{kind=link}
[成良强, 唐梅 , 任小平, 黄莉, 陈伟刚, 李振动, 周小静, 陈玉宁, 廖伯寿, 姜慧芳
, 任小平, 黄莉, 陈伟刚, 李振动, 周小静, 陈玉宁, 廖伯寿, 姜慧芳* ]
, 任小平, 黄莉, 陈伟刚, 李振动, 周小静, 陈玉宁, 廖伯寿, 姜慧芳]
|
|


第一作者联系方式: E-mail: chengliangmei1984@126.com, Tel: 13971402933
栽培种花生是异源四倍体, 基因组大, 构建花生的分子遗传连锁图谱并对相关性状进行QTL定位研究的工作缓慢。本研究以遗传差异大的亲本组配杂交组合富川大花生×ICG6375构建F2作图群体, 采用公开发表的2653对SSR引物, 构建了一张含有234个SSR标记、分布于20个连锁群的栽培种花生遗传图谱。该图谱覆盖基因组的长度为1683.43 cM, 各个连锁群长度在36.11~131.48 cM之间, 每个连锁群的标记数在6~15个之间, 标记间的平均距离为7.19 cM。结合F3在湖北武汉和阳逻环境下的主茎高和总分枝数鉴定结果, 应用WinQTLCart 2.5软件采用复合区间作图法进行了QTL定位和遗传效应分析。共检测到17个与主茎高和总分枝数相关的QTL位点, 贡献率在0.10%~10.22%之间, 分布于8个连锁群上。综合分析武汉和阳逻环境的鉴定结果, 获得重复一致的与主茎高相关的6个QTL, 其中 qMHA061.1和 qMHA062.1位于连锁群LG06上TC1A2~AHGS0153标记区间, 贡献率为5.49%~8.95%; qMHA061.2和 qMHA062.2位于LG06上AHGS1375~PM377标记区间, 贡献率为2.93%~5.83%; qMHA092.2和 qMHA091.1位于连锁群LG09上GM2839~EM87标记区间, 贡献率为0.53%~9.43%。
, TANG Mei, REN Xiao-Ping, HUANG Li, CHEN Wei-Gang, LI Zhen-Dong, ZHOU Xiao-Jing, CHEN Yu-Ning, LIAO Bo-Shou, JIANG Hui-Fang
Peanut is an allotetraploid crop with a large genome. The construction of genetic linkage map and QTL mapping of related traits has little progress for peanut. In the present study, a genetic map consisting of 20 linkage groups was constructed with 234 SSR markers based on 2653 published SSR markers by using the F2population derived from the cross between Fuchuan Dahuasheng and ICG6375. The genetic map covers 1683.43 cM, and the length of each linkage group varies from 36.11 to 131.48 cM, the number of markers in each linkage group varies from 6 to 15, with an average distance of 7.19 cM. Combining with the data of main stem height and number of total branches of F3 population in the environments of Wuhan and Yangluo, we performed QTL mapping and genetic effects analysis of QTLs by software WinQTLCart 2.5 using CIM (Composite Interval Mapping) method. As a result, 17 QTLs related to main stem height and number of total branches on eight linkage groups were detected with contribution percentage of 0.10%-10.22%. Comparing the QTLs detected in the environments of Wuhan and Yangluo, qMHA061.1 and qMHA062.1were in the same linkage region of markers TC1A2-AHGS0153 of linkage group LG06 with contribution percentage of 5.49%-8.95%, qMHA061.2 and qMHA062.2were between the markers AHGS1375 and PM377 on linkage group LG06 with contribution ratio of 2.93%-5.83%, qMHA092.2 and qMHA091.1 were in the same linkage region of the markers GM2839-EM87 in linkage group LG09 with contribution percentage of 0.53%-9.43%. The QTLs repeatedly detected are important for molecular breeding of peanut.
