
{kind=link}
{kind=link}
{kind=link}
{kind=link}
水稻白穗突变体 wp4的鉴定与基因精细定位
[王晓雯 , 蒋钰东, 廖红香, 杨波, 邹帅宇, 朱小燕, 何光华
, 蒋钰东, 廖红香, 杨波, 邹帅宇, 朱小燕, 何光华* , 桑贤春* ]
, 蒋钰东, 廖红香, 杨波, 邹帅宇, 朱小燕, 何光华, 桑贤春]
|
|


第一作者联系方式: E-mail:xwwang78@126.com
水稻开花灌浆期, 内外颖呈绿色, 含光合色素, 为阐释非叶片组织叶绿体发育的分子机制, 本文对EMS诱变获得的新型白穗突变体 wp4进行了研究。抽穗灌浆期, wp4的穗轴呈绿色, 内外颖呈乳白色; 内外颖叶绿体发育不健全, 无类囊体结构, 叶绿素 a、叶绿素 b和类胡萝卜素含量极显著降低。此外, wp4全生育期叶片呈淡黄色, 叶绿体内基质片层较少且排列松散; 与野生型相比, wp4的叶绿素 a含量显著降低, 叶绿素 b和类胡萝卜素含量虽略低, 但差异未达到显著水平。农艺性状分析发现, 与野生型相比, wp4的有效穗和结实率略有增加, 其他性状则略有降低, 但变化均未达到显著差异水平。遗传分析表明 wp4受一对隐性核基因调控, 利用1200株西农1A/ wp4的F2隐性群体, 最终将 WP4精细定位在第8染色体约79 kb的物理范围内, 根据水稻基因组注释计划, 该区间包含14个基因, 这为 WP4的功能和作用机制研究奠定了基础, 也为水稻标记性状育种提供了新的资源。
, JIANG Yu-Dong, LIAO Hong-Xiang, YANG Bo, ZOU Shuai-Yu, ZHU Xiao-Yan, HE Guang-Hua, SANG Xian-Chun
Rice Research Institute, Chongqing Key Laboratory of Application and Safety Control of Genetically Modified Crops, Southwest University, Chongqing 400716, China
At the flowering and filling stages, rice hull appears green color and possesses photosynthetic pigment. To clear the mechanism of chloroplast development in the panicle, we identified a novel white panicle mutant from the progeny of indica restorer line Jinhui 10 with seeds treated by EMS and termed it as wp4. The wp4 displayed green spike-stalks and milk-white hulls after the heading stage. The structures of chloroplast and thylakoid were severely destroyed and the contents of photosynthetic pigment decreased extremely significantly in the mutational hulls. Compared with the wild type, the wp4 displayed yellow green leaves and contained looser stromal lamellae. And the contents of chlorophyll a, b, and carotenoid were all declined while only the changing of chlorophyll a led to the significantly different level in statistics. Except for effective panicles number and seed setting rate, other detected agronomic characteristics decreased slightly but the changing did not come up to the statistically significant difference. Genetic analysis indicated that the white panicle of wp4 was controlled by a recessive nuclear gene and which was finally mapped on chromosome 8 with 79 kb physical distances according to 1200 mutational plants derived from the F2 generation of Xinong1A/wp4. The restricted region contained 14 annotated genes based on Rice Genome Annotation Project. These results provided a foundation for gene cloning and function analysis of the WP4. Meanwhile, the traits of wp4 could be available in rice breeding as a morphological marker.
