



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
一份新的水稻斑点叶突变体 spl32的鉴定和基因定位
[钟振泉*  , 罗文龙
, 罗文龙* , 刘永柱, 王慧, 陈志强* , 郭涛* ]
, 罗文龙, 刘永柱, 王慧, 陈志强, 郭涛]
|
|


第一作者联系方式: 钟振泉, E-mail: zhongzhq1990@163.com; 罗文龙, E-mail: lovenlong@163.com
**同等贡献(Contributed equally to this work)
从F2 (粤晶丝苗2号/H4) 群体中, 鉴定出一份显性斑点叶突变体 spl32 ( spotted leaf 32)。其叶片褐色斑点受自然光诱导, 在幼穗分化期从叶尖逐渐扩散至叶鞘, 台盼蓝染色表明斑点并非由细胞死亡引起。以从F5杂合个体分离出的正常叶色植株为对照, 斑点叶植株的穗粒数、结实率显著降低。斑点出现后, spl32的POD活性和MDA含量均显著高于对照; 同时, spl32叶片光合色素含量降低, 但荧光动力学参数并无显著变化。抽穗期人工接菌表明, spl32对水稻白叶枯病菌抗性较对照显著提高。遗传分析表明 spl32斑点性状由一个显性基因 Spl32(t)控制, 利用F2 (02428/ spl32)群体将其定位在第11染色体Ind-c和RM206之间, 推测该基因为一个新的水稻斑点叶基因。
A dominant spotted-leaf mutant of rice was isolated from F2 (Yuejingsimiao 2/H4) population. The mutant, designated as spl32 ( spotted-leaf 32), initiated brown spots on leaf apex at the panicle differentiation period, and then gradually spread them to whole leaf and sheath. Trypan blue staining indicated that the formation of spots was not caused by cell death. Taken normal green leaf plants segregated from heterozygous F5 as control (CK), we found seeds per panicle and seed setting rate of spotted leaf plants were significantly lower than these of CK. After appearance of spots, the POD activity and MDA content of spl32 were significantly higher than these of CK, while photosynthetic pigment content in spl32was reduced, without significant changes in chlorophyll fluorescence parameters. The resistance to rice bacterial blight in spl32was greatly improved by inoculation of Xanthomonas oryzae pv. oryzae at heading period. The spotted-leaf trait of spl32was verified to be controlled by a dominant gene that temporarily designated as Spl32(t). The novel rice spotted-leaf gene was mapped between markers Ind-c and RM206 on chromosome 11 with a F2 (02428/ spl32) population.
