


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
水稻卷叶突变体 rl28的特性与基因定位
[冯萍* , 邢亚迪* , 刘松, 郭爽, 朱美丹, 娄启金, 桑贤春, 何光华, 王楠*  ]
]
]
|
|


叶片是光合作用的主要器官, 适度卷曲有利于改善群体光照, 提高光能利用率, 因此, 发掘和研究叶片发育相关基因是改良株型和植物生长发育研究的重要基础工作。本研究报道了一个新的水稻稳定遗传卷叶突变体 rolled leaf 28 ( rl28), 与野生型相比, rl28从拔节期起叶片开始沿中轴脉向内侧卷曲, 叶片的卷曲度均极显著高于野生型, 且叶夹角也不同程度小于野生型。扫描电镜及石蜡切片观察表明, rl28叶片单位面积气孔数、气孔导度显著高于野生型, 蒸腾速率极显著高于野生型, rl28中脉增大及临近的2个泡状细胞数量减少。遗传分析表明该突变性状受1对隐性核基因控制, RL28基因被定位在第5染色体标记5-43和5-34之间, 物理距离为90 kb。本研究将为 RL28基因的图位克隆及功能研究奠定基础。
Leaves play a very important role in plant development for their function of photosynthesis. Moderate rolling leaves can facilitate the improvement of plant’s population structure and enhance light-use efficiency, which is very important in ideotype breeding. Therefore, the rolled leaf genes which regulate morphology in rice are important for exploring plant type and improving basic research in molecular biology. This study reported a new gene rolled leaf 28 ( rl28), which was derived from EMS-treated restorer line Jinhui10. The mutational trait inherited steadily after several generations’ self-crossing. Compared with the wild-type, the leaves of rl28 began to curl along the vasculan bundle in medial axis from jointing stage, leaf rolling index was significantly higher than that of the wild-type, and leaf angles were less than those of wild-type. Scanning electron microscopy and morphological analysis showed stoma number per 10-5 m2 and stomatal conductance were significantly higher than those of the wild-type, transpiration rate was significantly higher than that of wild-type. Compared with the wild-type, midrib of rl28 was much larger, and the