
{kind=link}
{kind=link}
{kind=link}
{kind=link}
稻-麦轮作下9000 kg hm-2产量水平扬麦20的群体质量及花后光合特征
[丁锦峰 , 黄正金, 袁毅, 朱新开, 李春燕, 彭永欣, 郭文善
, 黄正金, 袁毅, 朱新开, 李春燕, 彭永欣, 郭文善* ]
, 黄正金, 袁毅, 朱新开, 李春燕, 彭永欣, 郭文善]
|
|


第一作者联系方式: E-mail: jfdin@yzu.edu.cn
在稻麦两熟制条件下, 于2010—2011和2011—2012年度, 通过氮肥施用量、施用时期及比例的调控, 建立了扬麦20不同产量水平群体, 比较≥9000 kg hm-2群体(P1)与<9000 kg hm-2群体(P2)的产量构成、群体质量及花后光合特征, 为稻茬小麦大面积高产提供理论依据和技术支撑。P1较P2群体产量高约10%, 每穗粒数高约5.5%, 差异显著, 但两群体的穗数和千粒重差异不显著。P1群体的穗数、穗粒数和千粒重分别为482~538万 hm-2、47~49粒和34~39 g。籽粒产量与孕穗和开花期叶面积指数(LAI)及花后LAI衰减率和光合势呈抛物线关系, 与乳熟期LAI、粒(重)/叶比和群体生长率呈显著线性正相关。两年度试验结果表明P1群体具有以下特征, 孕穗期、开花期和乳熟期的LAI分别为6.5~7.0、5.0~6.0和4.0~4.5; 粒(数)/叶比为0.37~0.39, 粒(重)/叶比为13.5~14.5; 花后LAI衰减率、群体生长率、光合势和净同化率分别为0.13~0.15 d-1、19~20 g m-2 d-1、103×104~118×104 m2 d hm-2和9~11 g m-2d-1。花后21 d和28 d, 剑叶SPAD值、净光合速率、丙二醛(MDA)含量及3种抗氧化酶(CAT、POD和SOD)活性与产量相关性均达极显著水平。研究表明, 稻-麦轮作体系中, 扬麦20达到9000 kg hm-2产量水平的栽培技术关键点是获得适宜穗数的基础上, 主攻每穗粒数与千粒重的协调增加, 使群体在花前具有较高的光合面积和光合速率, 花后光合面积衰减速率低, 维持较高的光合面积, 从而充分积累花后光合物质, 在适宜库容基础上保障对库充实的需求。
The objective of this study was to fine-tune cultivation techniques for large-scale high yielding ( ≥9000 kg ha-1) wheat production in rice-wheat rotation system. Wheat ( Triticum aestivumL. cv. Yangmai 20) populations with yield level ≥9000 kg ha-1 (P1) and <9000 kg ha-1 (P2) were established through managing application amount, timing and splitting ratio of nitrogen fertilizer in the 2010-2011 and 2011-2012 growing seasons. The grain yield and grain number per spike in P1 population were 10.0% and 5.5% higher than those of P2 population with significant difference between each other. However, the differences in spike number and 1000-grain weight between P1 and P2 were not significant. The yield components of P1 population were 4.82-5.38 million spikes per hectare, 47-49 grains per spike, and 34-39 g of 1000-grain weight. Grain yield of different populations showed parabolic correlations with leaf area index (LAI) at booting and anthesis stages, leaf area decreasing rate and photosynthetic potential after anthesis, as well as significantly linear correlations with LAI at milk-ripening stage, grain weigh per square centimeter leaf and population growth rate. According to the experiments results in two years, P1 population was characterized with dynamic LAI of 6.5-7.0 at booting, 5.0-6.0 at anthesis and 4.0-4.5 at milk-ripening stage; grain/leaf ratio of 0.37-0.39 grains cm-2 leaf and 13.50-14.50 mg grain cm-2 leaf; LAI decrease rate of 0.13-0.15 d-1 after anthesis; population growth rate of 19-20 g m-2 d-1 after anthesis; photosynthetic potential of 1.03-1.18 million m2 d ha-1 after anthesis; and net assimilation of 9-11 g m-2 d-1 after anthesis. At the stages of 21 and 28 days after anthesis, the SPAD reading, net photosynthetic rate, malondialdehyde content, and activities of antioxidant enzymes (CAT, POD, and SOD) in flag leaves were significantly correlated with grain yield. Our results suggest the following key points of cultivar potential for 9000 kg ha-1 yield potential of Yangmai 20 in rice-wheat rotation system are suitable population spike number as the basis; synchronous increases of grain number per spike and 1000-grain weight; high photosynthetic area and photosynthetic rate before anthesis; slow decrease of photosynthetic area after anthesis with its maintainable high level, and an abundant accumulated assimilate meeting to the sink enrichment.