植物斑点叶(spotted leaf)突变体是指植物在没有遭受病原物浸染或明显逆境条件下在叶片或叶鞘上自发形成斑点的一类突变体[1, 2]。突变体产生的斑点类似于病原菌侵染时产生的病斑, 因此又称为类病变突变体(lesion mimic mutants, lmm)或类病斑(lesion simulating disease)突变体[3]。除少部分突变体斑点的形成可能由色素积累外, 大部分是由植物本身细胞死亡引起。这类斑点叶突变体的表型多与病原菌侵染后的过敏反应(hypersensitive response, HR)症状类似[4], 而且许多斑点叶突变体对某些植物病原物表现出了一定的抗性[5, 6, 7], 能激发病程相关蛋白的表达[2, 8, 9], 可以提高植株的持久、广谱抗病性。
斑点叶突变体或类病变突变体广泛存在于各种植物, 包括水稻、玉米、大麦和小麦等[6]。到目前为止, 在水稻中至少已经发现了50多个斑点叶突变体[6], 其中spl5[10]、spl7[11]、spl11[12]、spl18[13]、spl28[7]、OsLSD1[14]、NPR1[15]、OsPtila[16]、OsACDR1[17]、OsSSI2[18]、OsSL[19]、RLIN1[20]、RLS1[21]、NLS1[22]、lms1[23]、lm[24]和CHL1[25]等斑点叶或类病变相关基因已经被克隆。这些基因的作用机制各不相同, 且突变体性状大多是由局部细胞死亡引起的。如第一个被克隆的水稻类病变突变基因spl7负向调控细胞死亡[11]; Zeng等[12]克隆到的SPL11蛋白可能在控制植物细胞死亡及防御反应的泛素系统中起作用; Mori等[13]克隆的SPL18编码一个酰基转移酶, 与防卫反应基因PR1和PBZ1的表达相关, 且显著提高对稻瘟病和白叶枯病的抗性; OsLSD1编码一个锌指蛋白, 负向调控细胞死亡[14]; spl5编码1个转录剪接因子3b亚基3 (SF3b3), 与细胞坏死和抗性反应中的RNA成熟前体的剪接相关[10]。可见斑点产生过程非常复杂, 涉及许多生化代谢途径。因此, 这类突变体对植物过敏反应激活机制和植物抗病反应机制的研究具有重要意义。
斑点叶突变体是植物育种中一类重要的抗病种质资源, 尤其是突变表型对产量、品质等重要农艺性状无明显影响时, 有可能在育种中得到直接应用。在大麦中发现的类病变突变体mlo, 对几乎所有已知的白粉病(Erysiphe graminisf. sp. Hordei)生理小种都具有持久抗性[26]。自1979年以来, 带有mlo基因的大麦品种一直保持高抗性, 在欧洲许多国家已经大面积种植[27]。已报道的水稻斑点叶突变体中, 大多数对病原菌抗性出现不同程度增强, 如spl10、spl11、spl17、spl28和spl26等对稻瘟病和白叶枯病表现出广谱抗性[7, 12, 28, 29]。然而, 大部分突变体在类病变往往会伴随细胞坏死、植株矮小、产量减少等不良的农艺性状, 如spl30、spl31等会导致结实率和千粒重降低[30, 31], spl28、RLS1、lms1等会引起植株早衰[13, 21, 23], 这限制了它们在水稻育种实践中的直接应用。因此, 不断挖掘和鉴定新的水稻斑点叶突变体, 对水稻抗病机制研究和抗病育种实践都有重要意义。
从2个正常绿叶品种籼稻粤晶丝苗2号和H4的F2群体中, 我们鉴定出了一份新的斑点叶突变体, 暂命名为spl32(spotted leaf 32)。通过连续2年4代的观察, 在田间种植条件下, spl32的突变表型能够稳定遗传。本研究从形态学、生理生化、遗传机制等方面对spl32开展研究, 以期为突变基因的克隆、功能分析及应用奠定基础。
斑点叶突变体spl32的2个亲本中, 粤晶丝苗2号系广东省农业科学院水稻研究所选育的优质、高产、抗病水稻品种, H4是本实验室从中二软占的空间诱变后代中获得的水稻高抗稻瘟病株系。