叶片是水稻进行光合作用的主要场所, 抽穗灌浆期, 光合产物从叶输送到籽粒并储存下来, 最终形成稻谷产量。稻穗抽出剑叶叶鞘后, 颜色由白转为绿色, 授粉后, 随着籽粒的灌浆成熟, 又由绿渐变为金黄色。与叶片相比, 穗部叶绿体色素含量较少、光合效率较低, 穗部光合产物仅占水稻植株总光合产物的5.4%左右[1]。穗距离籽粒近, 根据“ 库” “ 源” 关系, 穗光合作用对水稻产量可能也具有一定的影响。如, 抽穗灌浆期套不透明袋, 水稻千粒重和结实率明显下降, 套透明袋则下降不明显[2]。但穗部光合作用对水稻产量的影响大小及穗叶绿体发育的分子机制还很不清晰。
在植物中, 叶绿体主要存在于叶片中, 叶绿体发育异常通常会导致叶色变化。叶色变异是一个复杂的生理过程, 受温度、光照、营养等外界环境的影响, 也受内部基因的调控。目前, 在水稻中已鉴定到160多个叶色突变体, 邓晓娟等[3]根据表型将其归为白化、黄化、温敏变色、亮绿、常绿、条斑条纹、转绿和紫色8类。目前, 已利用这些突变体克隆了30多个调控基因, 从而为叶片组织叶绿体发育和叶绿素合成的分子机制阐释奠定了基础[3, 4, 5]。然而, 对于非叶片组织, 由于相关突变体的匮乏, 叶绿体发育分子机制还很不清晰。在水稻中仅鉴定到6个穗部失绿突变体, 根据穗部表型分为两类, 一类仅内外稃白化失绿, 如wp1[6]、wlp1 (white leaf and panicles 1)[7]和wslwp (white striped leaf and white panicle)[8]; 一类除内外稃白化外, 穗轴和枝梗也呈白色, 如wp2[6]、wp(t)[9]和st-fon(streaked leaf and floral organ number)[10]。
为研究穗部光合作用对水稻产量的影响以及穗部叶绿体的发育机制, 我们从籼型水稻恢复系缙恢10号的EMS诱变库中筛选鉴定到1个新型白穗突变体。该突变体全生育期叶色淡黄, 抽穗灌浆期内外颖白色, 穗轴和枝梗不失绿, 暂命名为wp4(white panicle 4)。本文对其进行了表型鉴定和基因精细定位等研究。
白穗突变体wp4来源于晚籼水稻恢复系缙恢10号的EMS诱变, 多代自交后突变表型已稳定遗传。配制西农1A/wp4杂交组合, 利用其F1和F2群体进行遗传分析, 并利用F2分离群体中的隐性单株进行基因定位。西农1A是西南大学水稻研究所选育的一个早籼不育系, 穗和叶片颜色正常。
1.2.1 形态鉴定 田间种植wp4和野生型, 株行距20 cm × 25 cm, 常规管理。根据叶龄全生育期观察叶色变化, 抽穗期之后每隔1 d记载穗部颜色一次, 完全成熟后选取小区试验的中间10株, 调查株高、有效穗、穗长、穗粒数、结实率和千粒重等主要农艺性状, 并进行统计分析。
1.2.2 光合色素含量测定 开花期, 早晨8:30左右, 采集wp4及野生型的倒一叶和颖壳, 参照Wellburn[11]描述的方法测定光合色素含量。
1.2.3 透射电镜观察 参照Fang等[12]方法取开花期的叶片和颖壳, 经戊二醛和锇酸双重固定后, 利用不同梯度的乙醇逐级脱水, 再置换和包埋, 制超薄切片后, 以醋酸双氧铀和柠檬酸铅液双重染色, H600型透射电镜观察并照相。
利用改良的CTAB法提取亲本、基因池和隐性定位群体的DNA。把0.2 g左右的叶片剪碎放入预装3粒钢珠的2 mL离心管, 液氮速冻, 研碎后加入0.7 mL的CTAB溶液, 65℃水浴15 min, 每5 min颠倒1次, 再加入0.7 mL的氯仿萃取1 min, 12 000转 min-1离心3 min。取0.5 mL上清液到1.5 mL离心管内, 加1 mL预冷的无水乙醇, 上下颠倒后12 000转 min-1离心1 min, 倾去上清液, 无乙醇气味后加200 μ L的ddH2O溶解沉淀, 即得DNA。
