{kind=link}
{kind=link}
外源水杨酸对Cu胁迫下水培烟草生长及营养元素吸收利用的影响
[徐根娣1, 2  , 葛淑芳
, 葛淑芳1, 2 , 章艺3 , 吴玉环4 , 梅笑漫5 , 刘鹏1, 2, * ]
, 葛淑芳]
|
|


第一作者联系方式: E-mail: sky113@zjnu.cn
以耐Cu性较强的烟草品种W38 ( Nicotiana tabacumcv. W38)和耐Cu性较弱的本氏烟( Nicotiana benthamiana)为试验材料, 设置水培处理组即CK (Cu2+ 0 mg L-1, SA 0 μmol L-1, 即对照组)、T1(Cu2+ 4 mg L-1, SA 0 μmol L-1)、T2 (Cu2+ 4 mg L-1, SA 100 μmol L-1)、T3 (Cu2+ 4 mg L-1, SA 300 μmol L-1)、T4 (Cu2+ 4 mg L-1, SA 500 μmol L-1), 探讨SA对烟草生长特性及根、茎、叶不同器官中元素吸收规律的影响。结果表明, 经过4 mg L-1Cu胁迫15 d后, 2个品种的烟草生长均受到一定程度的抑制, 不同器官中Cu含量均显著升高, K、Ca、Mg、Fe、Zn、B、Mn的吸收受到抑制, 而在营养液中添加适宜浓度的SA能够有效缓解Cu胁迫对2个品种烟草根长、株高、鲜重的抑制作用, 并降低烟草体内Cu含量。与T1处理组相比, 本氏烟的根、茎、叶中Cu含量最大分别下降25.05%、39.78%和22.91%, W38的根、茎、叶中的Cu含量最大分别下降23.27%、37.30%和28.88%, 并促进了营养元素K、Ca、Mg、Fe、Mn的吸收, 但是对Zn、B元素的吸收影响并不明显。由此可知, 高浓度的Cu胁迫会抑制烟草的生长及营养元素的吸收和运输, 施加适宜浓度的外源水杨酸有利于营养元素的吸收和促进烟草的生长发育, 在本试验浓度范围内施加300 μmol L-1的水杨酸可显著缓解Cu胁迫对烟草的抑制作用。
A hydroponic experiment was conducted to study the effects from treatmeats including CK ( Cu2+ 0 mg L-1, SA 0 μmol L-1 as control), T1 (Cu2+ 4 mg L-1, SA 0 μmol L-1), T2 (Cu2+ 4 mg L-1, SA 100 μmol L-1), T3 (Cu2+ 4 mg L-1, SA 300 μmol L-1), T4 (Cu2+ 4 mg L-1, SA 500 μmol L-1) on growth characteristics, elements absorption and accumulation in two tobacco cultivars, N. tabacum cv .W38 (Cu resistant genotype) and N. benthamiana (Cu sensitive genotype) under copper stress. The result showed that the growth of two tobacco cultivars under 4 mg L-1Cu stress for 15 days was inhibited to some extent, meanwhile, the content of Cu in their organs was increased while the absorptions of K, Ca, Mg, Fe, Zn, B, and Mn were inhibited. The addition of exogenous SA could alleviate the inhibition of plant height, root length and fresh weight, and reduce Cu content of tobacco, but increase the absorption of nutritional elements K, Ca, Mg, Fe, and Mn. Compared with treatment T1, contents of Cu in root, stem and leaf were increased by 25.05%, 39.78%, 22.91% in N. benthamiana, and by 23.27%, 37.30%, 28.88% in N. tabacum cv. W38, respectively. In conclusion, appropriate concentration of SA promotes the growth and elements absorption in tobacco under copper stress. In this experiment, 300 μmol L-1 SA could significantly alleviate copper inhibition to tobacco.
