

{kind=link}
{kind=link}
套作大豆苗期茎秆木质素合成与抗倒性的关系
[邹俊林1  , 刘卫国
, 刘卫国1, * , 袁晋1 , 蒋涛2 , 叶素琴1 , 邓榆川1 , 杨晨雨1 , 罗玲1 , 杨文钰1, * ]
, 刘卫国, 袁晋]
|
|


第一作者联系方式: E-mail: jlzou9@163.com
为从茎秆强度的角度探索套作大豆苗期耐阴抗倒机制, 对套作大豆苗期茎秆木质素合成与抗倒性的关系进行了研究。采用耐阴性不同的3个大豆品种(系), 在大豆-玉米套作和大豆单作两种种植模式下, 测定茎秆的木质素含量及其合成过程中的苯丙氨酸转氨酶(PAL)、4-香豆酸: CoA连接酶(4CL)、肉桂醇脱氢酶(CAD)、过氧化物酶(POD)等关键酶活性以及茎秆抗折力和抗倒伏指数。结果表明, 套作大豆苗期倒伏严重, 茎秆抗折力、抗倒伏指数、木质素含量和相关酶活性均显著低于单作。不同大豆品种受套作荫蔽影响程度不同, 强耐阴性大豆南豆12茎秆抗折力降低幅度最小, 在套作环境下其茎秆抗折力、抗倒伏指数大, 茎秆木质素含量高, PAL、4CL、CAD、POD活性强。相关分析表明, 套作大豆苗期茎秆木质素含量与抗折力极显著正相关( r = 0.890, P < 0.01), 与倒伏率极显著负相关( r = -0.889, P < 0.01), 与4CL、CAD酶活性显著正相关。套作环境下, 强耐阴性大豆苗期茎秆中较高的4CL、CAD活性是其维持高木质素含量的酶学基础, 而高木质素含量有利于提高茎秆强度, 进而增强其抗倒伏能力。
, LIU Wei-Guo, YUAN Jin
Stem physical characteristics are closely related to the crop lodging, increasing stem strength is available to resolve the problem of lodging. To explore the effect of intercropping soybean stem strength on the mechanism of shade tolerance and lodging resistance, we studied the relationship between lignin synthesis and lodging resistance. Three soybean cultivars with different shade tolerances were planted in maize-soybean intercropping and soybean monocropping, respectively. The lignin content, activities of phenylalanine ammonia-lyase (PAL), 4-coumarate:CoA ligase (4CL), cinnamyl alcohol dehydrogenase (CAD), peroxidase (POD) which involved in lignin synthesis in stem as well as the snapping resistance and lodging resistance index were determined at seedlings stage. The results showed that the intercropping soybean lodged seriously, the snapping resistance, lodging resistance index, lignin content and activities of the four enzymes were significantly lower than those of monocropping soybean at seedlings stage. The three soybean cultivars were affected differently by intercropping condition, the reduction of snapping resistance of Nandou 12 with strong shade tolerance was the least, its lodging resistance index, lignin content and activities of the four enzymes (PAL, 4CL, CAD, POD) also maintained a relatively high level. The lignin content of intercropping soybean seedling stem was significantly positively correlated with the snapping resistance ( r = 0.890, P< 0.01) and the activities of 4CL, CAD, and significantly negatively correlated with the actual lodging percentage ( r = -0.889, P < 0.01). In a word, the high activities of 4CL and CAD in the soybean stem provide a enzymatic basis to maintain higher lignin content, resulting in higher stem strength and higher lodging resistance in intercropping condition.
