
{kind=link}
{kind=link}
基于 Glb 1基因序列的西南地区玉米地方品种的系统进化
[姚启伦1, 2  , 陈发波
, 陈发波1, * , 刘红芳1 , 方平1 ]
, 陈发波, 刘红芳|
|


第一作者联系方式: E-mail: yql641@aliyun.com
为探讨我国西南地区玉米地方品种 Glb 1基因序列多态性, 对40份西南地区玉米地方品种的 Glb 1基因序列测序, 并与GenBank中来自普通玉米、小颖玉米、繁茂玉米和摩擦禾的19条同源序列比对。结果表明, 来自西南地区四川、重庆、云南和贵州的玉米地方品种分别有35、25、33和25个多态性位点, 其遗传多态性分别是小颖玉米的87%、86%、85%和71%, 8个种群遗传多态性指数的大小排序为小颖玉米、四川地方品种、重庆地方品种、云南地方品种、贵州地方品种、普通玉米、磨擦禾和繁茂玉米, Tajima’D、Fu & Li’D和Fu & Li, F检验表明 Glb 1基因是中性进化基因。比较种群间DNA序列的共有多态性和固定差异, 玉米地方品种与小颖玉米、繁茂玉米、磨擦禾间存在较高的固定差异和较低的共有多态性。 Glb 1基因的系统进化关系及网状关系分析为玉蜀黍属的分类系统提供了佐证, 四川玉米地方品种有较高的遗传多态性支持我国玉米由印度经西藏传入四川的论点。
To explore the DNA polymorphism of Glb 1 genes in maize landraces from Southwest China, we cloned and sequenced the DNA sequences from 40 maize landraces, and downloaded the homologous sequences of 19 individuals representing common maize, parviglumis, Z. luxurians, and T. dactyloides from GenBank. By aligning these sequences and analyzing DNA variations in Glb 1 loci, we found 35, 25, 33, and 25 polymorphism sites in maize landraces from Sichuan, Chongqing, Yunnan, and Guizhou, respectively. Their genetic polymorphisms were 87%, 86%, 85%, and 71% of that in parviglumis. On the basis of estimates of per base pair and
per base pair, the highest level of sequence variation was found in parviglumis, followed in turn by maize landraces from Sichuan, landraces from Chongqing, landraces from Yunnan, landraces from Guizhou, common maize, T. dactyloides, and Z. luxurians. The estimates of Tajima’s and Fu & Li’s D statistic for Glb 1 gene were negative in the eight taxa, suggesting that neutrality cannot be rejected. In comparison, more fixed differences than shared polymorphisms between maize landraces and parviglumis, Z. luxurians, as well as