以从斑点叶突变体F5杂合个体分离出的正常叶色植株为对照, 用于农艺性状、酶活性及抗病性的对比; 以粳稻日本晴和02428分别与spl32杂交, 构建遗传分析和基因定位的F2群体。
2013年晚季和2014年早季, 将突变体spl32和对照种植于华南农业大学科研教学实验基地, 小区种植、管理与一般大田相同。全生育期观察田间突变体的表型, 包括突变体斑点出现的时期、颜色及分布情况等。成熟后随机选取10株, 考查株高、单株穗重、每株有效穗数、每穗粒数、每穗实粒数、结实率和千粒重等。
通过遮光试验[32]了解光照对斑点发生的影响。在大田条件下, 用宽度约为1 cm的锡箔纸对突变体spl32叶片已有斑点和无斑点部位进行遮光处理, 7 d后观察斑点发生及变化情况。
在突变体spl32出现斑点后, 取抽穗期突变体spl32植株上表现斑点的剑叶及其对照相同部位叶片进行台盼蓝染色[33]。将叶片浸泡在70℃的台盼蓝溶液中, 缓慢真空渗透5 min, 浸泡10 min, 然后加热沸腾2 min, 室温放置12 h后于2.5 g mL-1的水合氯醛中脱色3~4 d, 用体视镜观察细胞死亡情况并照相记录。
分别测定突变体spl32和对照在分蘖期、抽穗期、灌浆期和成熟期的超氧化物歧化酶(superoxide dismutase, SOD)、过氧化物酶(peroxidase, POD)、过氧化氢酶(catalase, CAT)活性和丙二醛(malondialdehyde, MDA)的含量。
SOD活性采用SOD (WST-1法)测试盒(南京建成生物工程研究所生产)测定, 该试剂盒采用黄嘌呤氧化酶法; POD活性采用愈创木酚法[34]测定, 以每分钟内OD470nm变化0.01为一个过氧化物酶活性单位(U); CAT活性采用高锰酸钾滴定法[35]测定; MDA含量采用硫代巴比妥酸(thiobarbituric acid, TBA)法[36]测定。
采用95%丙酮-乙醇法[37]分别测定突变体spl32及其对照抽穗期、扬花期和成熟期的叶片光合色素含量。将0.2 g待测样品剪碎, 浸泡于25 mL的95%的丙酮∶ 乙醇=2∶ 1的水溶液中, 黑暗环境浸泡48 h, 利用PE分光光度计测定浸提液的光吸收值, 3次重复。按Lichtenthaler等[38]修正的Arnon法计算叶片叶绿素含量。
测定突变体spl32和对照在抽穗期、扬花期、灌浆期和成熟期的叶绿素荧光动力学参数。随机选取长势相对一致的单株各7株, 利用MINI-PAM型便携式光合作用测定仪在凌晨时分测定剑叶固定部位。荧光程序设置为暗处理30 min, 光强为6000 μ mol m-2 s-1饱和光处理45 s。光系统II (PSII)最大量子产量Fv/Fm = (Fm-Fo)/Fm; PSII的实际量子产量∆ F/Fm′ = (Fm′ -F)/Fm′ ; 光化学猝灭系数qP= (Fm′ -Fs)/ (Fm′ -Fo′ ); 非光化学淬灭NPQ = (Fm-Fm′ )/Fm′ 。
用水稻白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)分别接种突变体spl32、对照和F2 (spl32/日本晴)及F2 (02428/spl32)群体。在田间条件下, 选取播种60 d后的水稻单株, 每个单株选3张全展新叶, 剪叶接种。其中对照和突变体分别接种全展倒一叶、倒二叶和倒三叶, 3组重复; 2个F2群体分别接种有斑点植株和正常绿叶植株各50株, 3周后测量病斑长度, 并统计分析。
2013年早季, 以spl32为母本、日本晴为父本, 以及以02428为母本、spl32为父本, 各配制杂交组合获得相应F1种子。2013年晚季种植F1, 2014年早季种植F2, 利用F2分离群体进行斑点叶性状的遗传分析和基因初步定位。
