
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
玉米与马铃薯间作对根际微生物群落结构和多样性的影响
[覃潇敏1  , 郑毅
, 郑毅1, 2, * , 汤利1 , 龙光强1 ]
, 郑毅, 汤利|
|


第一作者联系方式: E-mail: 18314583461@163.com, Tel: 18314583461
设置3个种植处理(玉米单作、马铃薯单作和玉米马铃薯间作), 通过田间试验和盆栽试验, 采用Biolog技术分析了玉米马铃薯根际微生物的群落结构和功能多样性的变化情况。结果表明, 与单作相比, 间作处理使玉米和马铃薯根际微生物对31种碳源的平均利用率(AWCD)分别增加17.36%、7.38%和3.76%、32.21%, 其中间作马铃薯的AWCD值最高, 单作玉米最低。间作玉米和间作马铃薯根际微生物群落的Shannon指数( H)、Simpson指数( D)、均匀度指数( E)、丰富度指数( S)均高于单作, 但在间作和单作之间差异不显著。根际微生物对6类碳源的利用强度以糖类、羧酸类和氨基酸最高, 并存在一定的差异。主成分和聚类分析表明, 玉米与马铃薯间作改变了根际微生物的群落组成, 糖类、羧酸类和氨基酸类碳源是区分单间作处理差异的敏感碳源。说明玉米马铃薯间作改变了根际微生物群落结构组成, 提高了根际微生物群落功能多样性。
Field and pot experiments were conducted with three planting treatments (maize monocropping, potato monocropping, maize, and potato intercropping) to investigate the changes of composition and function diversity of rhizosphere microbial community of maize and potato using Biolog technique. The results indicated that compared with monocropping, the average well color development (AWCD, utilization rate of 31 carbon sources) of rhizosphere microbes of intercropped maize and potato was increased by 17.36%, 7.38% and 3.76%, 32.21% respectively, among the intercropping and monocropping crops, the AWCD value of intercropped potato was the highest and the monocropped maize was the lowest. Functional diversity index of Shannon index ( H), Simpson index ( D), Evenness index ( E), and Richness index ( S) were higher in intercropping rhizosphere