RM系列SSR引物的序列来源于http://www. gramene.org/microsat。PCR总体积12.5 μ L, 包括1.25 μ L的10× PCR buffer, 0.75 μ L的25 mmol L-1 MgCl2, 0.5 μ L的2.5 mmol L-1 dNTPs, 8.0 μ L的ddH2O, 1.0 μ L的10 μ mol L-1引物, 1.0 μ L的模板DNA和0.1 μ L的5 U μ L-1Taq DNA聚合酶。PCR程序为94℃预变性3 min后, 94℃ 20 s、56℃ 20 s、72℃ 40 s, 35个循环, 再72℃充分延伸7 min。PCR产物经10%的非变性聚丙烯酰胺凝胶电泳, 0.2%的AgNO3染色10 min, 去离子水漂洗2次, 每次30 s, 然后再用0.1%的甲醛和2%的NaOH混合液染色至条带清晰可辨, 去离子水冲洗2次后观察照相。
前期研究中, 已利用高通量测序技术完成了西农1A和缙恢10号的DNA测序。根据二者之间的多态性, 选择6~12个碱基的差异位点, 利用NTI vector 11.0软件开发Indel标记。
定位群体中, 具有西农1A带型的单株记为A, 具有wp4带型的单株记为B, 具有西农1A/wp4带型的单株记为H, 用Mapmaker3.0进行数据分析和作图, 用Kosambi函数将重组率转化为遗传距离(用cM表示)。
田间种植条件下, 野生型的叶片全生育期均为绿色, 穗抽出剑叶叶鞘后呈翠绿色, 成熟期则为金
黄色。wp4的叶片分蘖前期为黄绿色(图1-A, B), 抽穗灌浆期则为浅黄色(图1-C, D); 稻穗抽出剑叶叶鞘后, 内外颖呈乳白色(图1-E, F, G)。与突变亲本缙恢10号相比, wp4的株高、穗长、穗粒数和千粒重略有下降, 有效穗和结实率则略有升高, 但差异均未达到显著水平(表1)。
 | 图1 野生型缙恢10号和突变体wp4的表型鉴定A: 分蘖前期缙恢10号(WT)和wp4植株; B: 分蘖前期WT和wp4叶片; C: 灌浆期WT和wp4叶片; D: 灌浆期WT和wp4植株; E: WT和wp4的穗; F: WT和wp4的籽粒; G: WT和wp4的一次枝梗。Fig. 1 Plant morphology of the wild type Jinhui 10 and the mutant wp4A: phenotype of the wild type (WT) and the wp4 at the early tillering stage; B: leaf blades of the WT and the wp4 at the early tillering stage; C: leaf blades of the WT and the wp4 at the filling stage; D: phenotype of the WT and the wp4 at the filling stage; E: panicles of the WT and the wp4 at the flowering stage; F: mature seeds of the WT and the wp4; G: primary rachis branches of the WT and the wp4 at the flowering stage. |