采用Michelmore等[39]提出的近等基因池分析法定位目标基因, 即根据F2表型, 分别选取15株正常植株和15株突变植株, 剪取等量叶片, 构建正常基因池和突变基因池。按CTAB法[40]提取亲本、基因池和F2群体DNA。参照http://www.gramene.org/ microsat/SSR引物序列, 根据粳稻日本晴和籼稻93-11的序列差异比对设计Indel引物, 并由上海生工生物工程有限公司合成。PCR扩增体系含2× PCR Reaction Mix 10 µ L (含100 mmol L-1 KCl、20 mmol L-1 Tris-HCl、3 mmol L-1 MgCl2、400 mmol L-1 dNTP)、Taq DNA聚合酶(5 U μ L-1) 0.2 µ L、引物(10 μ mol L-1)各1 µ L、模板DNA 1 µ L, 超纯水补至20 μ L。反应条件为94℃ 5 min; 94℃ 30 s, 55℃ 30 s, 72℃ 30 s, 35个循环; 72℃ 7 min。PCR产物经8%非变性聚丙烯酰胺凝胶电泳及银染显色后读胶。
F2定位群体中, 将具有spl32突变亲本带型的单株记为A, 具有02428带型的单株记为B, 具有杂合带型的单株记为H。根据公式[(H+2A)/2n]× 100计算遗传距离并构建连锁图谱, 其中H表示定位群体中出现杂合带型单株的数量, A表示出现spl32突变带型的单株数, n表示用于定位的隐性群体总株数。
田间生长情况下, 早季播种约45 d后(幼穗分化期) 突变体spl32开始在叶尖出现褐色斑点, 此后斑点由叶边缘向叶片内部扩散, 呈连续状分布; 灌浆后在叶鞘也逐渐出现斑点(图1)。spl32的斑点从幼穗分化期开始持续到成熟, 属于全生育期扩散型斑点叶突变体。
田间观察发现, spl32在成熟期没有明显的早衰现象。晚季农艺性状分析表明, 与对照相比, spl32突变体在株高、单株穗重和千粒重等方面无明显差异; 但每穗粒数显著低于对照, 每穗实粒数和结实率极显著低于对照(表1)。
利用均匀分布于12条染色体上的617个SSR标记分析spl32及其双亲粤晶丝苗2号和H4的DNA多态性, 只有2个标记在spl32与双亲间存在多态性, 其余位点均与至少一个亲本的条带大小相同。证明斑点叶突变体spl32的确是来源于粤晶丝苗2号和H4, 很可能是自然突变产生的新种质。
在自大田条件下, 分蘖盛期无斑点的新叶经遮光7 d后, 除遮光部位外, 其余部位正常产生褐色斑点; 去除锡箔纸7 d后, 遮光部位重新出现斑点。对于已经出现斑点的叶片部位, 经遮光处理后, 斑点没有消失(图2)。由此可知, spl32突变体的斑点产生受自然光的诱导。
 | 图1 斑点叶突变体spl32的表型A: 灌浆期spl32的田间表型; B: 对照(左)和spl32 (右)乳熟期的植株表型; C: 对照(左)和spl32 (右)灌浆期叶片; D: 成熟期spl32的叶鞘; E: 灌浆期spl32的叶片和穗部。Fig. 1 Phenotypes of spl32A: field phenotype of spl32during the filling stage; B: phenotypes of CK (left) and spl32 (right) during the milky stage; C: leaves of CK (left) and spl32 (right) during the filling stage; D: sheath of spl32during the mature stage; E: phenotypes of leaf and panicle during the filling stage in spl32. |
| 表1 对照(CK)和spl32的农艺性状分析 Table 1 Agronomic traits of the control (CK) and spl32 |