soil than these in the monocropping systems, but no significant difference. Among six types of carbon sources used by the rhizospheric microbes, carbohydrates, amino acids and carboxylic acids, had higher utilization rates with definite differences. Principal component analysis (PCA) and cluster analysis demonstrated that the amino acids, carbohydrates and carboxylic acids were the sensitive carbon sources for differentiating the changes of the microbial community induced by monocropping and intercropping. This study showed that intercropping can change the composition and enhance functional diversity of the microbial community in the rhizosphere of maize and potato.
土壤微生物是土壤生态系统的重要组分, 主导土壤生态系统养分循环和能量流动, 对于维持系统的稳定性和可持续性具有重要的作用[1]。微生物群落结构或微生物多样性与土壤可持续利用密切相关, 可作为生物指标指示土壤质量、评价土壤肥力[2]。其中, 土壤微生物功能多样性, 与土壤C、N、P等元素循环转化的生物化学过程之间的关系非常密切[3]。因此, 土壤微生物群落功能多样性是描述土壤微生物群落特征的一个重要指标, 反映了土壤中微生物的生态特征和土壤状况的重要信息。土壤微生物多样性的研究方法很多, 有关用C素的微平板测定方法(Biolog)是测定土壤微生物对不同碳源利用能力及其代谢差异, 进而用以表征土壤微生物功能多样性的一种方法, 目前已经广泛应用于土壤微生物群落功能多样性的研究[4, 5, 6]。
间套作作为我国传统农业的精髓, 具有增产、提高养分资源利用效率[7]、增加农田生物多样性和稳定性、持续控制病虫草害的优势, 并且改善农田生态环境, 促进生态平衡[8]。已有研究表明, 间作能改变根际土壤微生物群落多样性, 提高产量[9], 增加作物对磷的吸收[10]。玉米是我国主要的粮食作物, 马铃薯也是西南地区主要的粮食、经济作物, 玉米与马铃薯间作是一种代表高、矮秆作物的典型配置, 能发挥作物空间的互补作用, 在云南地区普遍种植。前期研究表明, 玉米马铃薯间作可以增加根际微生物的数量[11], 但二者间作对根际微生物群落功能多样性的影响的研究还未见报道。为此, 本研究通过田间和盆栽试验, 采用Biolog技术探讨玉米马铃薯间作根际微生物群落功能多样性, 旨在为玉米马铃薯间作这一种植模式的优质高产提供更深入的微生物学依据。