中国稻茬小麦种植面积9× 106~13× 106 hm2, 约占耕地的10%[1, 2, 3, 4]。长江中下游作为稻茬小麦主要种植区, 小麦种植面积和总产均超全国的12%[5], 其中稻茬小麦面积8.5× 106 hm2 [6]。近年来已报道[7]该区稻茬小麦高产创建中单产达9000 kg hm-2, 但高产稳产性亟待提高。究其原因主要是对稻茬小麦9000 kg hm-2高产群体生长发育特性、群体诊断指标和养分吸收规律尚不十分清楚, 关键栽培技术尚需进一步研究论证。研究表明, 不同产量水平群体产量结构及动态特征存在较大差别[8]。于振文等[9]根据黄淮麦区的生态特点, 提出旱茬小麦6000~7500 kg hm-2向9000 kg hm-2高产发展需要建立在合理群体结构的基础上, 并提出具有9000 kg hm-2高产潜力不同穗型品种的合理群体结构及产量构成主攻目标, 及旱茬小麦延长缓衰期、缩短速衰期、保持较长光合速率高值持续期, 同步提高生物产量和经济系数的高产途径。郝代成等[10]和高海涛等[11]研究表明, 黄淮麦区9000 kg hm-2高产旱茬小麦剑叶在展开前中期具有较高的光合速率, 后期下降缓慢, 延长了光合时间。前人关于稻茬小麦产量结构及衰老特征的研究以籽粒产量高于7500 kg hm-2为主[12, 13], 对9000 kg hm-2高产群体的产量结构及衰老特征少有涉及。本试验在稻麦两熟制条件下, 通过氮肥施用量、施用时期及比例的调控, 建立不同产量水平群体, 分析其产量构成、主要生育时期叶面积指数(LAI)、粒/叶比和花后光合特征及衰老相关酶活性差异及其与产量的关系, 探讨9000 kg hm-2高产稻茬小麦的群体产量结构、群体质量及其花后光合特征, 提出相关特征指标, 为稻茬小麦大面积高产栽培提供理论依据和技术支撑。
2010— 2011和2011— 2012年度, 在扬州大学江苏省作物遗传生理重点实验室试验场(119° 42′ N, 32° 39′ E)进行田间试验。该地属亚热带湿润气候区, 年平均温度13.2~16.0℃、降雨量800~1200 mm、日照2000~2600 h、无霜期220~240 d。试验年度气候条件见表1。试验田前茬为水稻(产量10 000 kg hm-2), 土壤为轻壤土。2010— 2011年度小麦播种前0~20 cm土层含水解氮65.81 mg kg-1、速效磷45.88 mg kg-1、速效钾101.98 mg kg-1、有机质15.5 g kg-1; 2011— 2012年度小麦播种前0~20 cm土层含水解氮65.56 mg kg-1、速效磷46.23 mg kg-1、速效钾102.05 mg kg-1、有机质15.9 g kg-1。供试品种为扬麦20, 由江苏里下河地区农业科学研究所提供。
采用3因素裂区设计, 共16个处理, 小区面积为18 m2, 3次重复。以施氮量(纯氮)为主区, 设2个水平, 即210 kg hm-2和262.5 kg hm-2; 以施氮比例(基肥∶ 壮蘖肥∶ 拔节肥∶ 穗肥)为副区, 设2个水平, 分别是3∶ 1∶ 3∶ 3和5∶ 1∶ 2∶ 2; 以穗肥追氮时期为裂区, 设剑叶露尖、孕穗期、抽穗期和开花期4个追氮时期。播种前施用基肥, 四叶期施用壮蘖肥, 叶龄余数2.5时施用拔节肥; 磷肥(P2O5)及钾肥(K2O)均为130 kg hm-2, 50%作基肥, 50%作拔节肥。两年度均于11月1日, 人工条播, 基本苗为225万株 hm-2, 行距30 cm, 三叶期定苗。其余管理措施同大田高产栽培。