分子遗传图谱广泛应用于基因定位、图位克隆以及分子标记辅助育种。栽培种花生(Arachis hypogaea L.)为异源四倍体(2n=40), 因富含脂肪、蛋白质、白藜芦醇等营养保健成分而深受人们的喜爱, 是我国第二大油料作物[1, 2]。在早期的研究中, 由于栽培种花生的DNA多态性相对较为贫乏, 而野生花生的多态性丰富, 因此, 早期的花生遗传图谱的构建多以野生花生为材料[3, 4, 5, 6]。随着SSR标记的大量开发, 近十多年来的研究结果显示, 栽培种花生中也存在较为丰富的遗传多态性。栽培种花生的遗传图谱的构建不断完善, 重要性状QTL的研究报道不断增多。Varshney等[7]以栽培种花生的RIL群体为材料, 构建了1个包含135个SSR标记的连锁图, 定位了4个与抗干旱相关的QTL。Ravi等[8]在Varshney构建遗传连锁图谱的基础上增加了56个标记, 构建了包含191个标记的图谱, 定位了13个与干旱抗性成分相关的QTL。Khedikar等[9]利用栽培种花生的RIL群体构建了一张含有56个SSR标记涉及14个连锁群的遗传图谱, 定位到与抗锈病相关的11个分子标记。Shirasawa等[10]构建了一张含有1114个位点的栽培种花生遗传图谱, 获得了与开花期、分枝角度、主茎高、荚果长、荚果厚和荚果宽相关的QTL共9个。Wang等[11]利用栽培种花生的F2群体构建了一张包含318个标记的遗传图谱, 定位到15个与番茄斑萎病毒(TSWV)相关的QTL, 37个与叶斑病相关的QTL。张新友等[12]运用栽培种RIL群体和F2群体定位了与产量和品质性状相关的QTL 62个。综合分析栽培种花生遗传图谱和QTL的研究结果发现, 花生遗传图谱的构建还很不完善, 缺乏高密度的遗传图谱。花生的主茎高和总分枝数是重要的农艺性状, 很多研究结果都表明, 主茎高和总分枝数与产量性状之间存在显著的相关性[13, 14], 而且主茎高还与植株的抗倒伏性相关。本文以花生的主茎高和总分枝数为调查性状, 采用2个环境下的数据, 拟构建一张遗传图谱, 这对花生的分子设计育种具有重要意义。
根据核心种质的遗传多样性鉴定结果, 选择遗传差异较大的地方龙生型品种富川大花生为母本、从ICRISAT引进的珍珠豆型材料ICG 6375为父本配制杂交组合, 构建218个F2单株和218个F3家系的检测群体。
2012年将F2材料种植于湖北武汉中国农业科学院油料作物研究所试验农场, 单株收获。2013年将上年收获的每个单株材料的种子分成两等份, 分别在武汉中国农业科学院油料作物研究所试验农场和武汉阳逻基地种植成F3株系。单粒播种, 每行12株, 行长2.0 m, 种植密度为0.4 m× 0.2 m, 常规田间管理。收获时去掉每个株行的边株, 收获中间10株, 按《花生种质资源描述规范和数据标准》[15]考察主茎高和总分枝数。
选取幼嫩叶片, 采用改良CTAB法提取DNA, 用1%的琼脂糖凝胶电泳检测DNA浓度与纯度。所用引物序列来源于参考文献[16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33]。PCR体系为10 μ L, 含10 mmol L-1Tris-HCl (pH 8.3)、50 mmol L-1KCl、300~400 μ mol L-1 dNTPs、10~40 pmol L-1引物对、10~20 ng模板DNA。PCR程序为Touchdown, 扩增条件为94℃预变性3 min; 93℃变性30 s, 65℃退火30 s (每个循环-1℃), 72℃延伸1 min, 共10个循环; 93℃变性30 s, 55℃退火30 s, 72℃延伸1 min, 共20个循环; 72℃延伸10 min。PCR产物经6%变性聚丙烯酰胺凝胶电泳检测, 硝酸银染色, 显影, 扫描保存。
统计每份材料扩增的条带, 与母本相同的带型记为“ 1” , 与父本相同的带型记为“ 2” , 同时具有亲本带型的记为“ 3” , 缺失或者不清的带型记为“ 0” 。
运用JoinMap 3.0构建遗传图谱, 首先采用New Project构建新的工作文件夹, 对options内的参数进行设置, 设置LOD≥ 3, 步长设置为0.05; 选择右边的LOD grouping (tree)等选项; 选择population内的create groups for mapping, 即得相应连锁群。连锁分群时应用WinQTLCart2.5复合区间作图法进行QTL定位, 以LOD值≥ 2.5作为检测标准。QTL的命名以性状加所在连锁群加年份命令, 若同一连锁群上出现2个或以上QTL时, 则QTL后面加“ .” 表示, 比如qMHA061.2表示位于LG06连锁群在第1个环境下即武汉环境下检测到的第2个与主茎高相关的QTL。
利用已经公开发表的图谱上公布的SSR和EST-SSR引物[19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35]共2653对在双亲(富川大花生和ICG6375)中进行筛选, 获得多态性引物275对(表1), 占8.52%。将获得的多态性引物在F2代群体的218个单株中检测, 共检测到281个位点, 多态率为8.71%。其中AHGS1314、AHGS1470、AHGS1672、AHGS2429、TC6E1和GNB533等6对引物均扩增出了2个位点。
通过统计F2代群体各单株的带型, 在检测到的281个位点中, 符合孟德尔分离比例的标记共137个, 占48.76%。
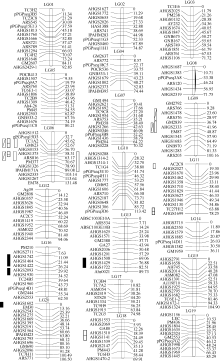