田间试验地位于云南农业大学寻甸大河桥农场, 地理坐标为23° 32′ N、103° 13′ E, 海拔1953.5 m, 地处昆明市东北部, 属北亚热带季风气候, 年平均气温14.7℃, 年降水量1040 mm。土壤含有机质25.06 g kg-1、碱解氮87.37 mg kg-1、速效磷23.31 mg kg-1、速效钾207.82 mg kg-1、pH 6.79。
盆栽试验在云南农业大学资源与环境学院植物营养系温室大棚进行, 供试土壤为云南农业大学后山旱地红壤。土壤含有机质11.76 g kg-1、碱解氮75.77 mg kg-1、速效磷14.22 mg kg-1、速效钾113.48 mg kg-1、pH 6.95。
供试玉米(Zea maysL.)品种为寻单7号; 马铃薯(Solanum tuberosumL.)品种为会-2。
1.1.1 试验设计 田间试验采用随机区组排列, 设玉米单作、马铃薯单作及玉米马铃薯间作(玉米和马铃薯各2行) 3个种植处理, 每个处理3个重复, 共9个小区, 每小区面积32.50 m2 (5.0 m× 6.5 m)。玉米行距0.5 m、株距0.25 m; 马铃薯行距0.5 m、株距0.35 m, 间作小区中玉米马铃薯的种植密度与单作的相同。
供试肥料为尿素、普通过磷酸钙和硫酸钾。玉米氮肥分3次施用, 分别为40%、25%和35%; 马铃薯氮肥分2次施入, 分别为60%和40%。磷肥(P2O5) 75 kg hm-2, 钾肥(K2O) 125 kg hm-2, 均以基肥形式施入, 各小区施肥量相同。马铃薯、玉米生育过程中中耕、培土、除草、病虫害防治等田间管理各处理保持一致。
盆栽试验设玉米单作、马铃薯单作及玉米马铃薯间作3个种植处理, 每个处理3次重复, 随机排列。间作处理中每盆玉米马铃薯各2株, 单作处理中玉米马铃薯每盆分别4株。采用400 mm× 280 mm的塑料盆, 每盆装土16 kg, 施肥量按150 mg kg-1土的纯养分来换算, N∶ P2O5∶ K2O = 2∶ 1∶ 1。氮肥分2次施用, 一半作基肥, 一半作追肥, 磷肥和钾肥作为基肥一次施入。供试肥料为尿素、普通过磷酸钙、硫酸钾。整个生育期按常规管理, 不使用杀菌剂和杀虫剂, 并及时人工除草, 定期调换盆的位置。
1.1.2 土壤样品的采集 于玉米(马铃薯)成熟期采样, 先将植株根系从土壤中整体挖出, 采用抖土法抖掉与根系松散结合的土, 然后将与根系紧密结合的土壤刷下来作为根际土样品。从田间试验单作处理每小区随机取作物4株, 间作处理每小区随机选2个间作带, 每个间作带内2种作物分别取2株, 然后将4株作物的根际土壤混合为1个样品; 盆栽试验单作处理每盆取4株, 间作处理2种作物分别取2株, 然后将根际土壤混合为1个样品, 最后将土样于4℃冰箱保存, 用于Biolog微平板分析。
采用Biolog ECO微平板方法[12] 测定土壤微生物代谢功能多样性。
采用平均颜色变化率(average well color development, AWCD)描述微生物代谢活性。采用Shannon多样性指数(H)、Simpson优势度指数(D)、均匀度指数(E)和群落丰富度指数(S)表征土壤微生物群落多样性。
AWCD = ∑ (Ci-Ri)/n
H = -∑ Pi (ln Pi)
D = 1-∑ Pi2
E = H/ln S
S = 被利用碳源的总数
式中, Ci为每一个微孔的光密度值, Ri为对照孔的吸光值, n为ECO板上碳源的种类, n=31; Pi为第i孔的相对吸光值与整个平板相对吸光值总和的比率, 即Pi = (Ci-Ri)/∑ (Ci-Ri); S为微生物群落利用碳源种类的数目, 即颜色变化孔数。