1.3.1 LAI和干物质积累量 分别于孕穗期、开花期、乳熟期和成熟期从每小区取0.15 m2样品, 植株分器官, 用长宽系数法测定绿叶面积后, 于105℃杀青1 h, 80℃烘至恒重, 测定干物重。LAI=取样绿叶面积(cm2)/取样面积(m2)× 10-4(m2 cm-2)。
1.3.2 SPAD值 于小麦开花期及花后每隔7 d, 从各小区随机取长势均匀单茎的剑叶, 采用SPAD-502型叶绿素计(KONICA MINOLTA, 日本)测定SPAD值, 每个小区重复10次。
| 表1 小麦不同生育阶段的积温、日照和降雨 Table 1 Accumulated temperatures, sunshine, and precipitation during various phases of wheat growth |
1.3.3 净光合速率(Pn) 于小麦开花期及花后每隔7 d, 从各小区随机取长势均匀单茎的剑叶, 采用LI-6400便携式光合仪(LI-COR公司, 美国, 人工光源900 μ mol m-2 s-1), 于晴天9:30— 11:30测定Pn, 每个小区重复5次。
1.3.4 过氧化物酶(POD)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)活性及丙二醛(MDA)含量 于小麦开花期及花后每隔7 d取20片长势均匀单茎的剑叶, 冰盒储存, 液氮速冻, -20℃冰箱保存。将冷藏鲜样擦净去叶中脉, 剪碎混匀称取0.50 g, 加5 mL磷酸缓冲溶液(pH 7.8)及少量石英砂, 冰浴研磨至匀浆, 4℃离心20 min (18 000× g), 上清液即为酶提取液。用紫外吸收法[14]测定POD及CAT活性, NBT光化还原法[14]测定SOD活性。采用硫代巴比妥酸法[14]测定MDA含量。3次重复, 取平均值。
1.3.5 产量及产量结构 成熟期分小区调查穗数、每穗粒数及千粒重, 收获1.2 m2计产, 3次重复, 折算成籽粒含水率为13%时的产量。
粒(数)/叶比=总结实粒数/孕穗期叶面积(cm2); 粒(重)/叶比=籽粒产量(mg)/孕穗期叶面积(cm2)。
叶面积衰减率(LAI d-1) = (LAI2 - LAI1) / (t2 - t1)。
群体生长率(g m-2 d-1) = (W2 - Wl) / (t2 - t1)。
光合势(× 104 m2 d hm-2) = 1/2 (LAI1 + LAI2) × (t2 - t1)。
净同化率(g m-2 d-1) = [(ln LAI2 - ln LAI1) / (LAI2 - LAI1)] × [(W2 - W1) / (t2 - t1)]。
式中, LAI1和LAI2为前后两次测定的叶面积指数, W1和W2为前后2次测定的干物质重, t1和t2为前后2次测定的时间。
以产量9000 kg hm-2为限, 将各小区分成两类, 分别构成P1 (≥ 9000 kg hm-2)和P2 (< 9000 kg hm-2)群体, 2011— 2012年度有4个小区出现倒伏, 分析时剔除。