近年来随着现代化工农产业的飞速发展, 土壤的重金属污染日趋严重。重金属在环境中具有难降解性和相对稳定性, 且会在植物的根、茎、叶中大量积累, 这不仅严重阻碍了植物的正常生长和发育, 而且会通过食物链进而危害动物和人类的健康[1]。水杨酸(salicylic acid, SA), 即邻羟基苯甲酸, 植物体内广泛存在, 目前已证明能够调节植物体内的许多抗逆生理过程, 如抗盐性、抗寒性、抗热性、抗旱性以及抗病性[2, 3], 同时作为一种植物激素, SA也是用于增加作物收成的一种重要措施, 具有广阔的应用前景[4, 5, 6, 7]。
重金属胁迫对植物造成毒害的机制主要是重金属离子取代植物必需营养元素与酶活性中心或蛋白中心的巯基结合, 或是干扰Ca2+等元素的运输并启动氧化还原反应产生自由基, 造成植物的氧化损伤, 进而造成植株代谢紊乱[8]。铅、镉等重金属胁迫能使黄瓜、大麦等幼苗的内源SA水平增加[9, 10], Freeman等[11]等发现拟南芥中自由态SA含量的升高, 能够激发丝氨酸乙酰转移酶活性, 进而使植株体内谷胱甘肽含量升高, 增强抗逆性。外源SA的施加还会影响植物对重金属和营养元素的吸收、运输。Drazic和Mihailovic[12]研究SA对Cd胁迫下大豆幼苗对营养元素吸收规律的影响, 发现SA能减低K含量却促进了Fe的吸收, 同时也促进了Mg和Ca的吸收与运输, 使植物体内各元素间达到动态平衡, 提高机体抗胁迫的能力。总之, 目前有关SA诱导植物重金属抗性的机理研究仍然处于起步阶段, 已有的研究一般证实了SA能在一定程度上缓解重金属对植物的毒害, 但不同植物或相同植物的不同组织对SA抗重金属的机制可能有所不同, 甚至互相矛盾, 其中有许多问题亟待解决, 且有关SA对烟草Cu胁迫下营养元素吸收运输的影响国内外至今尚未见报道, 因此本文以烟草为试验材料, 旨在探讨外源SA对烟草营养元素吸收运输的调控机制及其可能的调控技术途径, 以期为进一步提高烟叶品质提供一定的理论依据, 且为抗性生理增添素材, 为重金属污染的防治提供科学参考。
以烟草根相对伸长率为指标, 从云烟85、云烟87、MS云烟85、W38、MSK326、NC89、双-70、89112、本氏烟等9个烟草品种中筛选出耐Cu性较强的W38和耐Cu性较弱的本氏烟, 设置施铜和不施铜2个处理。烟草种子购买于云南玉溪烟草种子公司。SA购买于金华医药有限公司。