T. dactyloides were found. The genealogical and network analysis provided the evidence for the genus Zea taxonomy. Additionally, genetic diversity of landraces was greater in Sichuan than in the other three regions. This result supports the hypothesis that the first maize migration was from India to Sichuan by way of Tibet in China.
玉米(Zea mays L.)是全球的主要粮食作物、饲料作物和经济作物, 因其独有的生物学性状和遗传学特性而成为重要的生物学研究材料[1]。西南地区是我国玉米地方品种的主要分布地带, 地方品种集中分布在四川、广西、云南、贵州和重庆的高海拔地区。从已编入国家第一、二、三集种质资源目录的4662份材料来看, 玉米地方品种资源极其丰富, 除粉质型和有稃玉米外, 其余7个类型均存在于西南地区[2]。前人[3, 4, 5, 6]对西南地区玉米地方品种进行了大量基础性研究, 在表型上地方品种有丰富的遗传变异。具体表现在株叶穗型千姿百态、籽粒颜色五彩斑斓、抗性资源丰富多彩。基于SSR分子标记技术, 田孟良等[7]、吴渝生等[8]、刘永建等[9]、Yao等[10]先后在DNA分子水平对西南地区玉米地方品种进行了遗传多样性研究。研究结果表明, 西南地区玉米地方品种在分子水平上有丰富的遗传变异和较高的育种利用潜势。
近年来, 利用DNA序列变异研究生物群体演化已成为遗传学研究的一大热点。Glb 1是单拷贝核基因, 位于第1染色体的长臂, 编码存在于玉米成熟胚中的贮藏蛋白— — 球蛋白[11]。Belanger和Kriz[12]研究发现, 玉米Glb 1基因包含5个外显子和4个内含子, 基因编码区长度为2400 bp, Glb 1-S及因小片段核苷酸插入所衍生的Glb 1-I和Glb 1-O是Glb 1基因的3种突变体。由于Glb 1基因内插入突变所产生的突变体在近缘物种间存在较大变异, 从而可用于物种的分类及种群演化研究。Hilton和Gaut[13]首次用Glb 1和Adh 1基因分析栽培玉米的起源与进化; 基于Waxy、Glb 1、c 1和Adh 14个DNA序列差异, Tiffin和Gaut[14]分析了多年生大刍草和二倍体多年生大刍草的种群演化关系; 田孟良等[15]分析了西南糯玉米地方品种Waxy基因序列多态性。这些研究为基于Glb 1基因序列变异进行种群系统进化关系研究提供了范例。
西南地区玉米地方品种作为特殊的玉米育种资源, 其传入年代和时间尚无定论, 在较小时间与空间尺度上主效基因位点的变异形式及种群演化过程均未见相关报道。为此, 本研究通过对西南地区玉米地方品种Glb 1基因部分DNA测序, 分析Glb 1基因的遗传多态性, 旨在探究玉蜀黍属的系统进化关系, 为我国玉米的引入途径提供理论依据。
基于西南地区玉米地方品种的地域分布和种质特征, 结合田间鉴定地方品种农艺、经济性状及SSR标记的遗传多样性分析, 从四川、重庆、云南和贵州选取典型特色、代表性好的地方品种40份, 由长江师范学院玉米育种课题组提供, 材料编号及地理来源见表1。
在培养箱中发芽, 取黄化幼苗叶片约3 g在液氮中研磨, 采用CTAB法[16]]抽提并纯化基因组DNA, 利用琼脂糖凝胶电泳检测DNA浓度和质量。