利用Microsoft Excel 2010进行数据初处理、制图和主成分分析, 采用SPSS19.0软件进行统计方差分析及聚类分析。
平均颜色变化率(AWCD)反映了土壤微生物的代谢活性, 是土壤微生物群落利用单一碳源的重要
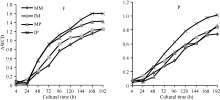
指标[13]。由图1可知, AWCD值随培养时间的延长而提高, 各处理的AWCD值在24 h之前很小, 说明在24 h之内碳源基本未被利用; 培养24 h后AWCD值快速升高, 反映出此后碳源被大幅度利用, 随后AWCD值缓慢增加直至趋于稳定。在各培养阶段, 间作处理的AWCD值均高于单作处理, 以间作马铃薯的AWCD值最高, 单作玉米最低。培养120 h时, 田间和盆栽试验中, 间作分别使玉米根际微生物的AWCD值比单作玉米增加17.36%和7.38%, 分别使马铃薯根际微生物的AWCD值比单作马铃薯增加3.76%和32.21%, 但单、间作处理间差异不显著(P< 0.05), 表明间作可以提高土壤微生物群落的代谢活性。
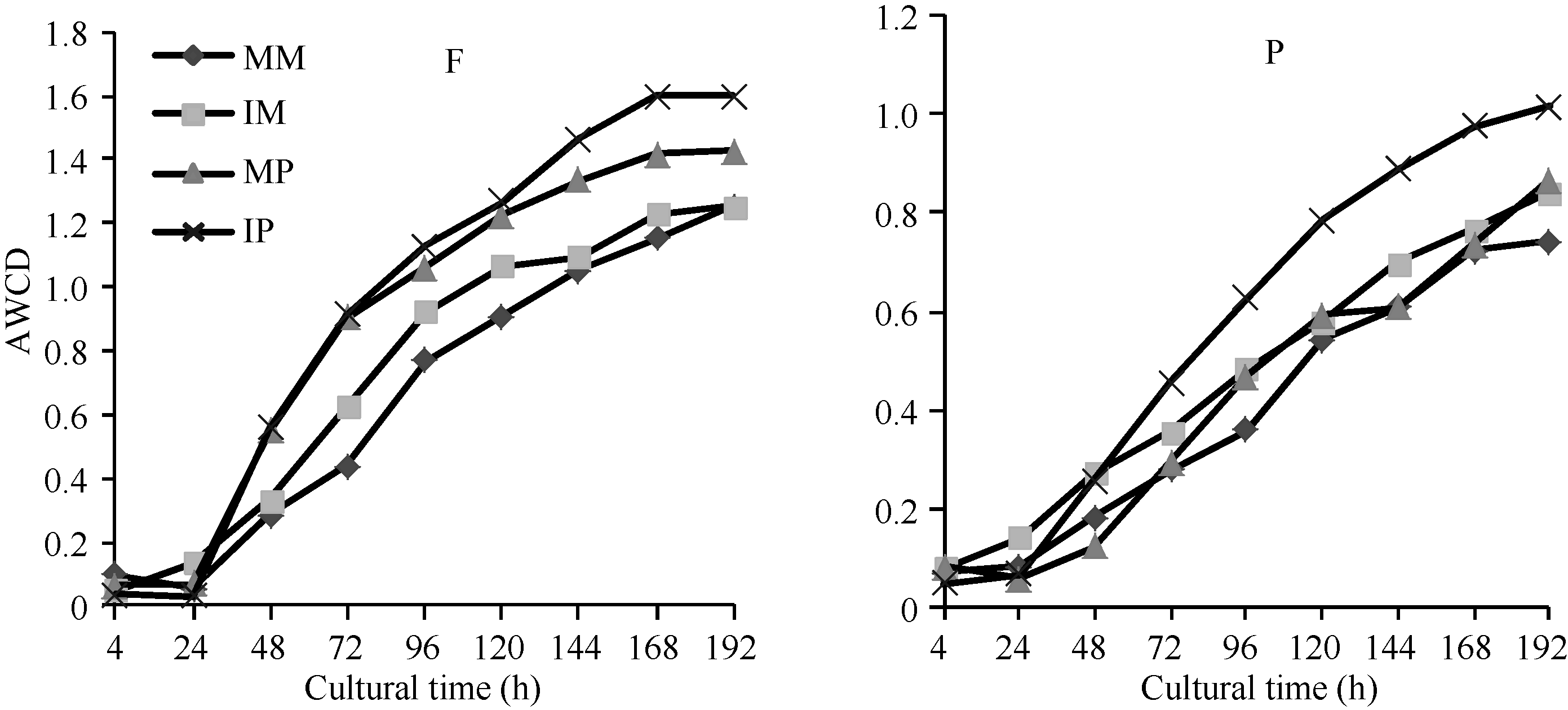 | 图1 根际微生物平均颜色变化率随培养时间的变化MM: 单作玉米; IM: 间作玉米; MP: 单作马铃薯; IP: 间作马铃薯。F: 田间试验; P: 盆栽试验。Fig. 1 Average well color development (AWCD) changes with incubation time of different treatmentsMM: monocropping maize; IM: intercropping maize; MP: monocropping potato; IP: intercropping potato. F: field experiment; P: pot experiment. |
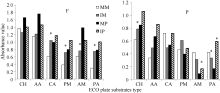
土壤微生物活性反映了群落总体的变化, 但未能反映微生物群落代谢的详细信息, 研究土壤微生物对不同碳源利用能力的差异, 有助于更全面地了解微生物群落代谢功能特性[14]。在田间和盆栽试验中(图2), 单作和间作条件下, 玉米、马铃薯根际微生物对6类碳源的利用情况有一定差异, 碳源利用强度总体以糖类、氨基酸类和羧酸类最高, 分别占总碳源利用强度的19.70%~30.91%、20.54%~ 26.04%、13.46%~18.18%和20.82%~31.39%、11.11%~24.87%、16.34%~24.20%。
间作对Biolog ECO板中6类碳源的利用均有影响(如图3), 但玉米和马铃薯根际微生物对6类碳源的利用情况存在较大的差异。田间试验中, 与单作相比, 间作提高了玉米根际微生物对氨基酸类、羧酸类、聚合物类、胺类和酚酸类碳源的利用, 分别是单作的1.1、1.7、1.9、1.2和2.6倍; 提高了马铃薯根际微生物对羧酸类、聚合物类和酚酸类碳源的利用, 分别增加17.79%、30.71%和27.26%。而间作降低了玉米根际土壤微生物对糖类的利用及马铃薯对糖类、氨基酸和胺类的利用, 但差异均不显著(P< 0.05)。