先将烟草种子经10%的H2O2表面消毒15 min, 然后在25℃下黑暗浸种24 h, 之后将其播种到铺有滤纸的培养皿中, 等种子发芽后转移到沙子中继续培养, 待幼苗的第4片真叶展开后, 选取长势较为一致的幼苗移入Hoagland 1/2完全营养液, 预培养3 d, 再转入完全营养液预培养6 d, 然后按以下处理继续培养: CK (Cu2+ 0 mg L-1, SA 0 μ mol L-1, 即对照组)、T1 (Cu2+ 4 mg L-1, SA 0 μ mol L-1)、T2 (Cu2+ 4 mg L-1, SA 100 μ mol L-1)、T3 (Cu2+ 4 mg L-1, SA 300 μ mol L-1)、T4 (Cu2+ 4 mg L-1, SA 500 μ mol L-1)。其中, Cu2+以CuSO4· 5H2O的形式提供, SA采用直接添加到营养液中的方式, 每3 d更换1次营养液, 处理15 d后取样测定相应生理指标, 每个处理3个重复。试验材料和试验所用浓度由前期大量预试验所得。
根系长度、株高用直尺直接测量; 植株鲜重用电子天平称量。收集烟草的根、茎、叶, 用去离子水反复冲洗, 茎、叶部分先在105℃下杀青15 min, 后与根一起于70℃的烘箱中烘干至恒重, 再研磨成粉末。精确称取每个样品0.3 g加入HNO3-HClO4消化, 待溶液变得澄清透明时, 高温烘干, 此时有白色晶体析出, 再用去离子水将其定容至50 mL, 即得待测溶液。采用7500a型电感耦合等离子体质谱仪(ICP-MS) (美国Agilent公司), 测定Cu、K、Ca、Mg、Fe、Zn、B和Mn等8种元素。
所有测定均设3次重复, 计算平均值和标准误差, 采用SPSS 18.0软件中的Duncan’ s法进行显著性差异分析, 用Microsoft Excel 2007软件制图。
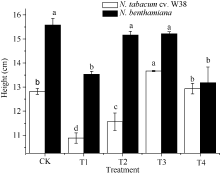
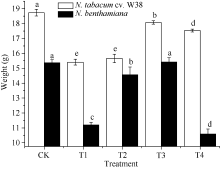
外源SA对Cu胁迫下烟草生长特性的影响如图1所示, T1处理下, W38和本氏烟的根长显著降低, 与对照组相比分别降低8.56%和12.23%, 由此可知, Cu胁迫对本氏烟的根长抑制率大于W38, 表现出其对Cu毒的敏感性。施加一定浓度的外源SA可以缓解根长的抑制作用, 当SA浓度为300 μ mol L-1时能有效缓解Cu毒, 与T1相比, W38和本氏烟的根长分别上升5.95%和13.50%。