近年来, 在我国南方地区大面积推广大豆-玉米套作种植模式, 扩大了大豆种植面积, 增加了我国大豆供给, 成为振兴我国大豆产业的新途径[1]。生产中, 苗期倒伏是制约套作大豆高产和优质的主要因素之一, 严重影响大豆产量和机械化收获。在大豆-玉米套作模式中, 大豆苗期与玉米共生, 受玉米遮阴而处于弱光照强度和低红光与远红光比值的光照环境, 导致植株茎秆变细, 节间过度伸长, 初花期前极易发生藤蔓化和倒伏[2, 3, 4, 5]。茎秆性状作为影响大豆倒伏的最主要因素, 其形态生理、生物力学等特征与抗倒性密切相关, 而茎秆机械强度是提高大豆抗倒伏能力的关键因素[6, 7, 8, 9]。木质素作为细胞壁主要组成成分之一, 对维持茎秆机械强度具有明显的作用[10, 11]。研究表明, 大豆茎秆抗倒株型具有较高的木质素含量和茎秆机械强度[12, 13]; 对小麦、油菜、水稻等作物研究同样发现木质素含量高的品种抗倒伏能力强[14, 15, 16]。但也有研究表明, 不同抗倒伏能力品种间木质素含量无明显差异; 有些倒伏品种的木质素含量甚至高于不倒伏品种。
目前, 针对套作模式下大豆苗期抗倒伏性研究还比较少, 且集中于施肥和调节剂对茎秆抗倒伏能力的调控效应, 而对此模式下大豆苗期茎秆抗倒伏能力及其差异形成的生理基础尚不清楚。本研究测定大豆-玉米模式下苗期茎秆抗折力、抗倒伏指数、木质素含量及其合成相关酶活性, 探讨木质素合成机制与茎秆抗倒伏能力的关系, 旨在为套作大豆的高产栽培和抗倒品种选育提供理论依据。
从2010年30份经耐阴性鉴定的大豆品种(系)中[2]选取3个耐阴性差异明显的大豆品种(系), 即B1 (南豆032-4, 弱耐阴性, 四川省南充市农业科学院育种材料); B2 (九月黄, 中度耐阴性, 四川省阆中市地方品种); B3 (南豆12, 强耐阴性, 西南地区套作大豆主推品种, 四川省南充市农业科学院选育)。选用半紧凑型玉米品种川单418。
2012年和2013年在四川农业大学雅安校区教学科研园区进行试验。采用裂区设计, 主区为大豆-玉米套作和大豆单作2种种植模式, 副区为3个大豆品种(系), 副区面积为24 m2, 重复3次。套作模式中, 采用宽窄行种植玉米, 窄行行距40 cm, 宽行行距160 cm, 穴距35 cm, 每穴2株; 分别于2012年6月12日和2013年6月13日于玉米宽行内播种大豆, 每幅种2行, 行距50 cm, 穴距10 cm, 与玉米行相隔55 cm, 每穴定苗1株。单作模式中, 大豆行距50 cm, 穴距10 cm, 每穴定苗1株。施肥和其他管理措施同大田生产。
自各小区播种大豆后40 d (即套作大豆生育期为V6, 单作大豆生育期为V7)开始调查取样。选取长势一致的植株20株, 其中10株迅速截取茎秆第一节间, 经液氮处理后于-80℃冰箱中保存, 用于茎秆木质素含量和苯丙氨酸转氨酶(PAL)、4-香豆酸: CoA连接酶(4CL)、肉桂醇脱氢酶(CAD)、过氧化物酶(POD)活性测定; 其余10株用于测定抗倒伏指数和茎秆抗折力。
参照邱丽娟等[17]方法调查倒伏性, 观察并记录小区倒伏植株(主茎与地面倾斜角度小于30° )占该小区全部植株的比率。植株倒伏程度, 以小区植株无倒伏为1级; 0< 倒伏植株比率≤ 25%为2级(轻倒); 25%< 倒伏植株比率≤ 50%为3级(中倒); 50%< 倒伏植株比率≤ 75%为4级(重倒); 倒伏植株比率> 75%为5级(严重倒伏)。