盆栽试验中, 间作提高了玉米根际微生物对糖类、氨基酸类、聚合物类碳源的利用, 其中糖类比单作显著增加了27.21%, 而降低了羧酸、胺类和酚酸类利用强度, 其中酚酸类比单作显著减少了20.67%; 提高了马铃薯根际微生物对6类碳源的利用, 其中胺类和酚酸类的代谢强度分别是单作的1.9倍和3.6倍, 且差异显著(P< 0.05)。
通过检测微生物对单一碳源利用程度, 可以反映微生物群体水平的群落结构, 用于测定微生物群落的功能多样性。如表1所示, 田间试验中, 间作玉米根际微生物高于单作玉米的代谢碳源包括6种碳水化合物、2种氨基酸、4种羧酸、2种聚合物、2种酚类和1种胺类; 间作马铃薯根际微生物高于单作马铃薯的代谢碳源包括3种碳水化合物、2种氨基酸、5种羧酸、3种聚合物、1种酚类和1种胺类。
盆栽试验中, 间作玉米根际微生物高于单作玉米的代谢碳源包括8种碳水化合物、3种氨基酸、3种羧酸、3种聚合物、1种酚类和1种胺类; 间作马铃薯根际微生物高于单作马铃薯的代谢碳源包括8种碳水化合物、4种氨基酸、4种羧酸、2种聚合物、2种酚类和1种胺类。
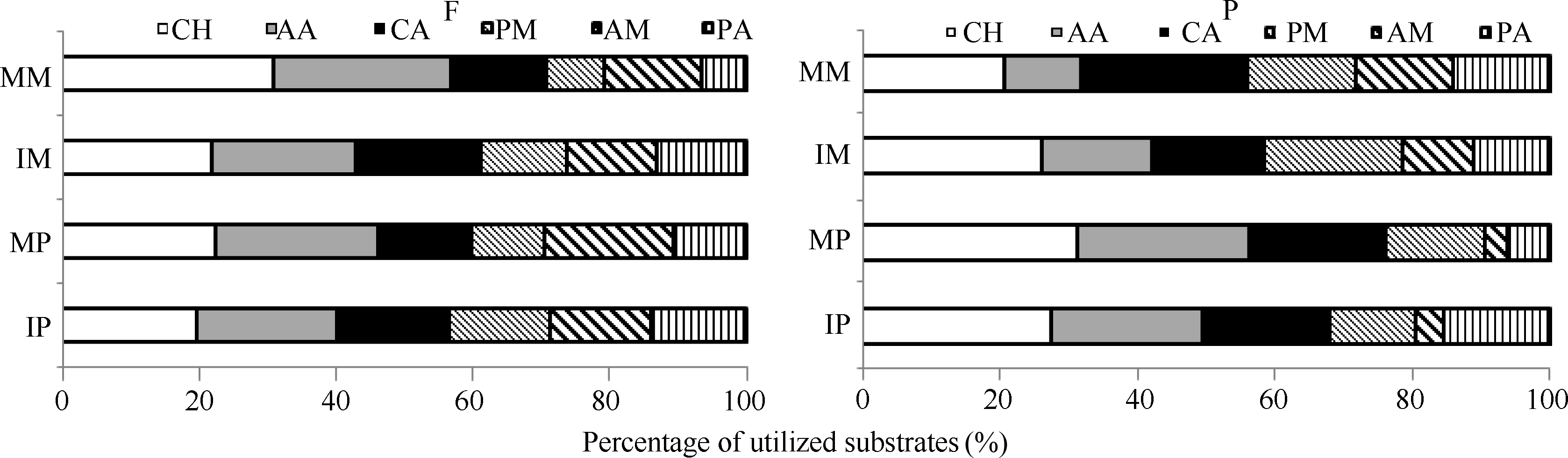 | 图2 根际微生物对碳源利用百分比CH: 糖类; AA: 氨基酸类; CA: 羧酸类; PM: 聚合物类; AM: 胺类; PA: 酚酸类。F: 田间试验; P: 盆栽试验。* 表示单作和间作处理间差异显著(P< 0.05)。缩写同图1。Fig. 2 Percentage of utilized substrates of rhizosphere microbeCH: carbohydrates; AA: amino acids; CA: carboxylic acids; PM: polymers; AM: amines/amides; PA: phenolic acids. F: field experiment; P: pot experiment. * means significant difference between monocropping and intercropping pattern at the 0.05 probobility level. Abbreviations are the same as those given in Figure 1. |
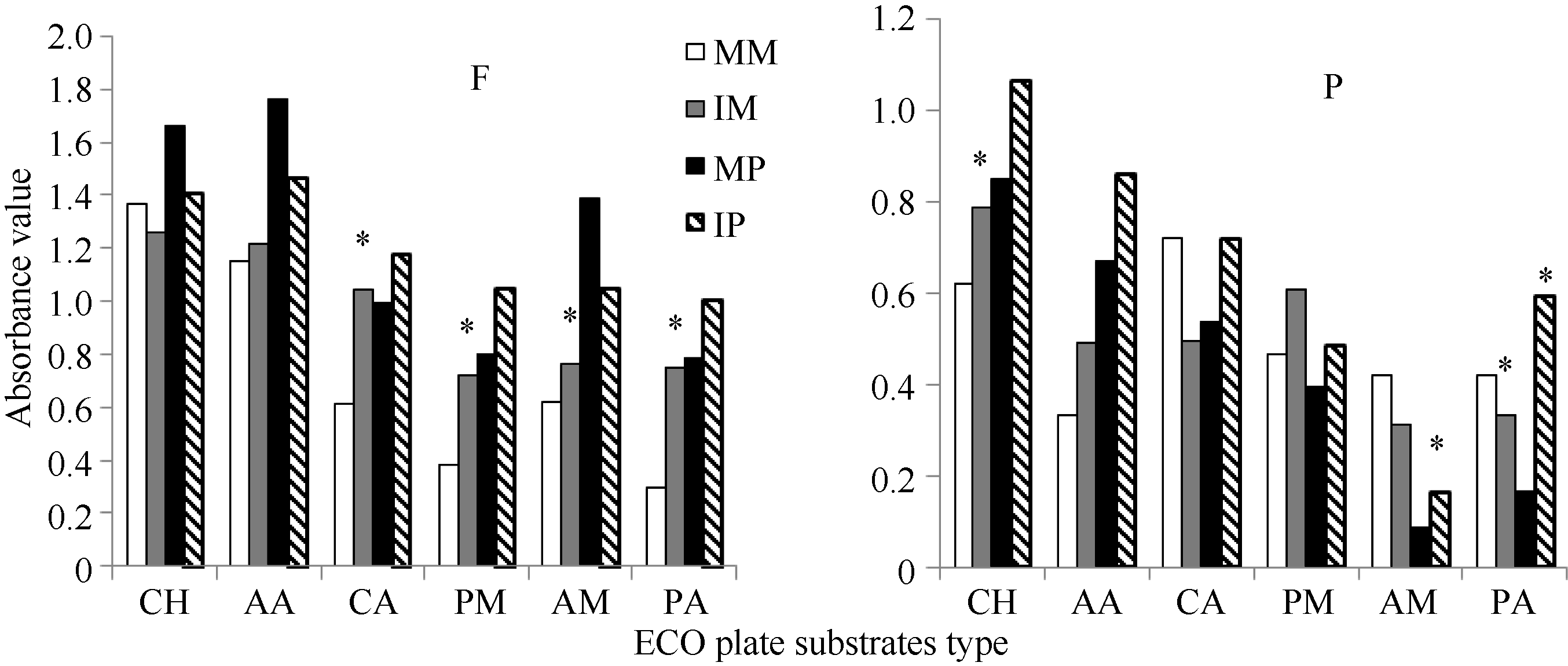 | 图3 根际微生物对6类碳源的利用强度F: 田间试验; P: 盆栽试验。* 表示单作和间作处理间差异显著(P< 0.05)。其他缩写同图1和图2。Fig. 3 Utilization intensity of rhizosphere microbe to six types of carbon sourceF: field experiment; P: pot experiment. * means significant difference between monocropping and intercropping pattern at the 0.05 probobility level. Other abbreviations are the same as those given in Figure 1 and Figure 2. |