由图2和图3可知, 2个品种烟草的株高和鲜重在不同处理的变化趋势大致是一致的, T1处理条件下其株高和鲜重均显著下降, 在一定范围内随着施加外源SA浓度的升高而不断增加, 但是当外源SA浓度大于500 μ mol L-1时, 却对烟草的生长产生抑制作用, 其中本氏烟表现更为突出, 这可能是因为高浓度的外源SA对烟草产生了一种新的胁迫作用。
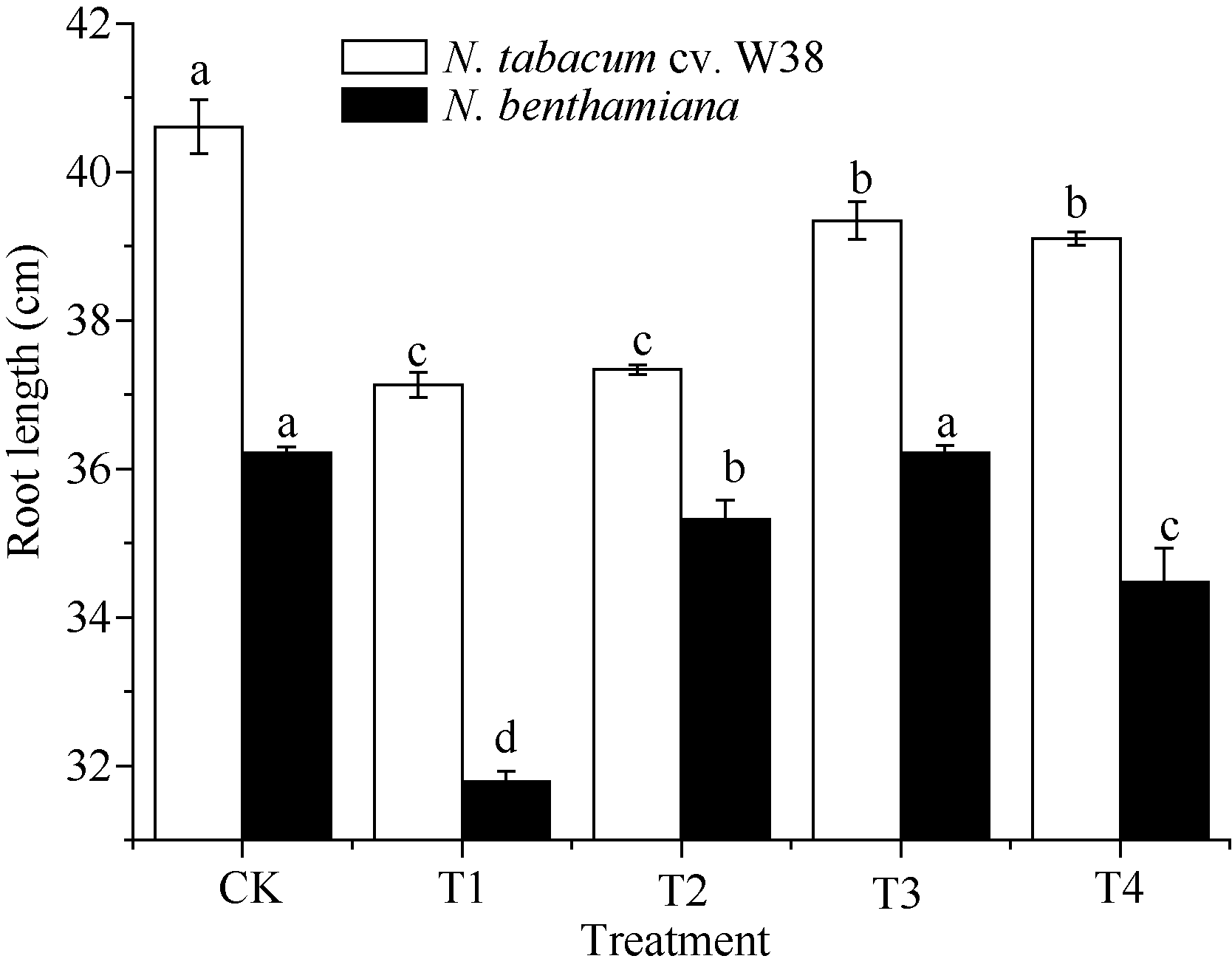
SA对铜胁迫下烟草根长的影响 Effects of SA on the root length (mean± SE) of tobacco under copper stress Bars superscripted by different letters are significantly different at the 5% probability level. CK: Cu2+ 0 mg L-1, SA 0 μ mol L-1, T1: Cu2+ 4 mg L-1, SA 0 μ mol L-1; T2: Cu2+ 4 mg L-1, SA 100 μ mol L-1, T3: Cu2+ 4 mg L-1, SA 300 μ mol L-1, T4: Cu2+ 4 mg L-1, SA 500 μ mol L-1.
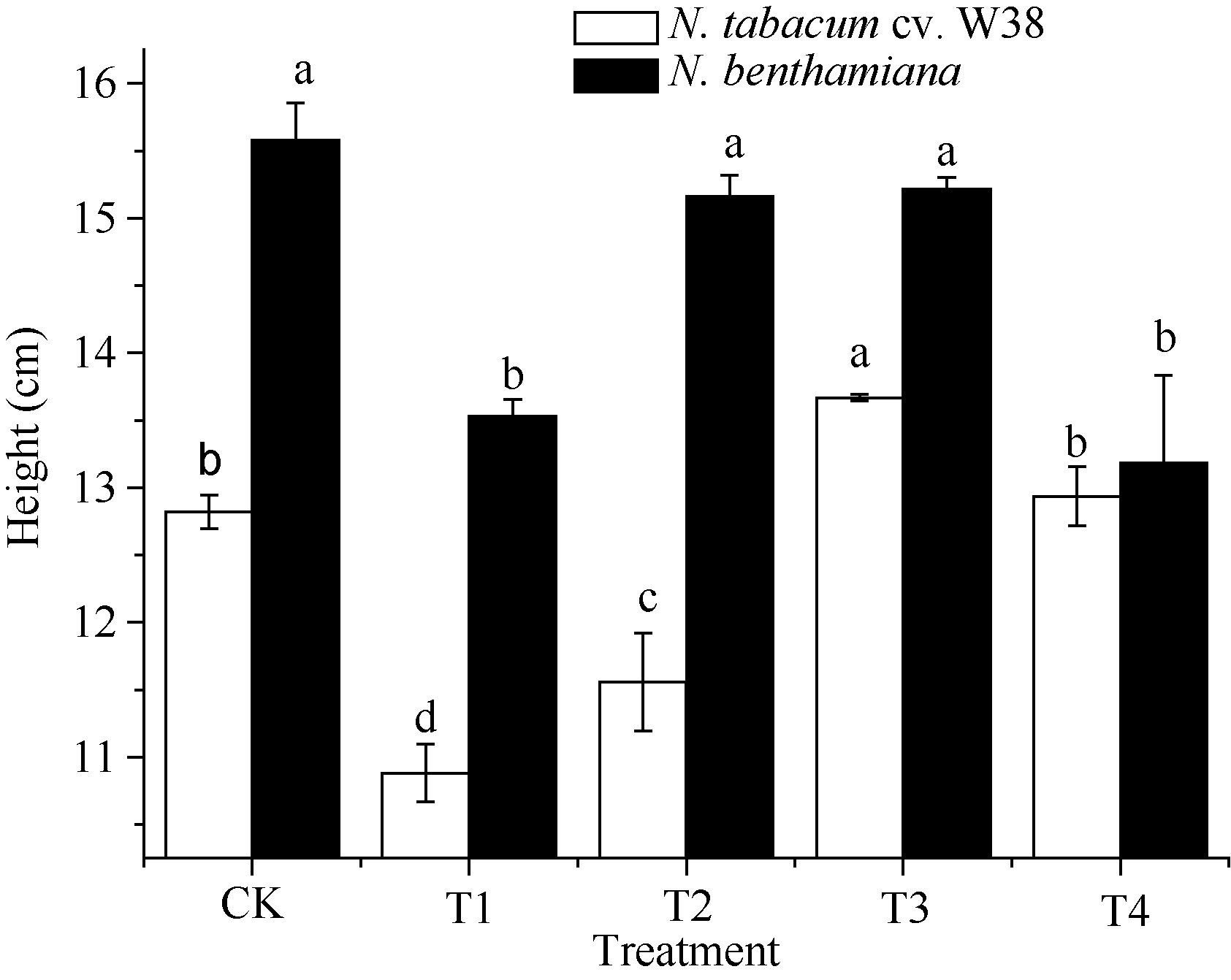 | 图2 SA对铜胁迫下烟草株高的影响Fig. 2 Effects of SA on plant height (mean± SE) of tobacco under copper stressBars superscripted by different letters are significantly different at the 5% probability level. Treatment codes are the same as those given in Fig. 1. |