number of the two adjacent vesicular cells decreased. Genetic analysis showed that the mutational trait was controlled by a single recessive nuclear gene. RL28 was finally mapped on chromosome 5 between SSR markers 5-43 and 5-34 with an interval of 90 kb. These results provide a foundation for cloning and function analysis of RL28.
水稻叶片是进行光合作用和呼吸作用的主要器官, 是产量形成的决定性因子, 也是株型构成的重要因素[1]。卷叶是水稻叶片形态特征之一, 适度卷曲不仅能够有效解决叶长和叶挺两者之间的矛盾[2], 而且能防止叶片披垂、改善植株通风透光性、增强下表面的光合作用, 故有利于水稻产量的提高[3]。
目前对水稻卷叶机理的研究, 主要是通过卷叶突变体来挖掘相关重要基因并研究其功能, 从分子水平解析卷叶调控的遗传机制[4]。叶片的发育是一个复杂而多级的过程, 叶片发生卷曲主要与叶片的近轴/远轴面的极性发育异常有关[5], 据此将水稻卷叶突变体分为两类, 即内卷和外卷。叶片内卷表现近轴化发育, 如SHALLOT-LIKE1(SLL1)编码与拟南芥KANADIs基因家族同源的MYB转录因子, 其功能缺失会导致离轴叶肉细胞的程序化死亡, 抑制离轴特征的发育, 从而引起sll1突变体叶片的高度内卷[6]; 与拟南芥ZIP/Ago7同源的OsAGO7基因过量表达导致叶片上表面泡状细胞数目减少和泡状细胞变小, 叶片内卷[7]; Rolling leaf14 (RL14)编码2OG-Fe(II)氧化酶, 上表面泡状细胞因气孔复合体变小, 蒸腾速率下降而体积明显变小[8]。叶片外卷表现为远轴化发育, ADAXIALIZED LEAF1 (ADL1)基因编码一种植物特异的类Calpain半胱氨酸蛋白酶, 参与叶轴形成、顶端分生组织 (SAM)的维持和叶原基位置特异性以及胚乳发育等, ADL1的突变使得叶极性和SAM叶原基的位置遭到破坏, 导致远轴面的上表皮及叶肉组织近轴化, 分化为类泡状结构, 进而导致叶片外卷[9]; Zou等[10]通过T-DNA插入的方法获得一个外卷叶突变体oul1, 其叶片外卷是敲除水稻Rice outermost cell-specific gene5(ROC5)基因造成的; 锌指类结构同源转录因子OsZHD1过量表达诱导叶片背面卷曲和叶片披垂[11]。此外, 研究发现叶片卷曲还与生长素有关, 在拟南芥和矮牵牛 (Petunia hybrida)中发现的基因YUCCA和FLOOZY的突变, 会引起内源吲哚乙酸含量的变化, 从而使叶片卷曲[12, 13]; 在水稻中发现了YUCCA同源基因NARROW LEAF7(NAL7), 编码一种黄素单加氧酶, 该基因突变引起IAA含量变化, 导致叶片卷曲[14]。
本研究利用甲基磺酸乙酯(ethylmethane sulfonate, EMS)诱变籼稻恢复系缙恢10号, 获得了一个可以稳定遗传的卷叶突变体rl28。在遗传分析和表型鉴定的基础上进行了分子定位, 最终将其定位在第5染色体上。这将为该基因的进一步图位克隆和功能分析奠定基础。
突变体rl28来自EMS诱变恢复系缙恢10号, 经过多代自交, 突变性状稳定遗传。2012年, 用表型正常的不育系西农1A与突变体rl28杂交, 同年在海南种植F1, 并收获F2种子, 于2013年在西南大学水稻研究所分别种植亲本和F2群体。
在植株成熟后, 分别选择10株缙恢10号和10株rl28突变体, 考察株高、有效穗、每穗粒数、每穗实粒数、千粒重、结实率等主要农艺性状, 同时测量叶片的长和宽。