土壤微生物群落利用碳源类型的多与少可以用各种多样性指数表示[15]。由表2可知, 在田间和盆栽试验中, 在培养120 h后, 与单作相比, 间作使玉米根际微生物群落Shannon指数(H)、Simpson指数(D)、均匀度指数(E)及丰富度指数(S)分别增加了5.26%、1.01%、4.43%、0和9.46%、0.21%、4.09%、19.23%, 使马铃薯分别增加4.78%、0.67%、4.35%、3.57%和1.41%、0.64%、-0.98%、11.11%, 但差异均不显著(P< 0.05)。
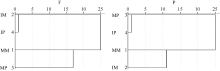
2.5.1 聚类分析 聚类分析可以将抽象对象的集合分组成为由类似的对象组成的多个类别, 可以更直观地显示各研究对象之间的远近关系, 通过对120 h不同处理的平均碳源利用率(AWCD)进行聚类分析(图4), 田间试验中, 间作玉米、间作马铃薯根际微生物具有与单作玉米、单作马铃薯明显不同的碳源利用模式。盆栽试验中, 单作马铃薯、间作马铃薯根际微生物群落利用碳源代谢特性相似, 单作玉米和间作玉米间具有明显不同的碳源利用模式。
| 表1 间作高于单作的碳源 Table 1 Increased Carbon source type utilized by maize and potato in different treatments |
| 表2 间作对根际微生物群落多样性指数的影响 Table 2 Effect of intercropping on rhizosphere microbial community diversity index |
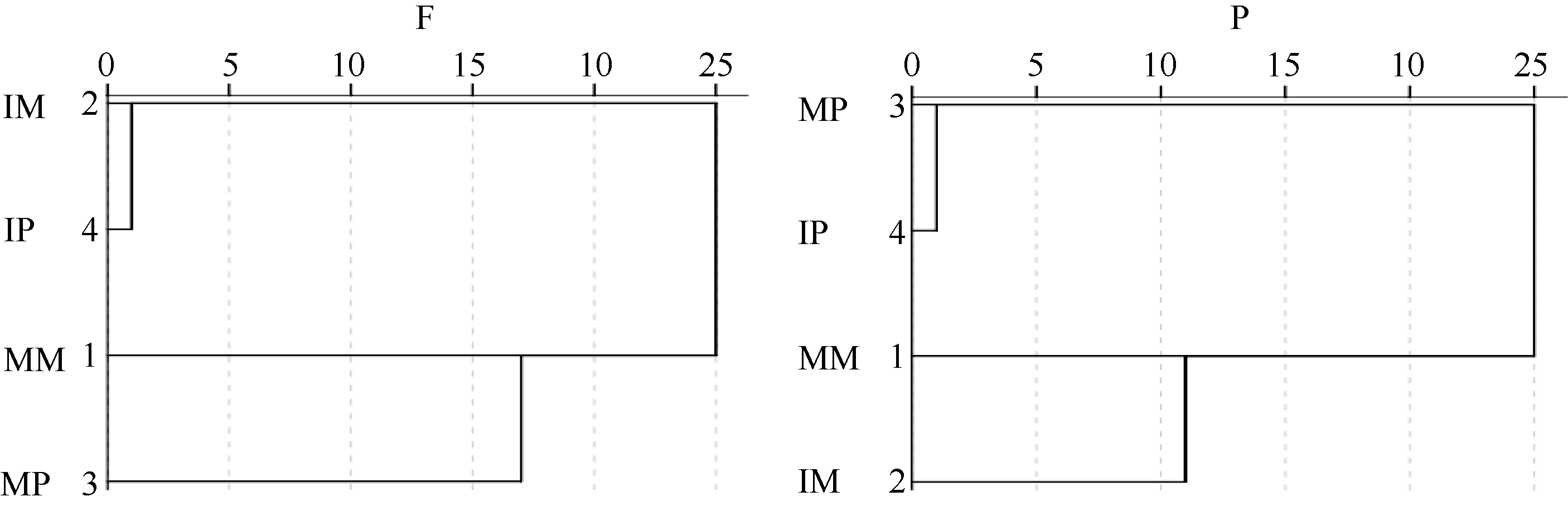 | 图4 根际微生物碳源利用特征的聚类分析F: 田间试验; P: 盆栽试验。其他缩写同图1。Fig. 4 Cluster analysis of carbon utilization profiles in maize and potato rhizosphere microbial communityF: field experiment; P: pot experiment. Other abbreviations are the same as those given in Figure 1. |
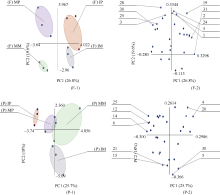
2.5.2 主成分分析 主成分分析是处理数学降维的一种方法, 将多个变量通过线性变换以选出较少个数重要变量。主成分个数的提取原则是相对应特征值大1的前m个主成分。根据此原则, 田间和盆栽试验中, 各共提取出了8个和10个主成分, 累计贡献率分别达到93.3%和100.0%, 第1主成分(PC1)贡献率分别是26.8%和25.7%, 第2主成分(PC2)贡献率分别是19.6%和18.0%。因第3~8主成分贡献率较小, 所以本文取前2个主成分的得分作图来表征微生物群落碳源代谢特征。
田间试验中, PCA分析表明(图5-F-1), 在PC1上, 玉米单间作、马铃薯单间作处理均有很好的分离, 单作玉米和单作马铃薯分布在负方向上, 间作玉米和间作马铃薯分布在正方向上; 在PC2上, 玉米单间作和马铃薯单间作处理的差异较小。同时, 如图5-F-2所示, 可以筛选出根际微生物利用能力较强的10种碳源, 分别是丙酮酸甲酯、聚山梨醇酯40、聚山梨醇酯80、肝糖、4-羟基苯甲酸、L-精氨酸、L-天门冬酰胺、L-苏氨酸、苯乙胺和腐胺。