采用SPSS 15.0软件进行方差分析(ANOVA), 用LSD法检验差异显著性。采用y = ax + b和y =ax2 + bx + c两种方程拟合主要生育时期LAI、粒叶比及花后叶面积衰减率、群体生长率、光合势和净同化率与产量的关系。用SigmaPlot 10.0软件绘图。
方差分析结果表明, 因气候条件差异, 不同施氮量对产量的影响在年度间有差异; 而施氮比例及追氮时期对籽粒产量的影响两年表现一致, 处理间存在极显著差异; 处理与产量的互作效应在年度间表现有所差异(表2)。综合2年结果, 在氮肥适当后移(3∶ 1∶ 3∶ 3)条件下, 穗肥适当早施(剑叶露尖、孕穗期)产量较高; 高施氮量(HN, 262.5 kg hm-2)的增产效果不明显。在施氮量210.0 kg hm-2 (LN)、氮肥运筹3∶ 1∶ 3∶ 3 (S3133)、剑叶露尖追氮处理(T1)下两年产量均高于9000 kg hm-2 (表3)。
两年度所有小区P1与P2的籽粒产量差异显著, P1较P2高10.31% (2010— 2011)和9.12% (2011— 2012); 产量构成三因素中, 两群体的穗数差异不大, 但穗粒数(P1较P2两年度分别高5.96%和5.01%)和千粒重(P1较P2两年度分别高1.69%和4.23%)都有显著差异(表4)。相关分析表明, 产量与穗粒数呈显著正相关(r2010-2011=0.66, P< 0.01; r2011-2012=0.84, P< 0.05), 与千粒重呈正相关(r2010-2011=0.45, P> 0.05; r2011-2012=0.66, P< 0.05), 而与穗数只微弱负相关宜穗数的基础上, 主攻每穗粒数与千粒重的协调增加。本试验条件下, P2群体的产量指标为穗数482~538万穗 hm-2、每穗47~49粒、千粒重34~39 g。
| 表2 不同处理籽粒产量的方差分析 Table 2 ANOVA of yield in different treatments |
| 表3 不同处理下小麦籽粒产量 Table 3 Grain yield of wheat under different treatments (kg hm-2) |
| 表4 不同产量水平群体的产量及其构成 Table 4 Grain yield and yield components of different yield level groups |
(r2010-2011= -0.14, P> 0.05; r2011-2012= -0.39, P> 0.05)。说明稻茬小麦9000 kg hm-2高产的栽培关键应在获得适
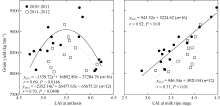
不同产量水平群体的LAI在孕穗期及开花期差异不显著; 而乳熟期P1较P2群体高25.26%和9.00%, 差异显著(表5)。孕穗期LAI与产量呈抛物线关系(r2010-2011=0.66, P< 0.05; r2011-2012=0.19, P> 0.05); 开花期LAI与产量呈抛物线关系, 乳熟期LAI与产量呈线性正相关(图1)。年度间两群体的LAI在孕穗期、开花期及乳熟期差异未达显著水平。综合两年度试验结果, 在保证孕穗期和开花期适当LAI的基础上, P1群体乳熟期高效功能叶片衰老速率明显较缓慢, 其孕穗期、开花期和乳熟期的LAI分别为6.5~7.0、5.0~6.0和4.0~4.5。