在拔节期分别取野生型缙恢10号和突变体的新鲜叶片, 分别用FAA固定液(70%乙醇90 mL, 40%甲醛5 mL, 冰醋酸5 mL)固定, 经脱水、透明、石蜡包埋。取10 μ m的横切片, 用番红、固绿染色, 在蔡司荧光显微镜下观察、照相。
在拔节期分别取野生型缙恢10号和突变体的新鲜叶片, 利用导电胶将样品固定, 在日立SU3500扫描电子显微镜下观察并照相。
在拔节期随机选取野生型和rl28各10株, 测定其功能叶长度并于叶片长度1/2处测量叶宽。同时计算功能叶的叶片卷曲指数LRI (leaf rolling index)[15], 取平均值。LRI = (Lw-Ln)/Lw, 其中Lw为叶片展开时最宽处的宽度(cm), Ln为叶片最宽处卷曲后两叶缘之间的距离(cm)。
在拔节期随机选取野生型和rl28各10株, 测量功能叶的叶夹角, 即茎秆和叶脉之间的夹角。
采用BSA法定位目标基因[16]。按CTAB法提取亲本和基因池DNA[17], 按碱煮法提取F2群体DNA[18]。参照http://www.gramene.org/microsat/的SSR引物序列, 根据西农1A和缙恢10号的序列差异比对设计InDel引物, 均由上海英骏生物技术有限公司合成。PCR总体系为12.6 μ L, 含1.25 μ L 10× PCR buffer、1 μ L 50 ng μ L-1 DNA模板、0.75 μ L 25 mmol L-1 MgCl2、0.5 μ L 2.5 mmol L-1 dNTPs、8.0 μ L ddH2O、1.0 μ L 10 μ mol L-1引物、0.1 μ L 5 U μ L-1TaqDNA聚合酶。PCR程序为94℃预变性5 min; 94℃变性30 s, 55℃退火30 s, 72℃复性30 s, 35个循环; 72℃延伸10 min。PCR产物经10%非变性聚丙烯酰胺凝胶电泳, 快速银染后观察[19]。
F2定位群体中, 将具有西农1A带型的单株记为A, 具有rl28突变亲本带型的单株记为B, 具有杂合带型的单株记为H。根据公式[(H+2A)/2n]× 100计算遗传距离并构建连锁图谱, 其中H表示定位群体中出现杂合体带型单株的数量, A表示出现西农1A正常带型的单株数, n表示用于定位的隐性群体总株数。
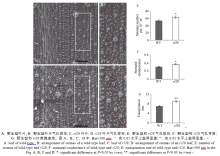
与野生型相比, 突变体rl28在苗期并没有明显差异, 而在拔节期时叶片开始沿中轴脉纵向内卷曲, 该性状一直持续到植株成熟(图1-A, B)。野生型的卷曲度较低, 倒一叶至倒三叶均在10%以下, 而突变体则很高, 超过了70%, 表现十分明显的卷曲特性, 均极显著高于野生型(图1-C)。突变体倒三片叶的叶夹角均小于野生型, 分别降低了3.1%、5.2%和4.5%, 其差异达到显著或极显著水平(图1-D)。
 | 图1 野生型与rl28的表型分析Fig. 1 Phenotype of wild-type (WT) and rl28 mutant |
为了进一步探究突变体叶片卷曲的原因, 我们对野生型和突变体的叶片进行了石蜡切片观察, 结果表明, 与野生型相比, rl28中脉的面积显著增大(图2-A~E), 中脉与第一个维管束(图2-M, N)、第一与第二个维管束(图2-K, L)之间的泡状细胞数量明显减少, 而其他维管束之间则没有变化(图2-I, J)。DROOPING LEAF(DL)是水稻调控中脉发育的关键基因, rl28的中脉面积显著增大, 因此利用QPCR检测了DL在野生型和rl28中的表达情况, 结果表明DL在野生型和rl28叶片间无显著差异(图2-F)。对野生型和突变体的叶片进行了扫描电镜观察、光合数据分析及单位面积气孔数量统计表明, rl28单位面积气孔数量(图3-A~E)和气孔导度显著高于野生型(图3-F)、蒸腾速率极显著高于野生型(图3-G)。