盆栽试验中, PCA分析表明(如图5-P-1), 在PC1上, 玉米单间作处理的差异较小, 马铃薯单间作处理有较好的分离; PC2上, 玉米单间作处理有较好的分离, 马铃薯单间作处理的差异较小。同时, 如图5-P-2所示, 可以筛选出根际微生物利用能力较强的10种碳源, 分别是环糊精、肝糖、D-纤维二糖、N-乙酰-D-葡萄糖胺、1-磷酸葡萄糖、L-α -甘油磷酸、γ -羟丁酸、衣康酸、L-天门冬酰胺和苯乙胺。
初始载荷因子反映主成分与碳源利用的相关系数, 载荷因子越高, 表示该碳源对主成分的影响越大。Choi等[16]认为, 主成分1和主成分2上高于0.18或小于-0.18的载荷系数可认为具有较高的载荷量。
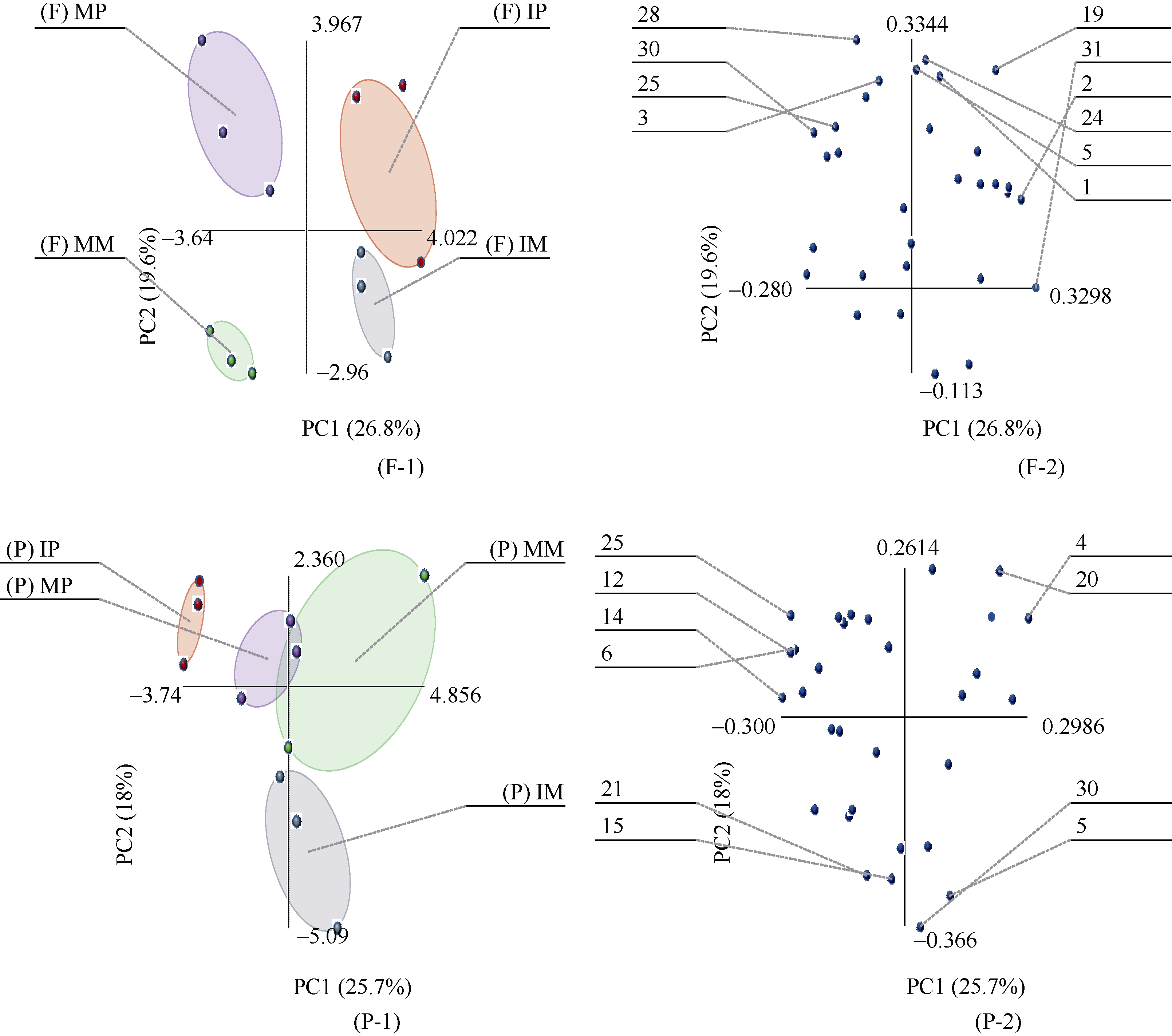 | 图5 单、间作处理根际微生物碳源利用特征的主成分分析F: 田间试验; P: 盆栽试验。其他缩写同图1。Fig. 5 Principal components analysis of carbon utilization profiles in maize and potato rhizosphere microbial community of monocropping and intercropping treatmentsF: field experiment; P: pot experiment. Other abbreviations are the same as those given in Figure 1. |
| 表3 ECO微孔板中在PC1和PC2上载荷较高的主要碳源 Table 3 Carbon substrates most heavily loaded on first two principal components (PC) in analysis of ECO micro-plate data |
土壤微生物群落AWCD反映了土壤微生物利用碳源的能力和代谢活性的大小, 其值越高, 土壤微生物群落代谢活性越高[17, 18]。研究表明, 合理的间套作可以提高土地利用率, 能够改善土壤根际微环境[19], 增加土壤微生物数量及改善微生物多样性[20]。能凤娇等[21]研究结果显示, 间作显著提高土壤微生物碳源利用能力和微生物群落功能多样性指数。本研究结果发现, 在整个培养过程中间作提高了玉米马铃薯根际微生物群落AWCD值, 其中间作马铃薯的AWCD值最高, 单作玉米的最低, 单、间作间差异不显著。说明间作在一定程度可以增强微生物的代谢活性, 提高土壤微生物对单一碳源的利用能力, 其原因可能是间作种植模式下存在2种作物的根系分泌物, 这些分泌物可以为根际微生物提供生长繁殖所需的能源, 影响根际微生物的数量和群落结构的变化[22]。
土壤微生物多样性指数、优势度指数、均匀度指数和丰富度指数是表征群落多样性的常用指数, 可以揭示土壤微生物种类和功能的差异[23]。董艳等[24]研究发现, 间作显著提高蚕豆、小麦根际微生物的Shannon多样性指数与丰富度指数。章家恩等[25]研究发现, 玉米花生间作改善了根区微生物群落功能多样性和代谢活性。本研究中, 间作玉米和间作马铃薯根际微生物群落的Shannon指数(H)、Simpson指数(D)、均匀度指数(E)及丰富度指数(S)分别高于单作玉米和单作马铃薯, 这表明间作使土壤微生物群落多样化, 其原因可能是间作系统中不同作物会产生不同的特异根系分泌物, 并形成与之相适应的根际微生物群落, 从而提高土壤微生物的整体代谢活性, 促进土壤微生物群落结构多样化的形成[25, 26]。