PCR扩增片段为Glb 1基因启动子(200 bp)及其后的约1000 bp序列, 扩增的引物序列为F: 5′ -CCG GAT AAG CAC GGT AAG GA-3′ ; M: 5′ -CTT GCT GAA GCT CGA CAG GA-3′ 。引物由上海生工生物工程有限公司合成, 扩增产物约1200 bp。PCR体系50 µ L含2× Taq Master Mix 25 µ L, 上、下游引物(10 nmol mL-1)各0.5 µ L, DNA 2 µ L (含50 ng DNA), 无RNase水 22 µ L。在Eppendorf AG PCR仪上进行反应, 反应程序为94℃预变性5 min, 1个循环; 94℃变性1 min, 53℃退火1 min, 72℃延伸2 min, 共35个循环; 最后于72℃延伸10 min。用上海生工生物工程有限公司SanPrep柱式DNA胶回收试剂盒回收PCR扩增产物, 将扩增产物连接到质粒载体PMD19-T (TaKaRa)后, 再转染感受态大肠杆菌菌株DH5a。在含有氨苄青霉素培养基上培养感受态菌株, 挑取阳性克隆, DNA测序由上海生工生物工程有限公司完成。
将测序得到的各DNA序列提交GenBank核酸序列数据库, 获取GenBank登录号(表1), 并在GenBank中下载部分近缘物种的同源序列: 普通玉米(AF064212、DQ356513、DQ356524、EF635959、EU643507)、小颖玉米(AF064233、EF539575、EF536901、EF539611、EF539645)、繁茂玉米(AF064228~ AF064232)和磨擦禾(AF064233~AF064235、AF377694)。根据植物学分类和地理来源将序列划分为四川玉米地方品种、重庆玉米地方品种、云南玉米地方品种、贵州玉米地方品种、栽培玉米、小颖玉米、繁茂玉米和磨擦禾共8个种群。通过对Glb 1基因序列的多态性分析及其统计学检验研究西南地区玉米地方品种的种群演化。
采用DNAMAN 6软件进行序列比对分析, 用DAMBE软件进行序列格式转换, 用MEAGA 6软件计算突变位点数、多态位点数、简约信息位点数和单显突变数, 用DnaSP[17]软件计算Glb 1基因同义替代位点数、非同义替代位点数、碱基替代位点数、遗传多态性指数 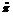

| 表1 西南地区玉米地方品种群体来源及Glb 1基因序列登录号 Table 1 Geographical origin and Glb 1 sequence accession number of maize landraces from the southwest of China |
采用最大似然法(Maximum Likelihood, ML)分析系统发育, 采用MEAGA 6软件分析ML, 贝叶斯推断采用MrBayes 3.2.3软件进行模型和参数估计[20], 利用Jmodeltest 2进行模型检验, 基于hLRT标准选择的最适碱基替代模型, 外显子 + 内含子序列数据最适碱基替代模型是GTR+G, 外显子序列数据最适碱基替代模型是T92+G, 内含子序列数据最适碱基替代模型是JC+G。ML分析采用最大似然法进行系统树构建(Test Maximum Likehood Tree), 并采用自展分析(Bootstraps, BS)进行检验, 自展分析重复250次。
采用系统网状重建法(The phylogenetic network reconstruction method)分析种群单体型的系谱关系。首先利用HyPhy软件中的GARD方法分别检测Glb 1基因外显子、内含子及外显子 + 内含子的重组信息, 结果表明Glb 1基因外显子为非重组DNA序列(lg Likelihood = -2715.35, AIC=5016.54), 因此, 利用外显子序列数据资料构建网络支系图[21, 22]。采用Network 5.4.4.0软件(Fluxus Technology Ltd., Clare, Suffoik, UK)中的中介连接网络法(the median-joining network method)进行网状支系分析。