参照周蓉等[18]和黄中文等[19]的方法, 抗倒伏指数=茎秆抗折力/(主茎长× 地上部鲜重)。用数字茎秆强度仪(YYD-1型, 浙江托普仪器有限公司)测定茎秆抗折力, 将第一节间的两端放于支撑架凹槽内, 两支撑点的距离为5 cm, 然后缓慢向下压, 直到茎秆折断为止, 此时读出的数值即为抗折力。参照Bhaskara- Reddy等[20]的方法测定木质素含量。参照张志良等[21]的方法测定PAL活性; 参照Knobloch等[22]的方法测定4CL活性; 参照Morrison等[23]的方法测定CAD活性; 参照熊庆娥等[24]的方法测定POD活性。
用Microsoft Excel 2003软件整理数据, SigmaPlot 12.3软件绘图, SPSS 17.0软件分析数据, Duncan氏新复极差(SSR)法进行显著性检验。对于木质素含量及PAL、4CL、CAD、POD活性文内均采用2013年试验数据。
2.1.1 倒伏情况 单作大豆苗期无倒伏; 套作大豆苗期倒伏严重, 不同大豆品种倒伏差异显著 (表1)。2012和2013年各品种倒伏程度基本一致, 南豆032-4(B1)严重倒伏, 九月黄(B2)重倒, 南豆12(B3)中倒或重倒; 综合两年数据, 各品种倒伏率表现为B1> B2> B3, B1倒伏率较B2、B3分别高出28.44%、69.02%。表明套作环境下大豆品种间植株抗倒伏能力差异较大, 耐阴性越弱的品种倒伏程度越严重。
2.1.2 茎秆抗折力 同一大豆品种, 套作下茎秆抗折力显著低于单作; 同一种植模式, 不同大豆品种无论是套作还是单作茎秆抗折力均表现出显著差异(表1)。套作模式下, B3茎秆抗折力显著高于B1, B2位于二者之间; 单作模式下, B3茎秆抗折力显著高于B1和B2, B1、B2二者间差异不显著。综合两年数据, 与单作相比, 套作模式下B1、B2、B3茎秆抗折力分别降低了71.37%、63.97%和55.10%。表明套作种植显著降低大豆茎秆抗折力, 而强耐阴性品种受影响程度相对较小, 其茎秆仍能够表现出较优的力学性能, 茎秆抗折力显著高于弱耐阴性品种。
| 表1 不同种植模式下大豆倒伏率、茎秆抗折力和抗倒伏指数 Table 1 Actual lodging percentage, snapping resistance and lodging resistance index of soybean in different planting patterns |
2.1.3 抗倒伏指数 除2013年B3品种套作与单作抗倒伏指数差异不显著外, 同一大豆品种, 套作抗倒伏指数显著低于单作; 同一种植模式, 不同大豆品种抗倒伏指数在套作和单作中表现不一致(表1)。套作模式下, B3抗倒伏指数显著高于B1和B2, B1、B2二者差异不显著; 单作模式下, 2012年B1抗倒伏指数显著低于B2和B3, B2、B3二者差异不显著, 2013年B1、B2和B3间差异均不显著, 但品种间消长规律与2012年相同。表明强耐阴性品种对套作荫蔽环境表现出较强的适应能力和调节能力, 其抗倒性较强。