| 表5 不同产量水平群体的主要生育时期叶面积指数(LAI)及粒/叶比 Table 5 LAI at the main growth stages and grain-leaf ratio of different yield level groups |
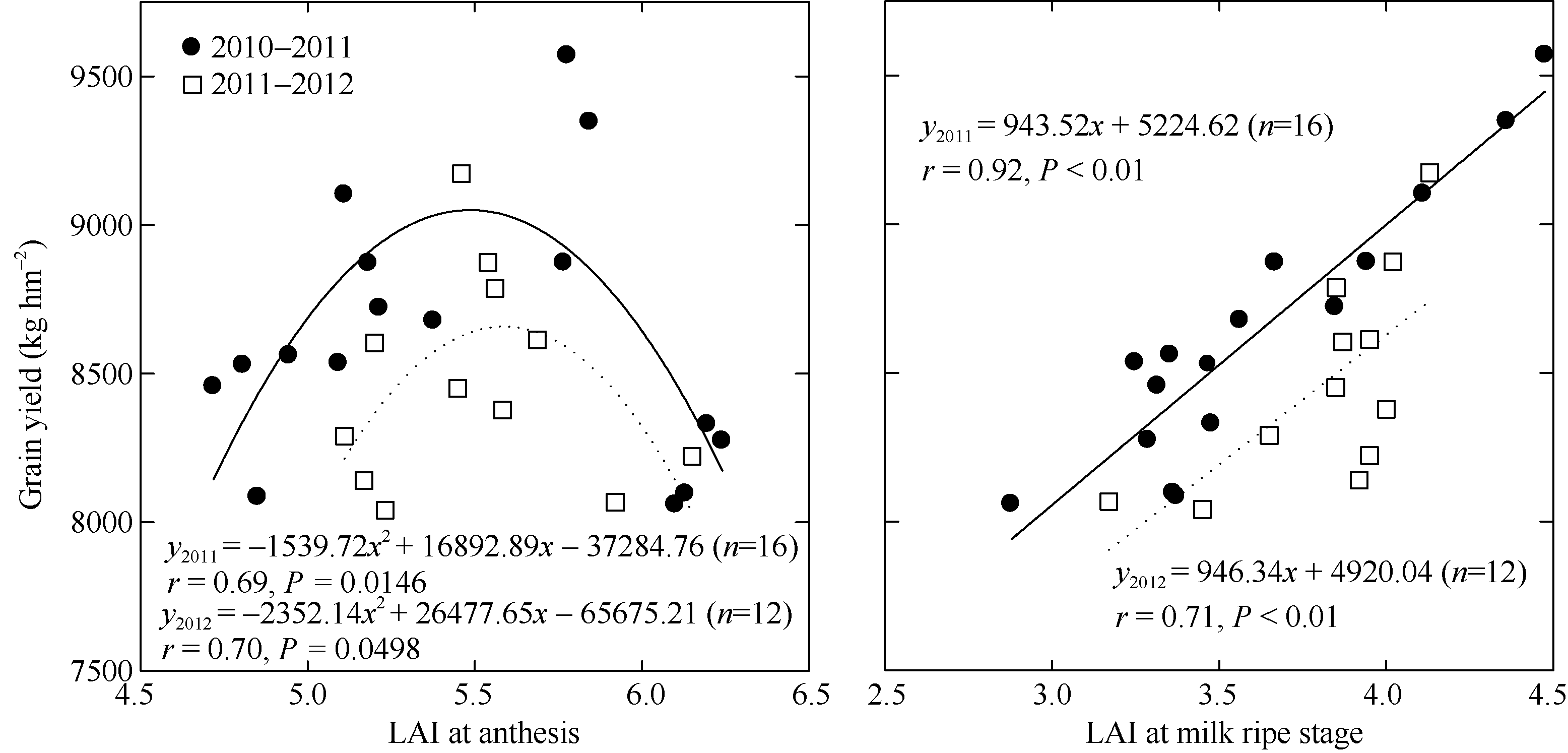 | 图1 开花期和乳熟期LAI与籽粒产量的关系Fig. 1 Relationship between LAI at the stages of anthesis and milk ripe and grain yield |
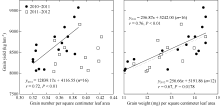
P1和P2群体的粒(数)/叶比无显著差异, 而粒(重)/叶比只在2010— 2011年度有显著差异, P1群体较P2群体高11.65% (表5)。粒(数)/叶比与籽粒产量在2010— 2011年度呈显著线性正相关, 2011— 2012年度呈散点分布; 粒(重)/叶比与籽粒产量呈显著线性正相关(图2)。总体来看, P2群体具有较高的源库协调水平, 关键是在适宜库容基础上提高源对库充实的供应能力; 其粒(数)/叶比为0.37~0.39 粒 cm-2叶, 粒(重)/叶比为13.5~14.5 mg cm-2叶。
 | 图2 粒叶比与籽粒产量的关系Fig. 2 Relationship between grain-leaf ratio and grain yield |
P1和P2开花期至成熟期叶面积衰减率、光合势及净同化率差异均未达显著水平, 但P1的群体生长率显著高于P2, 差异达显著水平, 两年度分别高18.44%和14.42% (表6)。花后叶面积衰减率及净光合势与籽粒产量均呈抛物线关系, 群体生长率与籽粒产量呈线性正相关, 均达显著水平; 净同化率与籽粒产量呈显著抛物线(2010— 2011)或显著线性正相关(2011— 2012)。年度间群体生长率及光合势差异均未达显著水平(图3)。两年度试验结果表明, P1群体花后LAI衰减率为0.13~0.15 d-1, 群体生长率19~20 g m-2 d-1, 光合势103× 104~118× 104 m2 d hm-2, 净同化率9~11 g m-2 d-1。
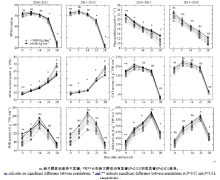
花后旗叶SPAD值、POD活性、CAT活性、POD活性均呈单峰曲线变化, 峰值期分别出现在花后7、14、7及21 d; 净光合速率逐渐下降; MDA含量逐渐上升, 开花期至花后14 d缓慢增加。P1群体花后旗叶SPAD值、净光合速率及CAT、POD和SOD酶活性均高于P2群体, 而花后剑叶MDA含量低于P2群体(图4)。花后21 d和28 d.的旗叶SPAD值和SOD酶活性以及花后7~28 d的MDA含量、SOD酶活性均存在显著差异; 年度间花后旗叶SPAD值、MDA含量及CAT酶活性差异均达显著水平, 但净光合速率、POD和SOD酶活性差异均未达显著水平(图4)。相关性分析表明, 花后21 d和28 d旗叶SPAD值、净光合速率、MDA含量及CAT、POD和SOD酶活性与籽粒产量相关性均达极显著水平(表7)。说明随植株花后剑叶光合能力和抗衰老能力的逐步增强籽粒产量提高, 在籽粒灌浆后期表现更为明显。