| 表1 野生型(WT)和wp4农艺性状分析 Table 1 Agronomic trait of the wild type (WT) and the wp4 |
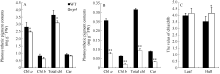
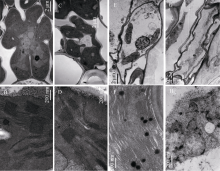
与野生型相比, 叶片中, wp4的叶绿素a含量显著降低, 叶绿素b和类胡萝卜素含量无变化(图2-A), 内外颖中, 叶绿素a、叶绿素b和类胡萝卜素含量均极显著降低(图2-B)。叶片中, 野生型和突变体的Chl a/Chl b值无明显差异; 颖壳中, 由于野生型的Chl a/Chl b值降低, 导致wp4的Chl a/Chl b值显著大于野生型。透射电镜观察发现, 与野生型相比, wp4叶肉细胞内的细胞器结构较为完整、清晰, 叶绿体中含有基粒、基质和基质类囊体, 但基质片层略少且排列较为松散(图3-A, B, C, D)。野生型颖壳中的叶绿体数量少于叶片部位, 内含基粒较薄, 但类囊体发育基本完整, 基质片层排列规则; 突变体wp4中, 颖壳部位的细胞形状不规则且细胞器结构异常, 质体没能发育成含有明显类囊体结构的叶绿体(图3-E, F, G, H)。
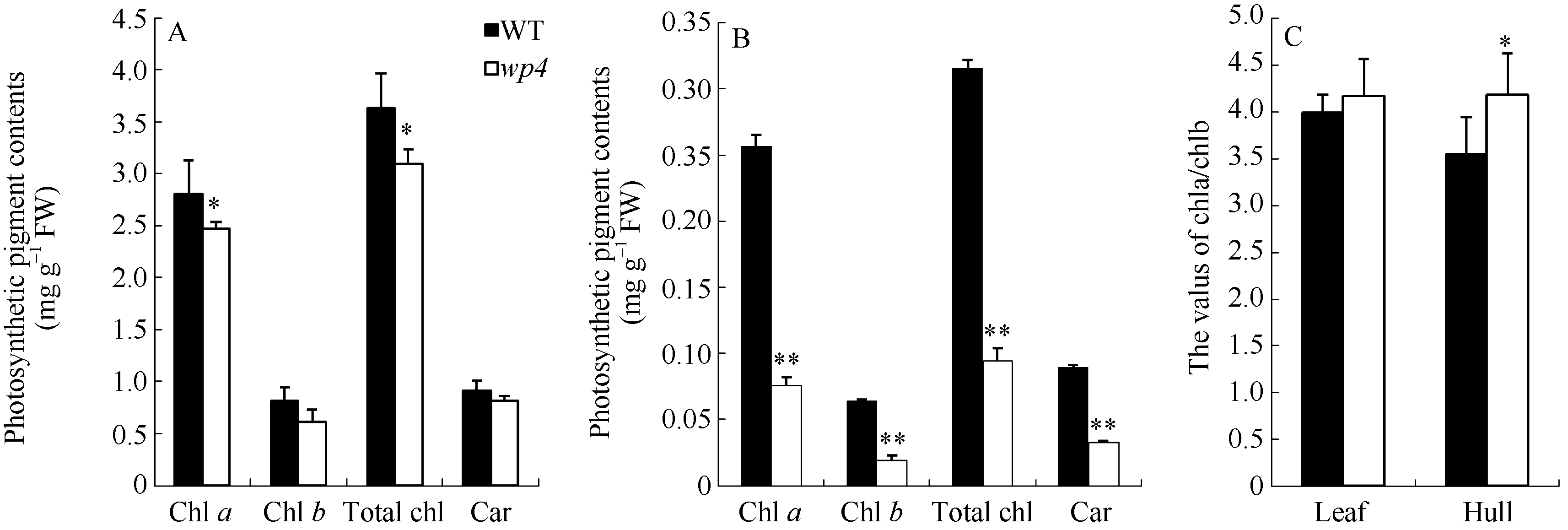 | 图2 开花期野生型(WT)和wp4的光合色素含量A: 叶片光合色素含量; B: 内外颖壳的光合色素含量; C: 叶片和颖壳中的叶绿素a和b的比值。Fig. 2 Photosynthetic pigment contents of the wild type (WT) and the mutant wp4A: photosynthetic pigment contents of the leaf blades in the WT and the wp4; B: photosynthetic pigment contents of the hulls in the WT and the wp4; C: the values of Chl a/Chl b in the leaf and hull. |
 | 图3 野生型缙恢10号和突变体wp4的细胞超微结构观察A: 野生型叶片内的细胞; B: 野生型叶片细胞中的叶绿体; C: wp4叶片内的细胞; D: wp4叶片细胞内的叶绿体; E: 野生型外稃内的细胞; F: 野生型外稃细胞中的叶绿体; G: wp4外稃内的细胞; H: wp4外稃细胞中的质体。Fig. 3 Cell structure of the wild type and mutational leaf blades observed by Transmission Electron MicroscopeA: cell structure of the leaf blade in the wild type; B: chloroplast in the leaf blade of the wild type; C: cell structure of the wp4leaf blade; D: chloroplast of the wp4 leaf blade; E: cell structure in the lemma of the wild type; F: chloroplast in the lemma of the wild type; G: cell structure of the wp4 lemma; H: plasmid of the wp4 lemma. |