同一大豆品种, 套作下茎秆木质素含量显著低于单作; 同一种植模式, 各大豆品种茎秆木质素含量在套作和单作中均表现为B3显著高于B2, B2高于B1 (图1)。方差分析表明, 种植模式(P= 0.002)和品种(P = 0.000)均极显著影响茎秆木质素含量, 且互作效应显著(P= 0.032), 耐阴性强的品种在套作环境下能保持较高水平的木质素含量, 有效提高大豆的抗倒伏能力。
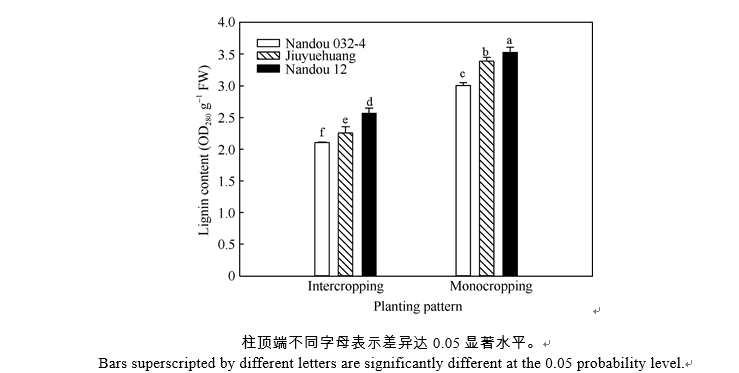 | 图1 不同种植模式下大豆茎秆木质素含量Fig. 1 Lignin content of soybean stem in different planting patterns |
2.3.1 苯丙氨酸转氨酶(PAL) 同一大豆品种, 套作茎秆PAL活性显著低于单作; 同一种植模式, 不同大豆品种茎秆中PAL活性在套作和单作中均有显著差异(图2)。套作模式中, B2、B3显著高于B1, B2、B3二者差异不显著; 单作模式中, B3显著高于B1、B2, B1、B2二者差异不显著。
2.3.2 4-香豆酸:CoA连接酶(4CL) 同一大豆品种, 套作茎秆4CL活性显著低于单作; 同一种植模式, 不同大豆品种茎秆中4CL活性在套作和单作中均有显著差异(图2)。套作模式中, B3显著高于B2, B2高于B1; 单作模式中, B2、B3显著高于B1, B2、B3二者差异不显著。
2.3.3 肉桂醇脱氢酶(CAD) 同一大豆品种, 套作茎秆CAD活性显著低于单作; 同一种植模式, 不同大豆品种茎秆中CAD活性在套作和单作中均有显著差异(图2)。套作模式中, B3显著高于B1, B2位于二者之间; 单作模式中, B3显著高于B2, B2高于B1。
2.3.4 过氧化物酶(POD) 同一大豆品种, 套作茎秆POD活性显著低于单作; 同一种植模式, 不同大豆品种茎秆中POD活性在套作和单作中均有显著差异(图2)。套作模式中, B3显著高于B1, B2位于二者之间; 单作模式中, B3显著高于B1、B2, B1、B2二者差异不显著。
 | 图2 不同种植模式下的PAL、4CL、CAD和POD活性Fig. 2 PAL, 4CL, CAD, and POD activities in different planting patterns |
相关分析表明, 套作下大豆茎秆木质素含量与倒伏率极显著负相关, 与抗折力显著正相关(表2)。说明大豆苗期茎秆木质素含量与茎秆抗倒伏能力密切相关, 木质素含量高, 大豆抗倒伏能力强。套作模式中, 茎秆木质素含量与4CL、CAD活性显著正相关, 与PAL、POD活性相关性不显著; 单作模式中, 茎秆木质素含量与PAL、4CL、CAD和POD活性均显著正相关。表明大豆茎秆中PAL、4CL、CAD和POD作为木质素合成的关键酶, 其活性影响木质素含量的高低, 而4CL、CAD活性提高是增加套作大豆茎秆木质素含量的酶学基础。
| 表2 大豆茎秆木质素含量及合成相关酶活性与抗倒伏能力相关系数 Table 2 Correlation coefficients of lignin content and enzyme activities in lignin metabolism with lodging resistance of soybean |