根据序列比对结果, 供试DNA序列平均长度为1132 bp, 变幅953~1157 bp。在DNA序列中检测到810个突变位点, 其中多态性位点625个, 简约信息位点232个, 单显突变(singleton) 114个。各取样序列外显子、内含子长度变异和多态性位点见表2。在8个种群中, 小颖玉米的突变位点数最多, 其外显子中有24个核苷酸同义替换、15个非同义替换; 四川地方品种、重庆地方品种、云南地方品种和贵州地方品种的突变位点数分别为35、25、33和25个, 以四川地方品种最高; 供试8个种群外显子平均长度为757 bp, 变幅751~762 bp, 表明Glb 1基因编码区长度较为保守。
比较各种群的遗传多态性指数 
| 表2 种群内Glb 1基因多态位点 Table 2 Polymorphic sites of Glb 1 genes in the eight taxa |
| 表3 种群内Glb 1基因序列多态性和中性进化检验 Table 3 Sequence polymorphism and neutral evolution test of Glb 1 genes in the eight taxa |
由种群内基因平衡进化检验结果(表3)可知, 各种群Tajima’ D、Fu and Li’ D和Fu and Li’ F均不显著, 且为负值, 表明Glb 1基因在各种群中是中性进化基因位点, 不存在选择作用。
共有多态性(shared polymorphisms)是未被遗传漂变消除的多态性, 与群间分歧年代负相关, 而与奠基群体大小正相关, 固定差异(fixed differences)代表种群在独立演化过程中不同的遗传漂变[23, 24]。通过比较种群间DNA序列的共有多态性和固定差异, 可分析其遗传相关性。由表4可知, 4个玉米地方品种群体DNA序列间具有较高的共有多态性, 而不存在固定差异, 其中四川地方品种与重庆地方品种、云南地方品种和贵州地方品种间的共有多态数分别达56、54和49, 固定差异数均为0。相反, 4个玉米地方品种群体与小颖玉米、繁茂玉米、磨擦禾3个近缘种群间存在较高的固定差异和较低的共有多态性, 与磨擦禾间的共有多态数均为0。就小颖玉米而言, 其DNA序列与4个玉米地方品种群体DNA序列间有较高的共有多态性, 共有多态数变幅为20~28; 与普通玉米有1个固定差异, 21个共有多态数。这在Glb 1序列上反映了西南地区玉米地方品种与近缘种群间的遗传关系, 玉米地方品种与普通玉米和小颖玉米有相近的遗传关系, 与繁茂玉米和磨擦禾有较远的遗传关系。
| 表4 种群间的共有多态数和固定差异数 Table 4 Number of shared polymorphisms and fixed differences between taxa |
基于8个种群Glb 1基因序列的“ 外显子+内含子” 区, 采用最大似然法(ML)构建系统发育树(图1)的似然值为-3310.072, 4种核苷酸的理论频率为A: 0.1804、G: 0.3507、T: 0.1571、C: 0.3118, 不变位点的比率为0.4838, γ 形值(gamma shape parameter)为0.6888。系统发育关系树显示出玉蜀黍和磨擦禾两大分支, 玉蜀黍分支分为A、B两个亚支。A亚支包括支系A1、A2、A3、A4和A5, 支系A1由7个云南地方品种、7个重庆地方品种、1个四川地方品种和5个普通玉米品种构成(BP = 72), 支系A2由分别来自重庆、四川和云南的1个地方品种构成(BP = 73), 支系A3包括1个小颖玉米、8个四川地方品种、1个重庆地方品种和1个云南地方品种(BP = 100), 来自贵州的10个地方品种、1个重庆地方品种和1个云南地方品种构成支系A4 (BP = 58), 1个小颖玉米单独构成支系A5 (BP = 89); B亚支包括支系B1和系B2, 支系B1仅由1个小颖玉米构成(BP = 97), 支系B2包括3个小颖玉米和5个繁茂玉米(BP = 100)。由系统发育树(图1)可见, 相同地理来源的玉米地方品种有相近的遗传关系, 多数同一地理来源的地方品种聚居在同一支系。
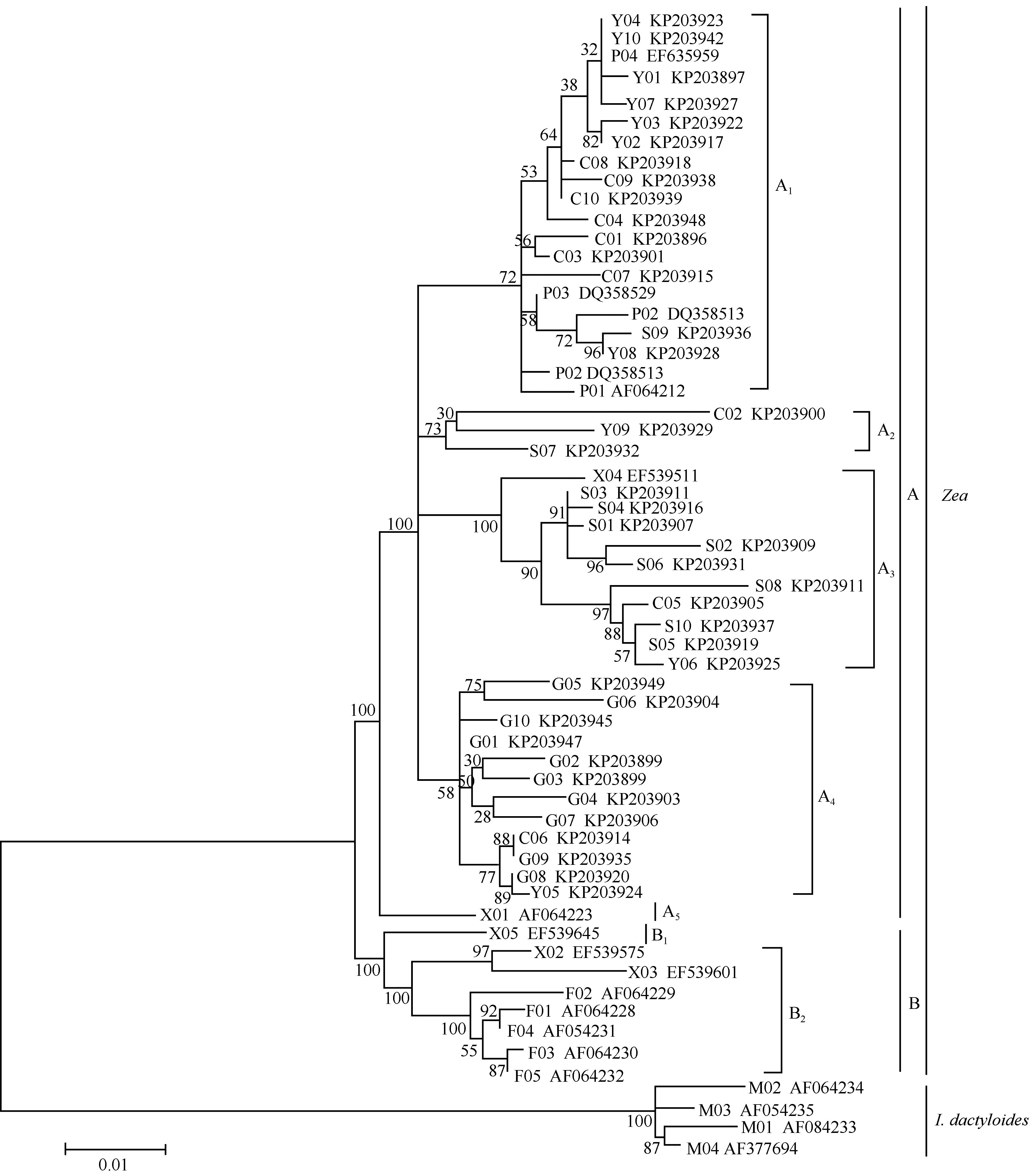 | 图1 8个种群Glb 1基因系统发育树Fig. 1 Phylogenetic tree ofGlb 1 genes in the eight taxa |