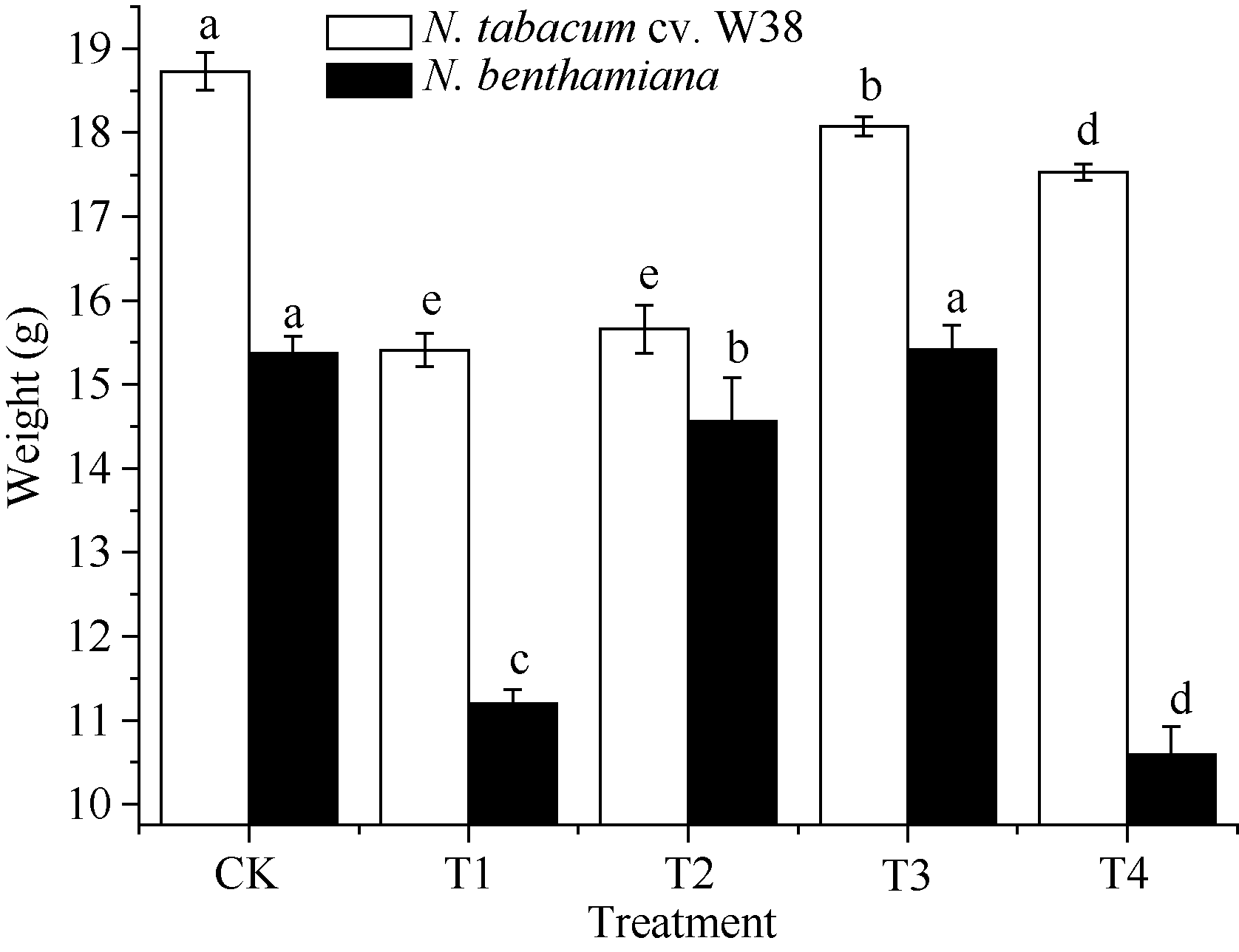 | 图3 SA对铜胁迫下烟草鲜重的影响Fig. 3 Effects of SA on fresh weight (mean± SE) of tobacco under copper stressBars superscripted by different letters are significantly different at the 5% probability level. Treatment codes are the same as those given in Fig. 1. |
如表1所示, 与对照组相比, 在4 mg L-1Cu胁迫下2个品种烟草地上部和地下部Cu含量均呈显著增加趋势。施加外源SA后, 2个品种烟草Cu含量均有明显的降低趋势。外源SA浓度为100 μ mol L-1时对本氏烟的缓解效果较好, 与T1胁迫下相比, 其根、茎、叶中Cu含量分别下降25.05%、39.78%和22.91%。对于W38品种来说, 当外源SA浓度为300 μ mol L-1时对其缓解效果较好, 其根、茎、叶中的Cu含量分别下降23.27%、37.30%和28.88%。
| 表1 SA对铜胁迫下烟草Cu含量的影响 Table 1 Effects of SA on the content of Cu (mean± SE) in tobacco under copper stress (μ g g-1DW) |
由表2可以看出, 与对照相比, T1处理条件下2个品种烟草根、茎、叶中K含量均显著下降, W38分别下降46.01%、25.13%和35.87%, 本氏烟分别下降49.62%、32.56%和35.30%, 这可能是Cu毒抑制了根系对K的吸收, 进而降低了烟草地上部K含量。添加外源SA后, 2个品种烟草根、茎、叶中K含量均有所增加, 但是当SA浓度为500 μ mol L-1时, 本氏烟各部分的K含量却呈现下降趋势, 说明高浓度的SA对烟草的生长发育有一定的抑制作用。
从表3可以看出, T1处理与对照组相比, 烟草根、茎、叶Ca含量迅速下降, 这说明Cu对Ca的吸收有抑制作用。在烟草植株各部位的Ca含量分布不同, 叶> 茎> 根。外源SA浓度为300 μ mol L-1时, W38各器官Ca含量均达最高, 与T1相比, 其根、茎、叶的Ca含量分别上升23.90%、17.22%和8.70%, 可见施加外源SA后对根部Ca的吸收促进作用最为明显。本氏烟的Ca含量随着外源SA浓度的增加而呈现递减趋势, 外源SA浓度100 μ mol L-1时, 缓解效果较好, 与T1处理相比, 本氏烟根、茎、叶Ca含量分别上升16.94%、11.21%和5.52%。因此, 在Cu胁迫下, 施加一定浓度的SA可以促进烟草尤其是根部Ca元素的吸收。