已有研究证明, 大豆的抗倒性与株高、茎粗、茎秆强度、地上部和地下部生物量等密切相关[9, 19, 25]。刘卫国等[2, 5]和王竹等[3]研究表明, 大豆-玉米套作模式中, 玉米遮阴导致大豆行间光强减弱、红光与远红光比值降低, 大豆发生避阴性反应, 株高增加、节间伸长、茎粗减小, 干物质积累减少, 容易倒伏。本试验进一步表明, 与单作相比, 套作大豆苗期茎秆抗折力减小, 田间倒伏严重。抗倒伏指数是衡量和评价大豆抗倒伏能力的重要参数[18, 19], 套作模式下抗倒伏指数显著低于单作, 这可能是套作大豆茎秆抗折力大幅减小而株高大幅增加造成的。总体来看, 与玉米共生期间, 光照环境的改变不仅影响套作大豆茎秆的形态建成, 还降低了茎秆强度, 使大豆抗倒伏能力降低, 倒伏的风险加大。
不同大豆品种的茎秆强度对荫蔽胁迫的响应有所差异。本试验中, 与九月黄和南豆032-4相比, 强耐阴性大豆品种南豆12受套作遮阴影响较小, 其茎秆抗折力下降幅度最小, 茎秆抗折力高, 抗倒伏指数大, 表现出较强的抗倒伏能力, 田间实际倒伏率最低。因此, 在套作大豆生产上要选择耐阴性强、茎秆综合性状突出的品种, 以实现增产增收。
木质素作为细胞壁主要成分, 具有增加细胞壁强度, 提高茎秆机械强度的功能。研究表明, 木质素含量增加可显著提高茎秆机械强度, 增强茎秆的抗压能力[26]; 茎秆木质素含量高的品种抗倒伏能力强, 不易倒伏[14, 27]。本研究表明, 套作大豆苗期茎秆中木质素含量显著低于单作, 套作大豆苗期茎秆木质素含量与抗折力极显著正相关(r = 0.890, P< 0.01), 与倒伏率极显著负相关(r = -0.889, P< 0.01)。说明茎秆中木质素含量降低是套作大豆茎秆抗折力下降的重要原因, 影响套作大豆的抗倒伏能力。本试验中, 与九月黄和南豆032-4相比, 南豆12茎秆木质素含量高, 抗折力和抗倒伏指数大, 田间实际倒伏率低。说明较高的茎秆木质素含量是其具有较强抗倒伏能力的生理基础, 使其能够在套作环境下维持较优的茎秆综合性状, 更适合于套作种植。因此, 茎秆中木质素含量高有利于增强套作大豆苗期的抗倒伏能力。
PAL、4CL、CAD和POD是木质素合成过程中的4个关键酶, 其中PAL催化L-苯丙氨酸脱氨生成反式肉桂酸, 4CL催化肉桂酸生成相应的CoA酯, CAD将肉桂醛类物质还原成相应的木质素单体; POD催化木质醇单体发生脱氢聚合反应, 参与并调节木质素在细胞壁的聚合[28]。本试验结果表明, 套作大豆茎秆木质素含量与4CL、CAD活性显著正相关, 与PAL、POD活性相关性不显著, 而单作大豆茎秆木质素含量与PAL、4CL、CAD和POD活性均显著正相关。可见, 4CL、CAD活性对套作大豆茎秆中木质素合成起重要的调节作用, 是套作大豆抗倒伏能力差异形成的酶学机制。木质素含量与POD和PAL活性之间没有表现出显著的相关性, 这可能与PAL、POD在套作环境下还参与其他抗逆性反应, 而未特异性参与木质素代谢有关[29, 30]。此外, 与单作相比, 套作大豆茎秆中4种酶的活性均有所降低, 相应地, 套作大豆茎秆中木质素含量也降低, 说明大豆茎秆木质素合成相关酶活性的变化是引起大豆单作和套作抗倒伏能力差异的重要原因。
套作环境降低大豆苗期茎秆木质素含量和相关酶活性, 抗折力和抗倒伏指数下降, 倒伏严重。强耐阴性品种具有较高的茎秆木质素含量和相关酶活性, 从而使茎秆抗折力较大, 表现出较强的抗倒伏能力, 较高的4CL、CAD活性是增强套作大豆抗倒伏能力的重要因素。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|