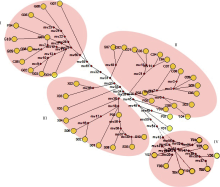
为进一步明确种群间系统演化关系, 利用近缘种小颖玉米、普通玉米和玉米地方品种Glb 1基因外显子区的序列变异进行网状进化关系分析, 网状支系图(图2)显示了来自小颖玉米、普通玉米和4个地方品种供试种群50个单体型的系谱关系。由图2可见, 供试种群的50个单体型被划分成四大支系。支系I由10个贵州地方品种单体型、1个重庆地方品种单体型和1个普通玉米单体型构成, 支系II由9个重庆地方品种单体型、4个四川地方品种单体型、2个云南地方品种单体型和3个普通玉米单体型构成, 6个四川地方品种和1个普通玉米单体型构成支系III, 8个云南地方品种单体型构成支系IV。其中, 5个小颖玉米均单独成支且位于中间位置, 3个普通玉米位于支点中央三, 玉米地方品种成群分居, 这与Glb 1基因序列的系统进化关系分析一致。
 | 图2 玉米地方品种、普通玉米和小颖玉米Glb 1基因外显子单体型的网状进化关系Fig. 2 Network relationships of exon haplotypes ofGlb 1 genes in maize landraces, common maize, and parviglumis |
基因序列多态性是生物种群演化的基础, 而序列多态性和基因进化是物种在同一演化过程中的两个不同层次[25]。分析单拷贝核基因序列的遗传多态性可了解基因在不同种群中的演化进程。Buckler等[26]通过分析现代玉米与其祖先种在多个基因位点上的序列变异, 发现现代玉米在驯化过程中保留了祖先种70%的遗传多样性。Chen和Li[27]的研究表明, 任两个玉米品种在DNA沉默位点的差异达1.4%, 是其他禾本科作物的14倍, 相当于灵长类黑猩猩与人的差异。本研究在Glb 1基因序列中检测到810个突变位点、625个多态性位点、232个简约信息位点, 遗传多态性指数 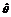
玉米起源于美洲, 在墨西哥进化后, 向北美和南美传播。玉米作为重要的粮食和饲料作物, 在美洲驯化历史达5000年之久, 并在全球范围内进行了长达3000多年的广泛栽培。目前还未发现具有现代栽培玉米植株形态的的野生玉米, 玉米首次传入我国准确的年代和途径尚无定论。据古籍记载, 1511年前玉米引入我国, 可能有两条引入途径, 一条是由印度经西藏传入四川, 另一条是由海路传入东南沿海地区, 再扩散到内地[1]。玉米传入我国后, 经近500年的风土驯化, 形成了极为丰富的地方品种类型, 并在其进化过程中产生了由新的隐性基因控制的多样化区域, 即次生基因中心[28]。
由Glb 1基因序列的系统进化关系发现, 地方品种和普通玉米在同一亚支, 表明地方品种与现代玉米相近的起源与演化, 同时支持Iltis和Doebley[29]对于玉蜀黍属的分类系统。基于SSR标记和B染色体多态性分析, 笔者[10, 30]发现四川玉米地方品种有较高的遗传多样性, 为玉米由印度经西藏传入四川提供了证据。本研究结果表明, 西南地区玉米地方品种有较高的遗传多态性, 其中四川玉米地方品种有最高的遗传多态性指数, 高于其余3个西南地区地方品种群体。就单体型的系谱关系而论, 四川玉米地方品种10个单体型分居两大支系, 且与普通玉米单体型相邻, 反映了单体型间较大的遗传差异。这为我国玉米由印度经西藏传入四川提供了佐证, 同时表明西南地区玉米地方品种最早在四川种植, 逐步向毗邻地区扩散。
Glb 1是中性进化基因, 玉米地方品种在Glb 1基因位点有较高的遗传多态性, 玉米近缘种属小颖玉米、繁茂玉米、磨擦禾与地方品种间存在较高的固定差异和较低的共有多态性。本研究结果为玉蜀黍属分类系统及我国玉米的引入途径提供了佐证。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|