西农1A/wp4杂交组合中, F1植株的叶片和稻穗颜色均发育正常; F2群体中, 根据株型颜色可分为两类, 一类与野生型类似, 叶片和稻穗的颜色均正常发育, 一类与wp4的表型相似, 全生育期叶色略淡、抽穗后稻穗呈乳白色或乳黄色。4558株F2群体中, 3438株表型正常, 1120株为突变表型, χ 2测验显示正常与突变单株符合3∶ 1的分离比[χ 2=1.56< χ 2(0.05)=3.84], 表明wp4的突变型受1对隐性核基因控制。
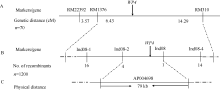
均匀分布在水稻12条染色体上的480对RM系列SSR引物, 112对在西农1A和wp4间呈现多态性, 其中, 位于第8染色体上的RM22392、RM1376和RM310在正常基因池和突变基因池之间也呈现多态性, 紧邻标记同时存在多态性暗示其可能与目的基因连锁。70个典型F2隐性单株检验表明WP4位于RM1376和RM310之间, 遗传距离分别为6.43 cM和14.29 cM (图4-A)。
在初步定位区间内, 设计开发Indel标记, 利用亲本间呈现多态性的标记进行精细定位, 多态性标记的引物序列见表2。结果发现, 1120株F2隐性单株中, Ind08-1、Ind08-2、Ind08-3和Ind08-4的交换株数分别为16、4、1和14个, 且前2个标记与后2个标记的交换株不同, 从而最终将WP4定位在Ind08- 2和Ind08-3之间约79 kb的物理距离内(图4-B, C)。
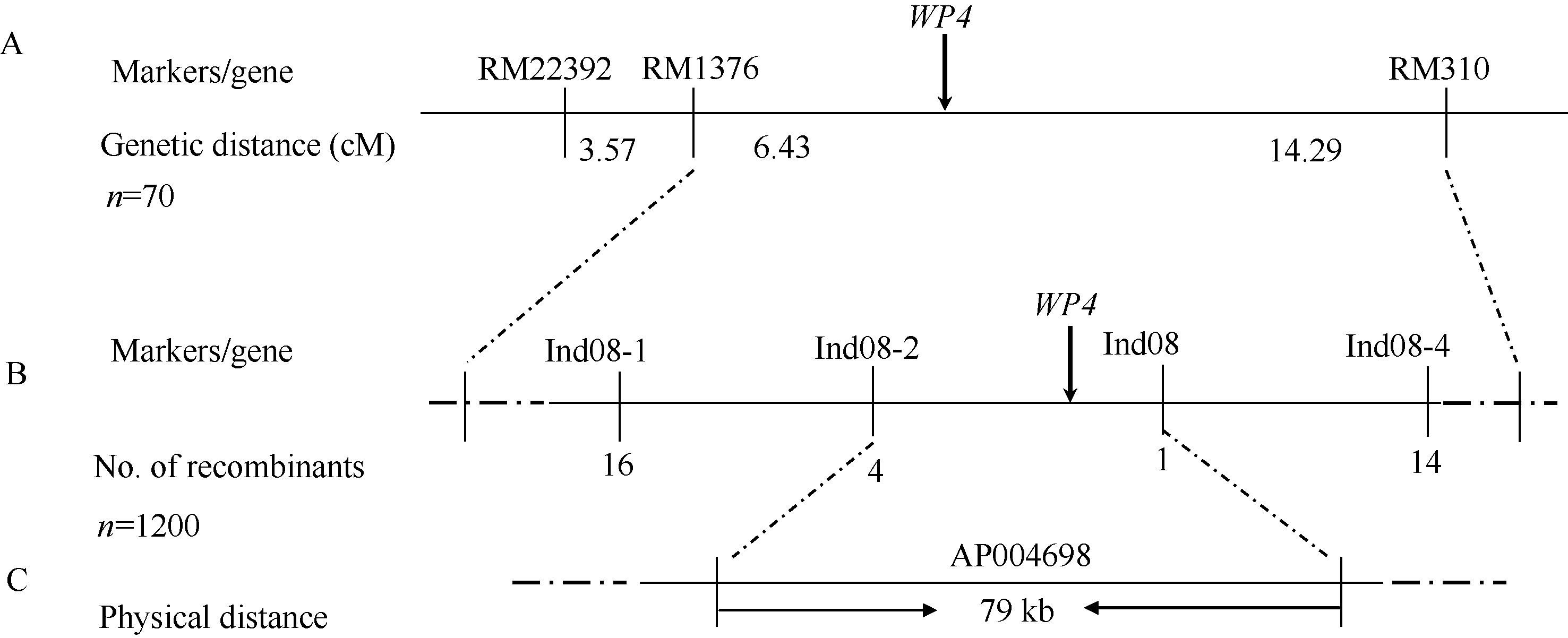 | 图4 WP4在水稻第8染色体上的分子定位Fig. 4 Molecular mapping of WP4 on rice chromosome 8 |
| 表2 WP4精细定位的InDel引物序列 Table 2 Newly designed InDel markers used for mapping the WP4 locus |
根据Gramene和RGAP (水稻基因组注释数据库: Rice Genome Annotation Project)提供的水稻基因组注释序列, 在WP4精细定位的79 kb物理范围内, 含有14个注释基因(表3)。其中, MYB家族转录因子、小檗碱结构域蛋白、表达蛋白和转座子编码基因各有2个, 其余6个分别编码B3 DNA结合结构域蛋白、无顶端分生组织蛋白(NAM)、牛心果碱氧化酶前体、stomatin-like蛋白、转移酶和核仁GTP结合蛋白。其中, 转移酶编码基因含有一个叶绿体信号肽, 我们对其进行了测序, 但野生型和突变体之间没有发现碱基差异。
| 表3 定位区间RGAP注释基因 Table 3 RGAP annotated genes in the restricted region of WP4 locus |
目前, 在水稻中已报道了6个白穗突变体, 分别为wp1、wp2、wp(t)、wslwp、st-fon和wlp1。wp1和wp2是较早报道的白穗突变体, 主要特征是成熟期穗部白色, 叶片出现不同程度的白色条纹, 基因分别位于第7和第1染色体上[6]。wslwp与wp1的表型类似, wp(t)则与wp2的表型一致, 二者的调控基因也被分别定位在第7和第1染色体上, 其可能互为等位突变体[8, 9]。st-fon除穗部白化和叶片出现白色条纹外, 还表现内外颖的异常发育, 对基因尚没有进行染色体定位[10]。wlp1除抽穗期穗部白化外, 四叶期之前叶片白化, 其后转绿[7]。与已报道的白穗突变体相比, wp4的表型明显不同, 穗抽出后呈乳白色, 一直持续到灌浆后期, 全生育期内叶片淡绿, 没有出现白化或白色条纹, 因此, wp4是一个新型白穗突变体。