| 表6 不同产量水平群体的花后LAI衰减率、群体生长率、光合势和净同化率 Table 6 LAI decrease rate, rate of population growth, photosynthetic potential, and net assimilation after anthesis of different yield level groups |
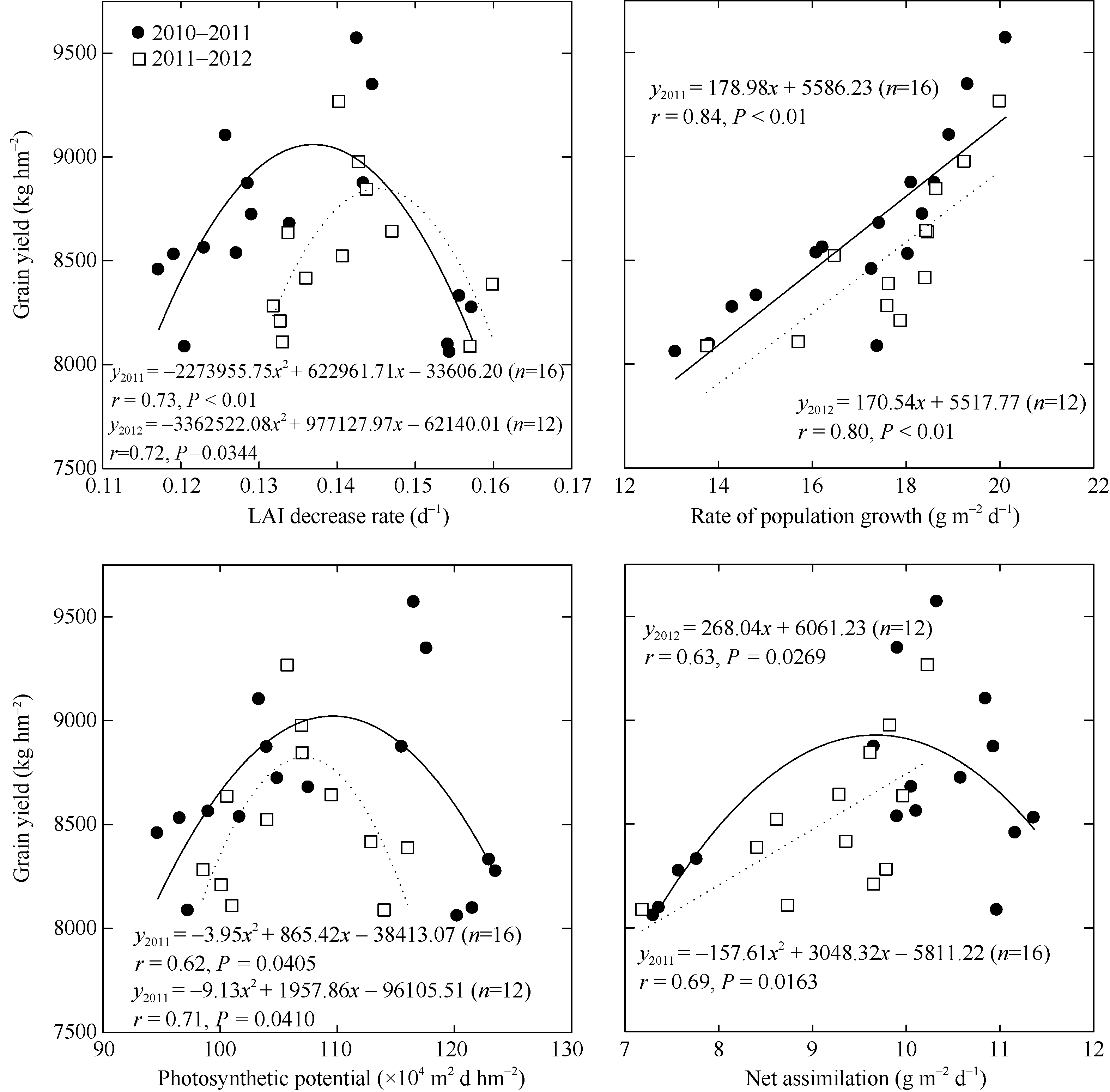 | 图3 花后LAI衰减率、群体生长率、光合势和净同化率与籽粒产量的关系Fig. 3 Relationships of yield with LAI decrease rate, rate of population growth, photosynthetic potential, and net assimilation after anthesis |
 | 图4 不同产量水平群体的花后剑叶叶绿素含量、净光合速率和抗氧化相关酶活性变化Fig. 4 Chlorophyll content, net photosynthetic rate and antioxidation relative enzyme activity in flag leaves after anthesis of different yield level groups |
研究表明, 长江中下游稻茬麦区适宜播期为10月底至11月初[15, 16]。本试验的播期为11月1日, 根据本课题组2010— 2011和2011— 2012年度同一大田下稻茬小麦晚播(11月20日)试验结果, 无小区能达到9000 kg hm-2产量水平, 说明稻茬小麦高产栽培需要播期适宜。稻-麦轮作体系下, 刘萍等[17]提出, 扬麦12的适宜密度为基本苗130~170万株 hm-2, 产量可达8000 kg hm-2, 本试验的基本苗高于这一标准, 为225万株 hm-2。朱新开等[18]认为扬麦10号施氮量在180~240 kg hm-2范围内, 最优施氮比例(基肥∶ 壮蘖肥∶ 拔节肥:孕穗肥)为3∶ 1∶ 3∶ 3, 其次为5∶ 1∶ 2∶ 2; 田纪春等[19]认为, 旱茬小麦在施氮量250 kg hm-2条件下, 基肥∶ 越冬肥∶ 追肥为4∶ 2∶ 4, 孕穗期追肥籽粒产量最高。本试验结果表明, 在氮肥适当后移(3∶ 1∶ 3∶ 3)条件下, 穗肥适当早施(剑叶露尖、孕穗期)产量较高; 高施氮量(262.5 kg hm-2)的增产效果不明显。在施氮量210.0 kg hm-2、氮肥运筹3∶ 1∶ 3∶ 3、剑叶露尖追氮处理下两年产量均高于9000 kg hm-2。
| 表7 籽粒产量与花后剑叶生理指标的相关性 Table 7 Correlations between grain yield and post-anthesis physiological characteristics of flag leaf |