由表3可知, 耐Cu性不同的W38和本氏烟相同部位中Mg含量的差异不大, 且在不同处理下对Mg的吸收变化趋势也是相似的。在Cu胁迫下2个烟草品种中Mg含量均有所下降, 但是在添加外源SA后并没有显著提高烟草各部位的Mg含量, 由此可知, 外源SA对Cu胁迫下Mg元素的吸收和运输影响不大。此外, Mg元素在2个烟草品种根中的含量均大于在茎和叶中。
| 表2 SA对铜胁迫下烟草K含量的影响 Table 2 Effects of SA on the content of K (mean± SE) in tobacco under copper stress (μ g g-1 DW) |
| 表3 SA对铜胁迫下烟草Ca、Mg含量的影响 Table 3 Effects of SA on contents of Ca and Mg (mean± SE) in tobacco under copper stress (μ g g-1 DW) |
由表4可知, T1处理对烟草微量元素吸收有不同程度的抑制作用, 其中Fe和Mn元素受到的抑制作用最大, B, Zn受到的抑制作用最小。施加外源SA后Fe、Mn元素的含量有所回升但是仍然低于对照组, 这说明外源SA缓解Cu毒的能力是有限的。而施加外源SA后, Zn、B元素含量并没有显著的变化, 可见SA对Cu胁迫下Zn、B吸收和运输的影响不大。
| 表4 SA对铜胁迫下烟草Fe、Zn、B、Mn含量的影响 Table 4 Effects of SA on contents of Fe, Zn, B, and Mn (mean± SE) in tobacco under copper stress (μ g g-1 DW) |
Cu是植物正常生命活动所必需的微量元素之一, 对维持植物正常的新陈代谢及生长发育均具有极其重要的意义, 然而, 适合植物生长的铜浓度范围却很窄, 土壤中稍微过量的Cu便会干扰细胞的正常代谢及离子平衡继而会对植物造成严重的毒害作用[13]。本研究表明, 烟草植株生长在Cu浓度为4 mg L-1时, 与对照相比, 其生长受到抑制, 表现为根长、株高以及单株鲜重值显著下降。已有的研究表明, SA对植物抗性的诱导具有浓度效应, 表现为低促高抑[14]。