本研究中将WP4精细定位在水稻第8染色体79 kb的物理范围内, 包含14个注释基因, 其中, 仅LOC_Os08g06220编码蛋白具有叶绿体信号肽。在水稻中, 约4500个核基因编码蛋白具有叶绿体信号肽, 这类蛋白多与叶绿体的发育相关[13]。如水稻花叶白穗突变体wlp1, 调控基因即编码一个靶向于叶绿体的50S核糖体L13蛋白[7]。因此, 我们对LOC_Os08g06220进行了测序, 但野生型和突变体之间没有发现碱基差异, 表明其不是目的基因。核仁GTP结合蛋白存在于细胞核内, 具有高度的保守性, 在真核和原核生物中广泛存在[14]。在酵母中, NOG (nucleolar GTP-binding protein)与TOR (target of rapamycin)互作, 通过营养供应调节60S核糖体前体的成熟[15]。通过线虫、酵母等真核生物的研究, 基本明确了NOG蛋白在真核生物核糖体形成中的关键作用[16, 17]。NbDER (Nicotiana benthamianaDouble Era-like GTPase)含有2个重复的GTP结构域, 参与叶绿体rRNA的剪切和核糖体的生物合成, RNAi导致烟草叶片黄化, 暗示DER在植物生长发育中具有重要的功能[18]。LOC_Os08g06230编码一个核仁GTP结合蛋白NOG1, 但其是否是WP4的目的基因, 以及植物中的NOG是否也具有类似真核生物的功能, 尚需进一步的研究。
目前报道的基因, 多具有一因多效性。如hw-1(t), 三叶期之前白化, 随后转绿, 同时又表现为矮化多蘖, 受单隐性基因调控[19]。突变体wp4除穗部白化外, 另一个典型的特征是叶色黄绿。黄绿叶是水稻一种常见的叶色变异, 目前已经鉴定了10多个相关突变体, 并克隆了YGL1、YGL2、YGL138(t)等调控基因, 这些基因在叶绿素合成和叶绿体发育过程中起重要作用[20, 21, 22]。相对于叶片白化、花叶等性状, 黄绿叶突变体的农艺性状尽管也受到了一定的影响, 但影响较小, 是叶色标记利用的理想材料[23, 24, 25]。本研究发现, wp4叶片尽管呈黄绿色, 但叶绿体结构较为完整, 光合色素中仅叶绿素a的含量下降达显著水平, 农艺性状中, 有效穗和结实率略有增加, 株高、穗长、穗粒数和千粒重则略有下降, 但均未达到统计学上的差异水平。这表明wp4可作为标记性状用于水稻的良种繁殖, 同时也暗示穗部光合作用即使对水稻的产量性状具有一定的影响, 效应值也较小。
白穗突变体wp4全生育期叶色黄绿, 抽穗灌浆期内外颖呈乳白色, 穗轴颜色不变。叶片部位, wp4的光合色素含量明显下降, 但仅叶绿素a与野生型相比在统计学上达显著差异水平, 叶肉细胞的叶绿体除基质片层略少且排列松散外, 结构基本完整。颖壳中, wp4的叶绿素a、叶绿素b和类胡萝卜素的含量均极显著下降, 细胞内无明显的叶绿体结构。突变体的农艺性状与野生型相比, 差异不显著。该突变体受1对隐性核基因控制, 利用1200株西农1A/wp4的F2隐性群体, 最终将WP4精细定位在第8染色体约79 kb的物理范围内, 包含14个注释基因。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|