 | 图2 遮光对突变体spl32叶片的影响A: 对照; B: 成熟期spl32叶片; C: 未发生斑点部位遮光7 d后; D: 遮光部位恢复光照7 d后; E: 已有斑点部位遮光7 d后。Fig. 2 Effects of shading on spl32leavesA: CK; B: spl32 during the maturity stage; C: part of non-spotted leaf after shading for seven days; D: leaf shaded for seven days then under normal light for seven days; E: spotted leaf after shading for seven days. |
经台盼蓝染色后的对照叶片(图3-B)和spl32叶片(图3-D)斑点部位均未出现深蓝色, 表明突变体斑点部位不存在细胞坏死。对照植株叶片和突变体spl32叶片上虽然没有深蓝色斑但有蓝色点分布, 这些点在整张叶片上分布比较均匀, 这可能是水稻植株在正常生长过程中的细胞更新。
 | 图3 spl32突变体的组织化学分析A: 对照叶片; B: 对照叶片染色后; C: 突变体spl32斑点叶片; D: 突变体spl32叶片染色后。Fig. 3 Histochemical analysis of spl32A: control; B: control stained; C: spottedspl32 leaf; D: stained spl32 leaf. |
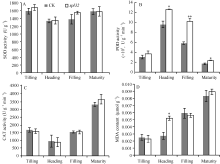
突变体spl32和对照在4个时期的SOD和CAT活力均无显著差异(图4-A和C); 而POD活力在抽穗期和成熟期均显著高于对照, 在灌浆期达到极显著水平(图4-B), 由此推测突变体植株体内可能积累了大量的H2O2。此外, spl32的MDA含量在分蘖期、灌浆期和成熟期与对照无显著差异, 但在抽穗期显著高于对照(图4-D); 说明spl32在抽穗期积累了更多的H2O2, 导致膜系统受损程度高于对照。推测斑点叶突变体spl32比对照有更强的抗氧化能力。
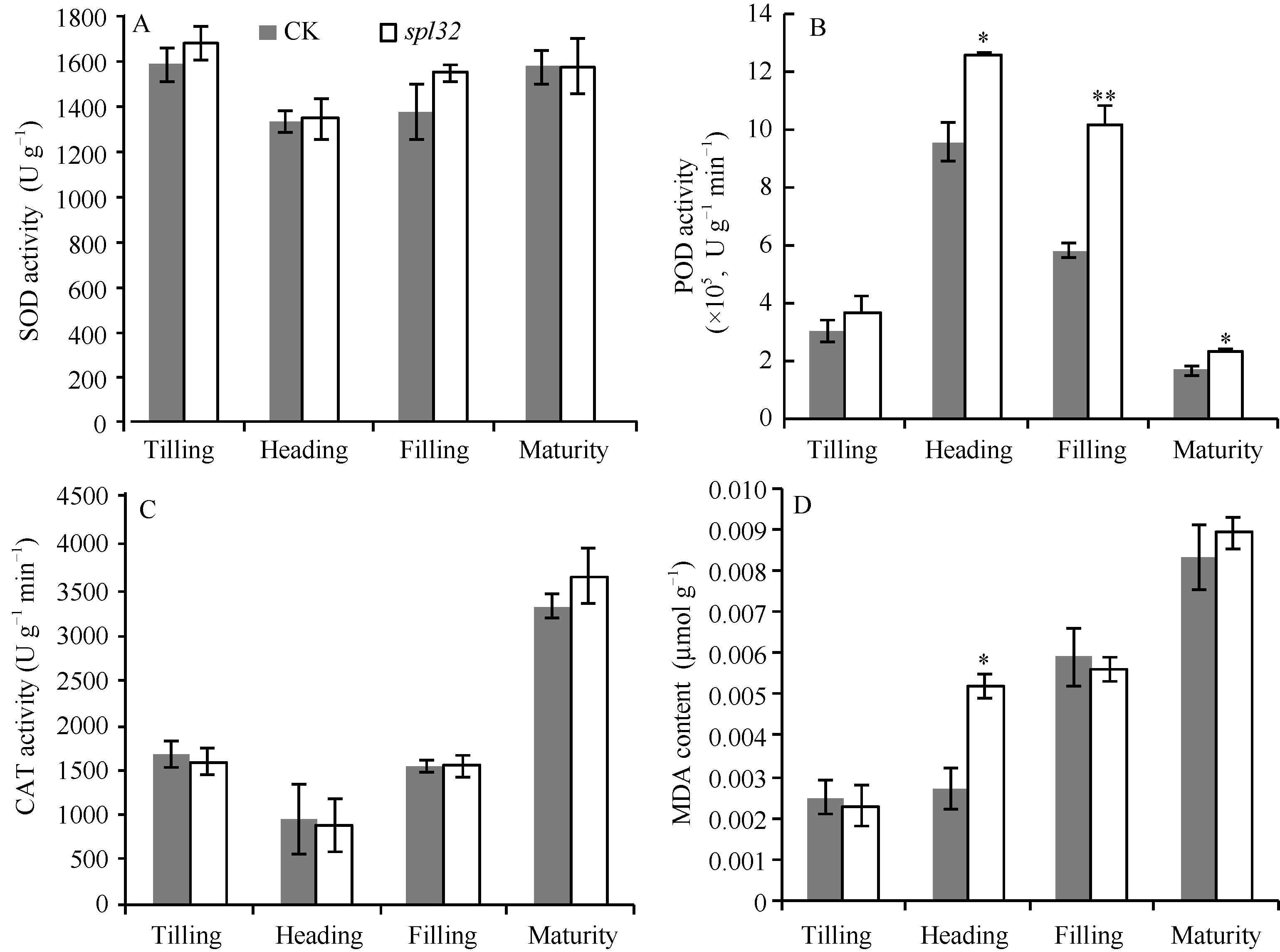 | 图4 突变体spl32与对照(CK)各时期的生理学特性比较A: 超氧化物歧化酶(SOD)(U g-1); B: 过氧化物酶(POD)(U min-1 g-1); C: 过氧化氢酶(CAT)(µ mol g-1 min-1); D: 丙二醛(MDA) (µ mol g-1); * 在 0.05水平上差异显著; * * 在0.01水平上差异显著。Fig. 4 Physiological characteristics of the control (CK) and the spl32 mutant during different growth stagesA: superoxide dismutase (SOD) (U g-1); B: peroxidase (POD) (U min-1g-1); C: catalase (CAT) (µ mol g-1min-1); D: malondialdehyde (MDA) (μ mol g-1). * Significantly different at P< 0.05; * * significantly different at P< 0.01. |
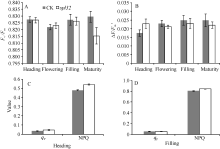
在抽穗期(斑点快速扩散), 突变体和对照各色素指标差异不大; 在扬花期(斑点已大量形成), spl32的叶绿素、类胡萝卜素、光合色素总量和Chl a/Chl b都极显著低于对照; 至成熟期, 突变体的叶绿素a和总叶绿素含量显著低于对照, Chl a/Chl b极显著低于对照(图5)。由此可知, spl32斑点的出现可能导致突变体光合色素含量降低。
图6表明spl32和对照在4个时期的Fv/Fm、∆ F/F′ 差异都不显著, 说明spl32植株叶片的PSII反应中心的能量捕捉效率和实际光合效率与对照无差异, 光能转化正常。spl32和对照在抽穗期和灌浆期的qP和NPQ均差异不显著, 说明其光合活性和光保护能力并无显著差异。由此可知, 突变体spl32斑点症状的出现, 并不会显著影响植株的光合作用。这可能是突变体在成熟期没有出现早衰的原因之一。