本研究表明, 100~300 μ mol L-1的SA可以不同程度地缓解铜对烟草幼苗根长、株高及鲜重的抑制作用, 特别是SA质量浓度为300 μ mol L-1时, 缓解效果最明显。说明SA可以诱导植物产生抗铜性。但当SA质量浓度达到500 μ mol L-1时, 不仅不会缓解铜对烟草幼苗生长的抑制作用, 反而产生更严重的毒害作用, 这可能是高质量浓度的SA与Cu共同作用产生双重毒害的结果。
营养元素的吸收平衡被破坏是重金属植物毒理学效应的一个重要方面[15]。植物Cu毒害的一个重要机制就是严重阻碍植物对养分的吸收及运输, 最终导致植物营养的亏缺或失衡。Cu、K、Ca、Mg、Fe、Zn、B和Mn都是植物正常生长和代谢所必需的营养元素, 其中Cu是许多氧化酶的成分及某些酶的活化剂, 在植物的光合作用、呼吸作用等方面的氧化还原过程中有着非常重要的作用[16], K提高植物的抗逆性, 能增强碳水化合物的合成和转运, 能激活多种酶, 能调节渗透浓度, 另外, K能促进蛋白质和核酸的形成。
Ca是细胞代谢的总调和者, 被称为“ 第二信使” , 在多种细胞信号转导途径中起着重要的作用, 是参与细胞构建及代谢必需的营养元素, Mg是植物叶绿素的重要组成成分之一, 在光合作用和呼吸作用过程中可以活化各种磷酸变位酶和磷酸激酶, 且Mg也可以活化DNA和RNA的合成过程[17], Fe作为植物必需的微量营养元素, 是许多重要酶的辅基, 如细胞色素、过氧化物酶等, Zn是植物多种代谢途径中关键酶的组成因子, 同样也是叶绿素合成的必需元素, B的主要生理功能是参与糖的转运与代谢, 可以调节植物细胞壁的形成、N代谢等生理过程, Mn是多种酶的活化剂, 可以直接参与植物的光合作用过程, 同时对氧化还原反应中的电子传递有着非常重要的影响。此外, Cu胁迫下植物的根原生质膜上的Ca2+通道会受阻, 从而导致根尖细胞Ca2+净吸收量下降, 原生质膜上Ca2+缺乏, 这种平衡被破坏, 进而对细胞的结构和功能造成严重的影响, 同时导致Mg在根系阳离子交换量的饱和度降低, 其吸收量的下降, 继而会导致叶绿素合成的受阻。K、Fe在植物体内参与多种酶的合成, 同时在植物体的吸收、代谢、蛋白质合成等过程中不可或缺[18]。Cu胁迫阻碍了植物的正常生长发育, 同时也影响了营养元素的吸收和运输。