综合前人研究结论和本试验结果, 我们认为长江中下游流域稻-麦轮作制下扬麦20达到9000 kg hm-2高产水平的关键栽培技术应包括11月1日前后播种, 基本苗225万 hm-2, 条播, 行距30 cm, 2~3 cm播深; 氮肥施用量210 kg hm-2, 磷、钾肥施用量均130 kg hm-2, 氮肥施用比例(基肥∶ 壮蘖肥∶ 拔节肥∶ 穗肥) 3∶ 1∶ 3∶ 3, 基肥于播种前施用, 壮蘖肥于四叶期施用, 拔节肥于叶龄余数2.5时施用, 穗肥于剑叶露尖或孕穗期施用, 磷钾肥50%基施, 50%于叶龄余数2.5时追施。其他小麦品种达9000 kg hm-2的关键技术有待进一步试验完善。
不同生态区域小麦产量≥ 9000 kg hm-2群体的产量结构存在差异, 于振文等[9]提出适于黄淮麦区旱茬中穗型小麦品种的群体质量指标为穗数750万 hm-2、穗粒数35粒、千粒重45 g左右; 杜永等[20]建议的群体产量构成为穗数620万 hm-2左右、穗粒数≥ 33、千粒重≥ 42 g。本试验地处江苏淮南麦区, 结果表明稻茬小麦产量≥ 9000 kg hm-2群体穗数、每穗粒数和千粒重分别为482~538万 hm-2、47~49粒和34~39 g, 相比于黄淮麦区旱茬及稻茬小麦产量≥ 9000 kg hm-2群体表现出穗数低、每穗粒数高、千粒重低的特征。相同生态区域下, 朱新开等[12]研究提出稻茬小麦7500 kg hm-2高产群体穗数、每穗粒数和千粒重, 本试验9000 kg hm-2高产群体较之, 穗数高60万 hm-2, 每穗粒数高8粒, 千粒重低4 g左右。表明稻茬小麦实现公顷产9000 kg关键在于适宜穗数的基础上, 协调增加每穗粒数与千粒重。
适宜的LAI是小麦群体质量的基础指标[8]。张洪程等[21]明确了黄淮麦区河南旱茬小麦产量≥ 9000 kg hm-2群体孕穗期LAI约7.3、抽穗后20 d LAI约4.2; 朱新开等[12]研究认为稻茬小麦8000 kg hm-2高产群体LAI在孕穗期为6.0~6.5、开花期为5.0~6.0、乳熟期约3.4。本试验结果表明, 稻茬小麦产量≥ 9000 kg hm-2群体孕穗期、开花期和乳熟期的LAI分别为6.5~7.0、5.0~6.0和4.0~4.5, 相比于旱茬小麦产量≥ 9000 kg hm-2群体, 孕穗期和开花期LAI略低, 乳熟期LAI接近; 相比同区域稻茬小麦8000 kg hm-2高产群体, 本试验产量≥ 9000 kg hm-2群体孕穗期适宜LAI较高, 花后可维持更高的LAI。
粒叶比是衡量小麦群体库源协调水平的综合指标, 高产群体需要源库关系在高水平上取得动态平衡[8, 22]。郭文善等[22]认为群体最高LAI相近, 产量随粒叶比的提高相应增加, 最大LAI为7.5左右时, 粒(数)/叶比为0.298, 粒(重)/叶比为11.665时产量达8700 kg hm-2。封超年等[23]认为粒叶比高的群体花后光合面积衰减慢、叶片活性氧消除能力强、光合功能期长, 有利于提高灌浆速率和粒重。慕美财等[24]认为旱茬小麦产量≥ 9000 kg hm-2群体在增源、扩库和畅流等方面具有优势, 在增加光合源方面更具优势。本试验结果表明, 产量≥ 9000 kg hm-2群体最高LAI适宜值在6.5~7.0, 粒(数)/叶比为0.37~0.39, 粒(重)/叶比为13.50~14.50, 说明具有较高的源库协调水平, 关键是在适宜库容基础上提高源对库充实的供应能力。