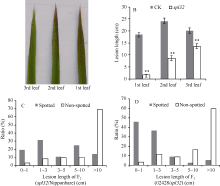
如图7-B, spl32全展倒一叶、倒二叶和倒三叶的病斑长度均小于对照, 其中倒一叶和倒二叶达到极显著水平, 说明斑点症状的出现显著提高了植株对白叶枯病的抗性。抗性差异源于不同功能叶上斑点面积不同。接菌时斑点面积为倒一叶> 倒二叶> 倒三叶(图7-A), 而感病程度为倒一叶< 倒二叶< 倒三叶, 说明植株抗病性与叶片斑点多少有关。
图7表明, 斑点叶个体对白叶枯病病菌的抗性明显高于正常绿叶个体。其中, F2(spl32/日本晴)中斑点叶个体的病斑平均长度2.28 cm, 极显著低于正常绿叶的12.49 cm; 约85%的斑点叶植株的病斑小于3 cm, 而超过60%的正常绿叶植株的病斑长度大于10 cm (图7-C)。在F2(02428/spl32)群体中也得出类似的结果, 斑点叶植株的病斑平均长度4.69 cm, 极显著低于正常绿叶植株的14.63 cm (图7-D)。此外, 2个F2群体中斑点叶植株的病斑长度也有所不同, 这可能与两个群体的生育期差异所致。F2(02428/spl32)群体的抽穗期要比F2(spl32/日本晴)群体的迟, 在接种时F2(02428/spl32)中斑点叶个体上的斑点面积也小于后者, 因此其斑点叶个体对白叶枯病的抗性相对低于后者。由于粤晶丝苗2号、H4、日本晴和02428都对白叶枯病表现高感, 病斑长度均大于10 cm; 推测突变体的白叶枯病抗性是由斑点变异引起的, 即两者呈共分离。
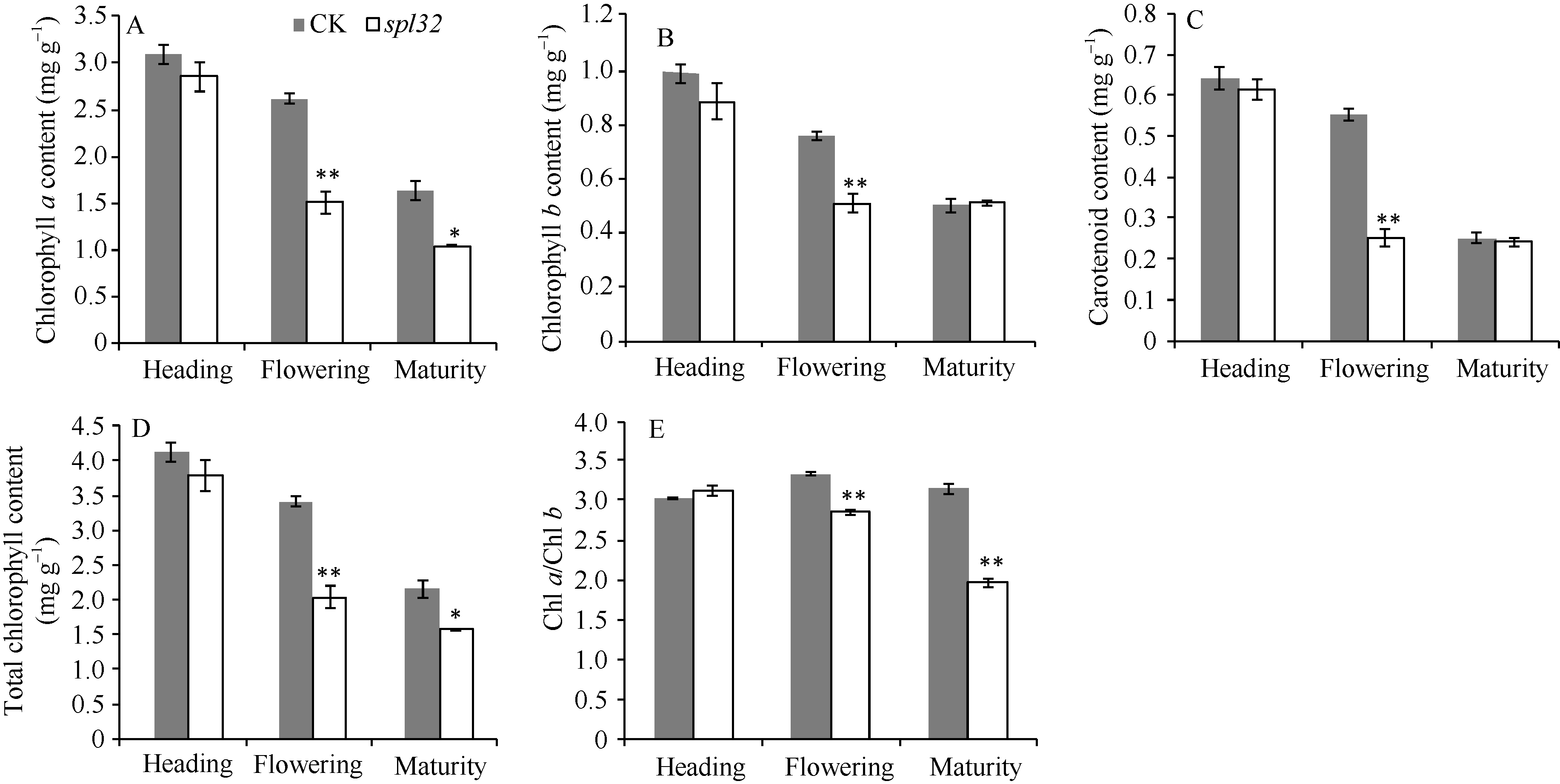 | 图5 突变体spl32和对照(CK)各时期叶片光合色素含量分析A: 叶绿素a含量(mg g-1); B: 叶绿素b含量(mg g-1); C: 类胡萝卜素含量(mg g-1); D: 总叶绿素含量(mg g-1); E: Chl a/Chlb; * 在0.05水平上差异显著; * * 在0.01水平上差异显著。Fig. 5 Pigment content in leaves of spl32 and control (CK) during different growth stagesA: chlorophyll a content (mg g-1); B: chlorophyll b content (mg g-1); C: carotenoid content (mg g-1); D: total chlorophyll content (mg g-1); E: Chl a/Chl b. * Significantly different at P< 0.05; * * significantly different at P< 0.01. |
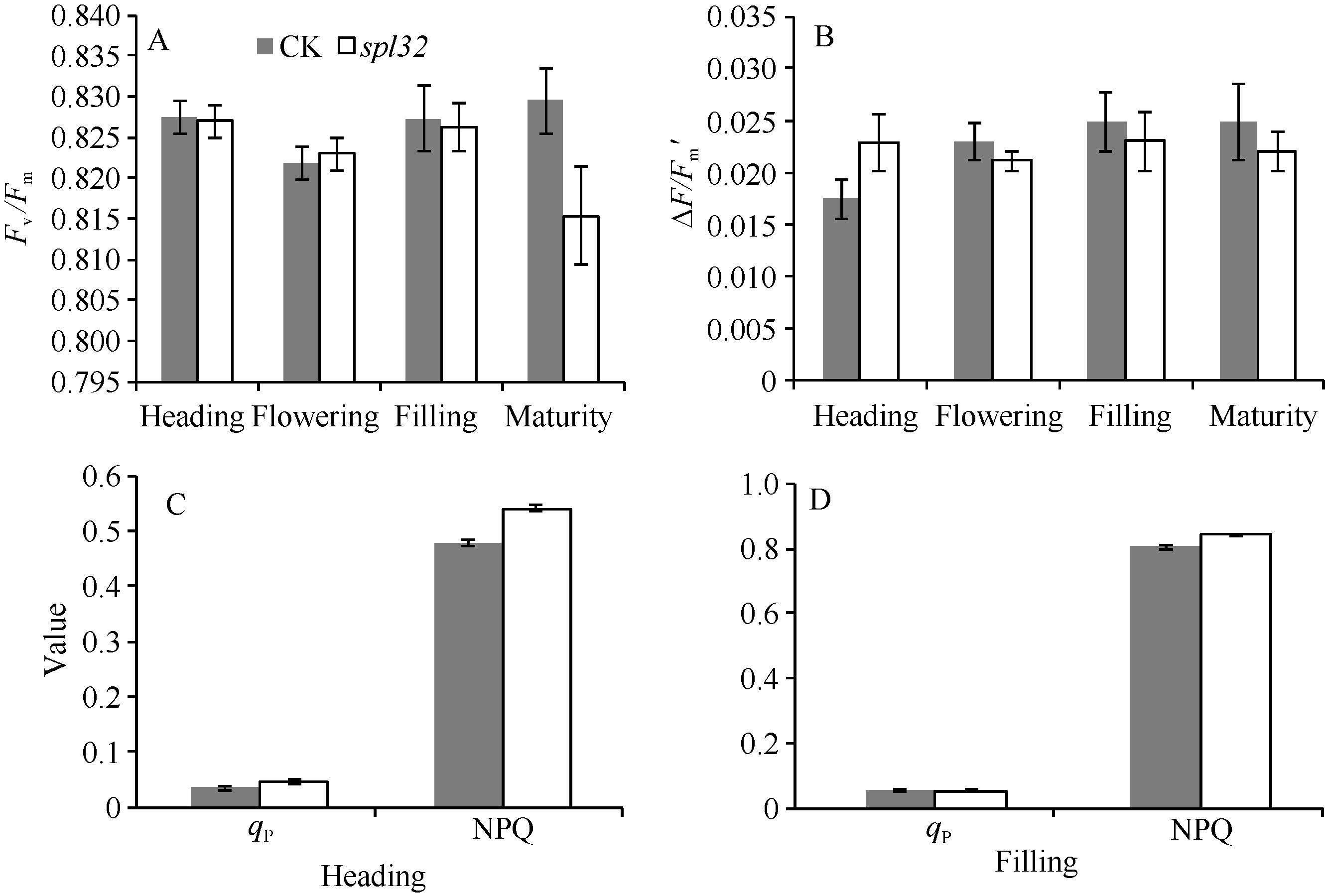 | 图6 突变体spl32与对照(CK)的叶绿素荧光动力学参数比较Fig. 6 Chlorophyll fluorescence kinetic parameters of the spl32 mutant and control (CK) |
A: CK和spl32在各时期叶片的Fv/Fm比较; B: CK和spl32在各时期叶片的∆ F/Fm'比较; C: 抽穗期CK和spl32的qP和NPQ值比较;
D: 灌浆期CK和spl32的qP和NPQ值比较。
A: Fv/Fm of spl32 and check during different growth stages; B: ∆ F/Fm' of spl32 and check during different growth stages; C: the qP and NPQ of spl32and check during heading stage; D: the qP and NPQ of spl32 and check during filling stage.