本研究发现, 过量的Cu改变了烟草细胞阳离子的交换能力, 致使烟草根系和地上部的Cu含量增加, 抑制了植株的生长, 减少了烟草对K、Ca、Mg、Fe、Zn、B和Mn养分的吸收, 降低了烟草植株各部位K、Ca、Mg、Fe、Zn、B、Mn元素的含量。可能机制之一是Cu2+竞争性地抑制了其他离子的吸收, 如Cu2+通过与膜上特殊通道受体结合位点结合, 使根系原生质膜Ca2+通道受阻, 阻碍Ca2+在膜上的整合, 进而影响了Ca2+跨膜运输的GTP蛋白的活性, 降低了原生质体Ca2+通道的流量, Ca2+平衡遭到破坏, 进而影响细胞结构和正常的代谢功能, 导致Ca2+在根系阳离子交换量的饱和度下降。可能机制之二是Cu2+胁迫影响根系活力, Cu2+浓度超过一定值时会明显降低根系活力[19, 20], 对烟草发育产生毒害, 本试验中K、Ca、Mg、Fe、Zn、B和Mn的吸收在Cu2+胁迫时明显下降, 主要原因可能是根系活力减弱导致养分吸收受阻。由此可见, Cu胁迫造成的根系元素吸收受抑及养分分配平衡失衡都加剧了植株Cu毒害, 并且降低了植株的产量和品质。此外我们还发现, 外施SA有效的抑制了Cu胁迫下Cu含量的增加, 缓解了Cu这种重金属的过量吸收对烟草造成的不良影响, 同时, 外施SA可以有效缓解烟草对大量元素K、Ca、Mg和微量元素Fe、Mn吸收量的减少, 这可能是因为添加外源SA对溶液中Cu2+形成了络合物或螯合物, 从而降低了Cu的胁迫性。在植物体内蛋白质、核酸等重要物质的合成和代谢都离不开K、Mg的参与, 它们在植物体内含量的降低必将导致植物的代谢紊乱[21], 这与SA减轻Cu对细胞膜结构的损伤有关。此外, 外源SA对Cu胁迫下W38和本氏烟营养元素吸收均有很好的促进效果, 且对铜敏感型品种本氏烟的营养元素吸收的促进作用更为显著, 这与周媛[22]的结果较为一致。由此可知, 外源SA的添加能够有效缓解Cu胁迫下烟草对营养元素吸收的不足, 促进元素的吸收和由地下向地上部分的运输。
烟草对营养元素的吸收是有平衡要求的, 营养元素在烟草的不同器官部位分配不均衡将严重危害其正常的代谢及生长发育, 而Cu胁迫下施加一定浓度的外源SA可以在一定程度上恢复烟草对营养吸收的动态平衡和缓解烟草营养缺乏和离子毒害症状。在本实验浓度范围内施加300 μ mol L-1的SA可显著缓解Cu胁迫对烟草生长的抑制作用及促进烟草对营养元素的吸收利用, 进而提高烟叶的品质。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|