结合稻茬小麦9000 kg hm-2群体指标如茎蘖动态[25]、花后干物质积累量[26]的研究结果, 可以看出, 稻茬小麦实现9000 kg hm-2产量水平要比一般高产群体发展动态更为合理, 群体数量稳中有升; 群体质量尤其是孕穗开花后优化提高, 花后群体生产力更具优势, 源库协调水平有所加强。
产量≥ 9000 kg hm-2群体花后衰老特征已有较多的研究报道, 岳寿松等[27]认为黄淮麦区旱茬小麦产量≥ 9000 kg hm-2群体较一般高产群体, 剑叶衰老初期可溶性蛋白质含量高, 叶片衰老期间细胞膜质过氧化水平低; 郝代成等[10]认为产量≥ 9000 kg hm-2群体在灌浆中后期能保持较高的剑叶净光合速率, 灌浆后期下降缓慢, 越到后期优势越明显。本试验结果表明, 产量≥ 9000 kg hm-2群体不同生育时期剑叶SPAD值、净光合速率及活性氧保护酶(POD、CAT和SOD)活性均高于产量< 9000 kg hm-2群体, 而MDA含量均低于产量< 9000 kg hm-2群体。
已有大量研究试图解析小麦单叶及群体光合生产能力与产量的关系, 但其结论不尽相同[28, 29, 30]。本研究结果表明, 花后叶面积衰减率和光合势与产量呈抛物线关系; 群体生长率与产量呈显著线性正相关; 净同化率与籽粒产量呈抛物线或线性显著正相关。产量≥ 9000 kg hm-2群体花后生理指标为LAI衰减率0.13~0.15 d-1、群体生长率19~20 g m-2 d-1、光合势103~118万 m2 d hm-2、净同化率 9~11 g m-2 d-1。结果还表明, 稻茬小麦花后21 d及28 d旗叶SPAD值、净光合速率及活性氧保护酶活性与籽粒产量均呈线性显著正相关, MDA含量与籽粒产量呈线性显著负相关。说明提高稻茬小麦灌浆成熟期剑叶光合及抗衰性能有利于籽粒产量的提高, 这与黄淮旱茬小麦9000 kg hm-2群体较一般高产群体具有剑叶衰老期间抗衰性强, 后期光合优势明显的特点基本一致。小麦高产研究针对的是群体范畴, 植株剑叶的光合衰老特性一定程度上反映了群体冠层的光合状况, 但对于稻茬小麦9000 kg hm-2高产群体冠层光合特性仍有待明确。
稻-麦轮作下9000 kg hm-2产量水平扬麦20的穗数、穗粒数和千粒重分别为482~538万 hm-2、47~49粒和34~39 g; 孕穗期、开花期和乳熟期的LAI分别为6.5~7.0、5.0~6.0和4.0~4.5, 粒(数)/叶比为0.37~0.39, 粒(重)/叶比为13.5~14.5; 花后LAI衰减率、群体生长率、光合势和净同化率分别为0.13~0.15 d-1、19~20 g m-2 d-1、103× 104~118× 104 m2 d hm-2和9~11 g m-2 d-1。其栽培技术关键点是获得适宜穗数的基础上, 主攻每穗粒数与千粒重的协调增加, 使群体在花前具有较高的光合面积和光合速率, 花后光合面积衰减速率慢, 维持较高的光合面积, 从而充分积累花后光合物质, 在适宜库容基础上保障对库充实的需求。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|