 | 图7 斑点叶性状与白叶枯病抗性的相关性分析A: 接菌时3片功能叶的斑点情况; B: 3片功能叶的病斑调查; C: F2(spl32/日本晴); D: F2(02428/spl32)。Fig. 7 Association analysis of the spotted-leaf trait and bacterial blight resistance |
* 在0.05水平上差异显著; * * 在0.01水平上差异显著。
A: spots on the three functional leaves when inoculated; B: lesion length of the three functional leaves in CK and Spl32(cm);
C: F2(spl32/Nipponbare); D: F2 (02428/spl32). * Significantly different at P< 0.05; * * Significantly different at P< 0.01.
spl32分别与日本晴和02428配制的F1均表现为斑点叶表型, 说明突变表型由显性基因控制。分别统计F2(02428/spl32)群体和F2(spl32/日本晴)群体中的斑点叶和正常绿叶个体数, 调查其分离比例(表2)表明, 该斑点叶性状受1个显性核基因控制, 暂命名为Spl32(t)。
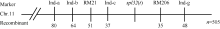
选用02428× spl32杂交的F2群体为定位群体, 共获得505个F2隐性单株(即正常叶表型)。利用本实验室均匀分布于12条染色体上的617对SSR标记对亲本spl32和02428进行多态性分析, 其中共185个标记表现出明显的多态性, 多态率为30.08%。进一步利用在两亲本间表现出多态的引物, 扩增正常基因池和突变基因池, 发现第11染色体的标记RM21和RM206与突变体表型有明显的连锁。在定位区间周围, 进一步开发Indel分子标记13个, 其中4个有多态性(表3)。利用RM21、RM206以及4个Indel标记对505个正常绿叶的F2单株进行分析, 结果标记RM21、Ind-c、RM206和Ind-g分别有51、37、35及48个交换株, RM21交换株包含Ind-c交换株, Ind-g交换株包含RM206交换株, 且RM21、Ind-c交换株与RM206、Ind-g交换株互不重叠, 因此将Spl32(t)基因初步定位在Indel标记Ind-c与SSR标记
| 表2 斑点叶突变体spl32的遗传分析 Table 2 Genetic analysis of spotted leaf mutant spl32 |
RM206之间, 与2个标记间的遗传距离分别为3.4 cM和3.2 cM (图8)。该区间内没有已知可产生斑点叶表型的基因, 推测Spl32(t)是一个新的斑点叶基因。进一步的定位工作正在进行中。
| 表3 定位基因Spl32(t)所用的4个Indel引物序列 Table 3 Four Indel primer sequences used for Spl32(t) mapping |
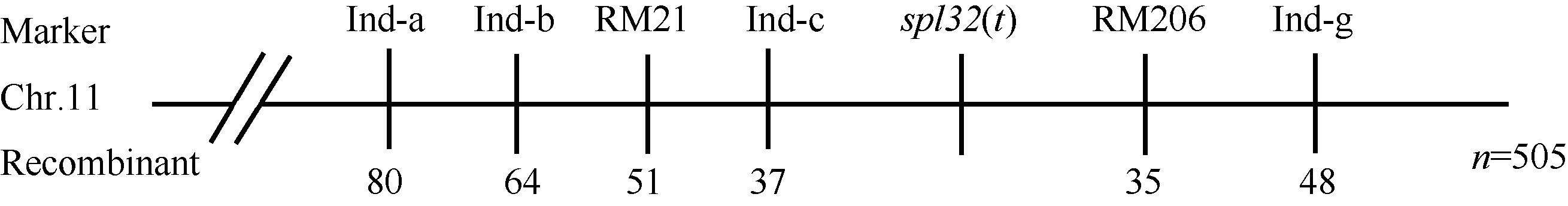 | 图8 突变体基因Spl32(t)的初步定位Fig. 8 Primary mapping of Spl32(t) |
在植物中已发现的大量斑点叶或类病斑突变体中, 斑点或者类病斑的出现往往还伴随着其他农艺性状的改变。如突变体spl28和RLS1基因会引起植株早衰[7, 21]。HM47突变体株高、分蘖数、穗长、每穗粒数、结实率和千粒重都显著低于其野生型[41]。本研究的斑点叶突变体spl32从幼穗分化期开始出现褐色斑点, 属扩散型斑点叶突变体。与对照相比, 突变体spl32的每穗粒数显著降低, 每穗实粒数和结实率极显著降低。梁颖等[42]研究表明, 叶绿素含量是影响油菜生物产量的主导因素。本研究中突变体光合色素含量在斑点出现后极显著降低, 说明斑点的出现影响了突变体色素合成, 进而影响植株光合产物的积累, 这可能是引起突变体每穗实粒数和结实率下降的主要原因。斑点出现后的叶绿素荧光动力学参数无明显变化, 说明Spl32(t)基因突变对植株光合作用途径影响不大, 这也可能是整体农艺性状下降幅度不显著的原因之一。