李鑫等[22]研究发现, 不同种植模式下土壤微生物对6类碳源的利用强度差异较大。本研究田间试验发现, 间作提高了玉米根际微生物对氨基酸、羧酸、聚合物、胺类和酚酸类碳源的利用, 降低了对糖类的利用; 提高马铃薯根际微生物对羧酸、聚合物和酚酸类碳源的利用, 降低了对糖类、氨基酸和胺类的代谢强度。盆栽试验中, 间作提高了玉米根际微生物对糖类、氨基酸类、聚合物类碳源的利用, 降低了对羧酸、胺类和酚酸类的利用; 提高了马铃薯根际微生物对6类碳源的代谢强度。从对单一碳源的利用差异来看, 田间和盆栽试验中, 间作玉米根际微生物代谢碳源分别主要有17种和19种高于单作, 间作马铃薯根际微生物代谢碳源分别主要有15种和21种高于单作。表明玉米与马铃薯间作条件下产生的根系分泌物可能使土壤微生物对某些碳源的利用能力有一定的抑制作用, 这些碳源基质利用能力的消弱与有利于利用此类碳源基质的土壤微生物种群数量的减少有一定关系, 其原因可能是间作中作物根系的互作可增加根系土壤中含有更多的维生素、碳水化合物、氨基酸和有机酸等含量, 使植物根系分泌物种类改变及化合物积累, 增强了土壤微生物功能多样性, 但可能存在一定的正反馈调节作用[27]。
应用31种碳源的光密度值聚类分析的结果显示, 田间试验中单、间作种植下玉米马铃薯根际微生物群落具有明显不同的碳源代谢模式; 盆栽试验中单、间作玉米根际微生物群落也具有明显不同的碳源利用模式, 这可能是因为间作种植在很大程度上改变了土壤微环境。
主成分分析解释了不同处理土壤微生物碳源利用存在的差异。水稻与西瓜间作改变了根系分泌物中糖类、氨基酸和酚酸的组成, 增加了根际土壤的微生物多样性[28]。小麦与蚕豆间作系统中, 糖类、羧酸类和氨基酸类是微生物利用的主要碳源, 可作为区分不同种植模式下微生物碳源利用类型的依据[24]。本试验条件下, 在田间试验中, 玉米和马铃薯的单间作处理在PC1上出现了明显的分异。在盆栽试验中, 马铃薯单间作处理在PC1上出现了较好的分离; 玉米单间作处理在PC2上出现较好的分离, 表明间作种植明显改变了根际微生物的功能多样性。间作改变根际微生物群落功能多样性是由微生物对PC1和PC2上载荷较高的碳源利用差异引起的, 在PC1和PC2上载荷较高的主要碳源是糖类、氨基酸类和羧酸类, 这3类碳源可作为区分间作种植模式下微生物碳源利用类型的敏感碳源。表明玉米与马铃薯间作可能改变了根系分泌物的组成, 尤其是氨基酸、糖类和有机酸的组成, 本研究结果与上述研究结果相似。
玉米、马铃薯间作提高了根际土壤微生物群落碳源利用率(AWCD)、微生物群落的Shannon指数(H)、Simpson指数(D)、均匀度指数(E)及丰富度指数(S), 但根际微生物对6类碳源的利用存在一定的差异。玉米与马铃薯间作改变了根际微生物的群落组成, 糖类、羧酸类和氨基酸类碳源是区分单间作处理差异的主要碳源。间作种植方式是维持土壤微生物群落功能多样性和土壤生态系统稳定性与可持续发展的重要措施。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|