胞间CO2浓度升高(图2-E, F), 且对杂交后代212的影响最大。亲本与杂交后代在去除遮阴初期净光合速率均低于未遮阴处理, 随后逐渐恢复, 其中杂交后代小偃81和亲本8602恢复较快, 而亲本小偃54和另一杂交后代212恢复较慢, 表明小偃54和212对由弱光转入自然强光的适应性较差。Yang等[18]认为, 小偃54具有较强的抗寒和抗旱能力, 与本试验结果不一致, 可能是因为逆境环境不同。去除遮阴后亲本与杂交后代Pn的恢复趋势与旗叶总叶绿素含量变化趋势一致。在恢复自然光照后, 亲本与杂交后代品种Gs和Ci均低于对照, 但在随后恢复过程中其变化趋势与Pn变化趋势略有不同, 表明在转入自然强光后Pn降低是由气孔限制和非气孔限制共同造成的[4, 19, 20]。
叶绿素荧光参数可以从光合作用内部变化的角度进一步解释植株对光强变化的适应性[21]。李潮海等[22]研究发现, 由遮阴转入自然光照后, Fv/Fm迅速下降, 4~5 d后逐渐回升。贾士芳等[23]亦指出, 弱光胁迫结束后随着恢复时间延长, 玉米Fv/Fm可逐渐恢复至对照。本试验也有类似发现, 说明叶片在经过弱光处理转入自然光后PSII对光能的利用效率受到抑制[24], 随着恢复时间延长, 叶片对自然光照逐渐适应, PSII对光能的利用效率得到恢复, 但亲本与杂交后代间的趋势不同, 其中8602和小偃81的恢复能力高于小偃54和212, 即8602和小偃81在转入自然强光后能以较快的速度解除光抑制, 进而实现对高光强的高效利用。从本研究结果可以看出, 遮阴期间Φ PSII升高, NPQ显著降低, 这与Xu等[1]的研究结果一致, 说明虽然小麦在弱光下获得的光能减少, 但可以通过提高PSII实际光能转化效率, 减少非辐射能量的耗散来补偿光照不足引起的Pn下降, 遮阴提高叶片对光能的吸收和转化效率是一种弱光逆境的自我补偿机制。去除遮阴后, 亲本与杂交后代品种的Φ PSII均出现不同程度下降(图3-C, D), 说明一段时间遮阴后植株已在一定程度上适应了弱光条件, 转入自然光照后光能利用能力均降低, 且小偃81降低幅度最小, 8602次之, 说明去除遮阴后自然强光对小偃81和8602光化学活性影响较小。相对较高的PSII光化学活性是杂交后代品种小偃81在去除遮阴后Pn高于两亲本和另一杂交后代品系的原因之一。光化学反应除了包括CO2同化外, 还包括Mehler反应、光呼吸、氮代谢等[25, 26, 27]。去除遮阴后第7天, 小偃54的Φ PSII已恢复至对照水平, 但Pn尚未恢复, 表明抗逆品种小偃54还通过其他光化学途径增加了对吸收光能的耗散。在去除遮阴后, 亲本与杂交后代品种的非光化学耗散能力虽然有所升高, 但仍低于对照, 且品种间无显著差异, 表明短期的弱光胁迫后, 旗叶用于耗散的能量减少, 但随后NPQ均很快恢复, 说明植株通过提高非辐射性热能耗散的形成来耗散不能被光化学转化的过剩光能。
Wk反应PSII供体侧的受伤害程度, Wk越高, PSII供体侧特别是放氧复合体受伤害程度就越大[28, 29], Ψ o表示光合电子从QA传递给下游电子传递体的能力[9, 29]。遮阴10 d及去除遮阴初期, 4个基因型的Wk与对照比都有明显升高, 但亲本与杂交后代间并无显著差异, 且均在一段时间后恢复至对照水平(图4-A, B), 表明造成亲本与子代间光合差异的原因并不是PSII供体侧受到伤害程度的不同。在弱光下, 亲本与杂交后代品种的Ψ o与对照比并无显著差异(图4-C, D), 表明弱光下PSII的受体侧并没有受到伤害。去除遮阴后, 4个基因型Ψ o逐渐下降, 且8602下降幅度最大, 表明其QA以后电子传递能力受到影响更大, 即PSII受体侧受到伤害程度大于其他3个品种, 可能是其转入自然强光第1天Pn较低的原因所在, 但在去除遮阴后第7天, 所有基因型电子传递到QA以后的能力都已恢复, 不再是造成光合低于对照的原因。
小麦产量由单位面积穗数、每穗粒数和每粒粒重构成, 而光合产物是小麦产量的根本来源, 其中旗叶是主要功能叶, 小麦籽粒产量的大部分都来源于其光合产物。因此, 旗叶的光合能力在很大程度上决定了小麦的产量。在去除遮阴后, 杂交后代小偃81旗叶的光合能力迅速恢复至对照水平, 表明其光合强度的变化小于另外3个品种, 但植株光合产物形成的多少除了取决于光合强度外, 还与光合时间、光合有效面积、光合消耗等因素有关。而本文主要是通过对光合电子传递链的分析来研究不同品种冬小麦在去除遮阴后光合能力存在差异的原因, 对产量及其构成因素涉及较少, 去除遮阴后自然强光对亲本与杂交后代产量的影响尚需进一步研究。
去除遮阴后自然强光对亲本小偃54的影响低于另一亲本8602, 而杂交后代小偃81光合特性的受影响程度低于两亲本, 说明可以通过遗传改良提高小麦后代的光合特性和抗逆能力, 进而适应北方地区小麦生育后期光照不足、连阴骤晴等不良气候条件。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|