斑点叶形成的原因很复杂, 它是由植物自身的生理原因和外界环境中的众多因素共同作用决定的[43]。大部分斑点或者类病斑的出现是由于植物本身细胞死亡引起的, 如Frye等在番茄过量表达Pto基因导致了斑点叶的产生[44]; 水稻斑点叶突变体spl5是因为活性氧产生的途径被突变而加强, 使得细胞内O2܋
大量积累, 导致叶片上形成斑点性状[45]。部分突变体斑点的形成则推测是色素的积累。Huang等[30]研究表明斑点叶突变体spl30的红棕色斑点为某种未知色素积累造成的。本研究鉴定的spl32经台盼蓝染色, 未发现有细胞死亡的情况, 说明突变体斑点的产生不是由于细胞死亡引起的, 具体原因还有待进一步的研究。同时, 本研究中的突变体spl32斑点表型的出现受自然光照的诱导, 且其斑点形成信号不能系统传递。
许多斑点叶突变体出现也伴随抗逆性和抗病性的增强。虞锦玲等[46]研究一份经日本晴诱变的类病斑突变体发现, 突变体植株的SOD、POD、CAT活性和MDA含量均明显高于野生型植株, 说明类病变的出现增强了突变体的抗逆性。一些水稻斑点叶突变体如spl10、spl11、spl17、spl28和spl26等对稻瘟病和白叶枯病表现出广谱抗性[7, 12, 28, 29]。本研究发现突变体spl32的SOD和POD活力均高于对照, 说明突变体较对照有更强的抗氧化能力, 抗逆性更强。同时本研究中突变体spl32对白叶枯病抗性极显著增强, 且白叶枯病抗性与斑点共分离。水稻中虽然发现了许多抗病基因, 且其中一些基因如Xa21、xa5、Xa7、xa13、Pi-zt、Pi-ks、Pi-sh等[47, 48]已经运用到抗病育种中去, 但是这些基因存在小种专一化和抗性不持久等问题。突变体spl32斑点的出现不会
引起植株的早衰, 不显著影响植株的光合作用, 且可以提高植株的白叶枯病抗性, 激活植株的整体抗性, 是水稻抗病育种潜在的种质资源, 对植物抗病机理及植物抗病育种等相关研究具有重要意义。
迄今, 通过分子标记定位的几十个水稻类病变基因分布于除第9染色体以外的其他11条染色体。其中在第11染色体上鉴定了2个斑点叶相关的基因。一个是叶鞘坏死基因nls1, 被定位于第11染色体短臂, 已经被克隆, nls1突变体在叶鞘出现自发坏死斑。nls1编码一个典型的CC-NB-LRR类R蛋白, 其功能获得性突变导致了突变体特定组织细胞的自主性死亡[22]。另一个是类病斑及早衰基因lms1[49], 被定位于第11染色体长臂末端, 其突变体植株在叶片和茎部出现黄褐色小斑点, 抽穗期后呈现穗、茎、叶明显干枯, 并快速衰亡。其性状受单隐性核基因控制。本研究的突变体其斑点性状首先出现在叶片上, 受单显性核基因Spl32(t)控制, 初步定位于第11染色体长臂标记Ind-c和RM206之间, 该区间内没有已知可造成类似表型的基因, 因此Spl32(t)是一个新的斑点叶基因。
突变体spl32的褐色斑点可能是由于未知色素积累而形成, 对此类非细胞死亡引起的斑点叶突变体的相关研究还很少。本研究从形态学、生理生化、分子遗传学等方面对spl32开展研究, 为Spl32(t)的克隆和功能验证奠定了基础, 对研究植物斑点叶的发生机制具有重要意义, 也为水稻抗病种质资源的创制提供了新的思路。
水稻斑点叶突变体spl32在幼穗分化期开始在叶尖出现褐色斑点, 此后斑点由叶边缘向叶片内部扩散, 直至遍布整个叶片; 其斑点的出现受自然光诱导, 但并非由细胞死亡引起。斑点的出现会降低其光合色素含量, 但对荧光动力学参数并无明显影响, 不影响植株的PSII。与对照相比, 穗粒数和结实率等部分农艺性状显著下降, 抗氧化能力增强; 对白叶枯病的抗性增强, 且白叶枯病抗性与斑点共分离。该性状受1个显性基因Spl32(t)控制, 位于第11染色体Ind-c和RM206之间。本研究结果为Spl32(t)基因的进一步克隆和功能分析奠定了基础。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|