根据F2代群体中检测得到的281个多态性标记, 运用JoinMap 3.0软件进行遗传连锁分析, 获得包含20个连锁群的连锁图(图1), 各连锁群的信息列于表2。在281个多态性标记中, 有234个标记进入连锁群, 47个标记未进入连锁群。构建的图谱总长度为1683.43 cM, 连锁群长度在36.11~131.48 cM之间, 平均每个连锁群长度为84.17 cM; 每个连锁群的标记数在6~15个之间, 平均为11.7个; 相邻两两标记间的平均距离为7.19 cM, 图谱上偏分离标记共116个, 占49.57%。分析各连锁群的长度, 最长的为LG06, 达131.48 cM; 最短的为LG14, 仅36.11 cM。共有9个连锁群的长度> 84.17 cM, 包括LG01、LG09、LG10等, 其余11个连锁群的长度< 84.17 cM。比较分析各连锁群上的标记数, 发现除LG08和LG14这2个连锁群较少外(分别为6个和7个), 其余18个连锁群相对较多, 在9~15个之间。
表2结果还表明, 图谱上的相邻两两标记间的距离不同。相邻两两标记之间的距离≤ 10 cM的有162个, 占69.23%。相邻标记之间距离最小为1.61 cM, 位于LG06连锁群上的GM623与AHGS0153之间; 距离最大为 32.7 cM, 位于同一连锁群(LG06)上的AHGS1512与pPGPseq15D3之间。图谱上偏分离标记分布也不均匀。LG15连锁群上偏分离标记最多, 11个标记发生了偏分离, 占连锁群上标记数目的84.62%。LG04和LG18连锁群上的偏分离标记最少, 均只有2个标记发生了偏分离, 分别占22.22%和20%。
| 表1 引物多态性比例 Table 1 Percentage of polymorphic primers tested |
通过对亲本和218个F3家系的主茎高和总分枝数在武汉和阳逻环境下的鉴定, 获得的结果列于表3。从表3看出, 2个亲本在2种环境下的差异为20 cm左右, 总分枝数相差12条左右。武汉环境下F3的平均主茎高56.27 cm, 变异范围31.50~87.50 cm; 总分枝数17.66条, 变异范围4.00~61.00条; 主茎高和总分枝数的最高值与最低值间的差异均超出了双亲之间的差异。阳逻环境下F3的平均主茎高54.30 cm, 变异范围31.00~86.00 cm; 总分枝数18.04条, 变异范围4.30~55.40条; 阳逻环境下的主茎高和总分枝数的最高值与最低值间的差异也均超出了双亲之间的差异。统计分析不同主茎高范围的家系数和不同分枝数范围的家系数, 获得的分布图列于图2。从图2看出, 两种环境下的主茎高分布基本一致, 符合正态分布; 总分枝数的分布也基本一致, 呈偏正态分布。
| 表2 本研究开发的连锁群信息 Table 2 Information of the linkage groups developed in the present study |
 | 图1 花生遗传连锁图谱武汉环境下主茎高QTL; |
亲本以及后代群体性状变异
Variation of parents and populations traits
| 环境 Env | 性状 Trait | ICG 6375 (父本) | 富川大花生 Fuchuan Dahuasheng (母本) | 最大值 Max. | 最小值 Min. | 平均值 Mean | 标准差 SD | 偏度 Kurt | 峰度 Skew | 变异系数 CV (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| 武汉 Wuhan | 主茎高 Main stem height | 65.40 | 83.60 | 87.50 | 31.50 | 56.27 | 9.62 | 0.40 | 0.46 | 17.10 |
| 总分枝数 Number of total branches | 11.30 | 23.20 | 61.00 | 4.00 | 17.66 | 8.65 | 1.32 | 3.77 | 48.98 | |
| 阳逻 Yangluo | 主茎高 Main stem height | 62.40 | 89.30 | 86.00 | 31.00 | 54.30 | 11.44 | 0.27 | -0.44 | 21.06 |
| 总分枝数 Number of total branches | 10.80 | 26.60 | 55.40 | 4.30 | 18.04 | 9.39 | 1.06 | 1.14 | 52.07 |
武汉和阳逻环境下后代群体主茎高和总分枝数频次分布图 Common markers of the linkage groups developed from the present study and by Shirasawa (2013)本研究构建
的连锁群
Linkage group from the studyShirasawa (2013)
整合的连锁群
Linkage group by Shirasawa (2013)共有标记数
No. of common markers本研究构建的连锁群
Linkage group
from the studyShirasawa (2013)
整合的连锁群
Linkage group by Shirasawa (2013)共有标记数
No. of common markersLG01 A01 7 LG11 B01 10 LG02 A02 5 LG12 B02 8 LG03 A03 8 LG13 B03 9 LG04 A04 6 LG14 B04 6 LG05 A05 7 LG15 B05 11 LG06 A06 9 LG16 B06 10 LG07 A07 10 LG17 B07 4 LG08 A08 4 LG18 B08 8 LG09 A09 8 LG19 B09 10 LG10 A10 9 LG20 B10 13
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|