 | 图2 野生型与突变体rl28拔节期组织学观察Fig. 2 Histological observation of leaves of wild-type (WT) andrl28 mutant in jointing stage |
 | 图3 野生型与突变体rl28拔节期扫描电镜观察Fig. 3 Scanning electron microscope observation of leaves of wild-type (WT) andrl28 mutant in jointing stage |
rl28突变体的株高、千粒重、穗长、有效穗数、剑叶长宽、倒二叶长宽、倒三叶长宽与野生型相比无明显差异, 而突变体的结实率只有40.2%, 相比野生型极显著的下降(图4)。
 | 图4 野生型(WT)与突变体rl28的农艺性状分析Fig. 4 Analysis of agronomic traits in the wild-type (WT) and rl28 mutant |
用表型正常的不育系西农1A与rl28杂交, F1表型正常, 说明该突变体受隐性基因控制。F2代群体中出现明显的分离, 分别表现双亲性状, 其中正常单株2280株, 突变单株752株。经卡方测验, 正常株∶ 突变株符合3∶ 1分离比(χ 2=0.05< χ 20.05=3.84), 表明rl28突变体受隐性单基因控制。
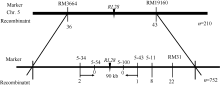
选用平均分布的400对引物对初步定位群体的亲本和基因池进行连锁性分析, 发现位于第5染色体的引物RM3664和RM19160与突变位点连锁。通过F2代突变单株分析, 将rl28突变位点定位在RM3664和RM19160之间, 遗传距离分别为8.57 cM和10.24 cM。为了进一步缩小定位区间, 在RM3664和RM19160之间筛选出6对在亲本间呈现出多态性的引物, 分别是RM31、5-11、5-34、5-43、5-54和5-100 (表2), 通过F2代突变单株分析, 最终将rl28突变位点定位在引物5-34和5-43之间, 分别只有2个和1个交换株, 且没有包含关系, 5-54和5-100与RL28共分离, 没有交换株, 物理距离为90 kb (图5)。通过信息学比对发现该区间共有12个开放阅读框, 其中3个编码表达蛋白, 2个编码还原转座子, 2个编码成束蛋白, 1个编码谷氧还蛋白, 1个编码植物生长响应因子15, 1个编码剪接因子U2AF, 1个编码锌指蛋白、1个编码Ras相关蛋白(表1)。
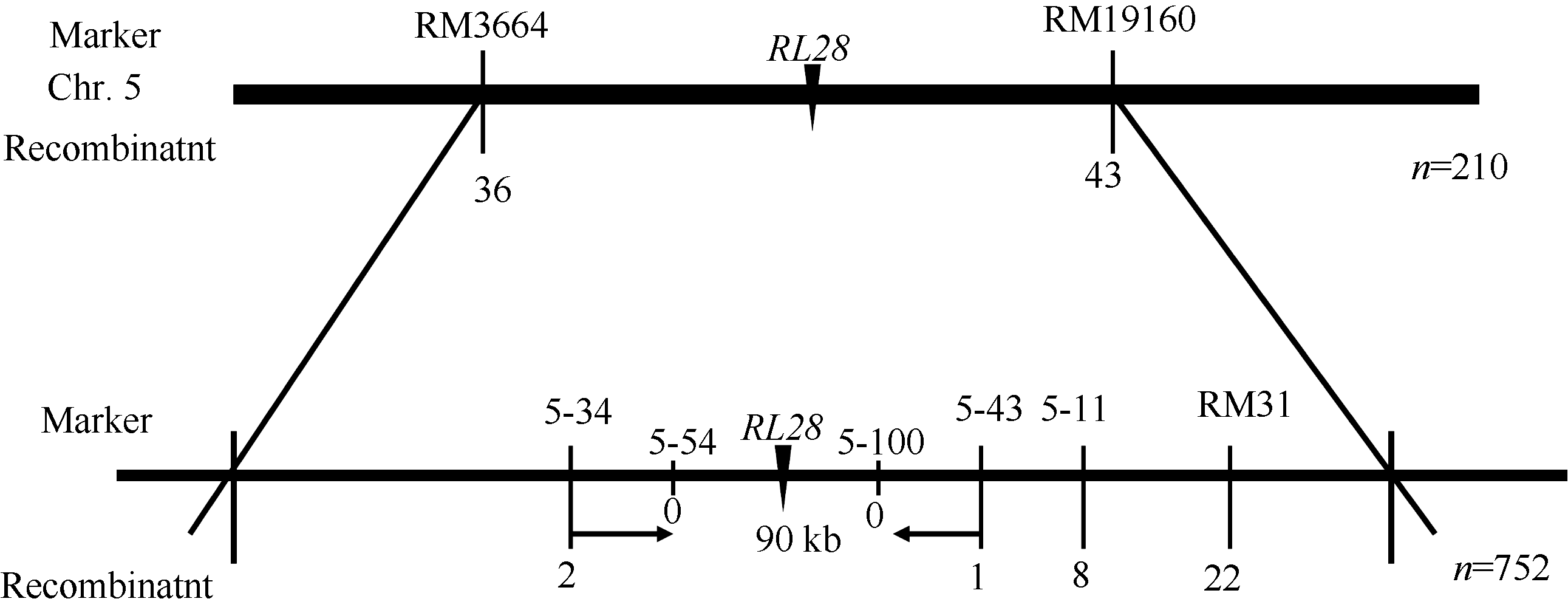 | 图5 RL28基因在第5染色体上连锁图谱Fig. 5 Linkage map of RL28 on chromosome 5 of rice |
| 表1 定位区域内预测基因 Table 1 Predicted genes in mapping region |
| 表2 第5染色体上的连锁标记 Table 2 Polymorphic markers used in the fine mapping on chromosome 5 of rice |
叶片是水稻最主要的同化器官, 是塑造理想株型的重要性状之一, 卷叶是异型叶中的一种, 其最直接的效应是使植株保持挺立, 改善上层叶片的透光率, 提高群体光合效率。随着分子生物学的不断
发展, 利用突变体研究水稻叶片的发育和形态成为新的研究热点[5]。
通过对拟南芥、玉米和水稻等植物突变体的研究表明, 卷叶突变的形成除了与植物叶片腹-背极性发育及环境有关, 如sll1、adl1等[6, 9], 水稻叶片还可以通过控制泡状细胞数量的多少或体积的大小来控制叶片的卷曲, 比如Roc5、SEMI-ROLLED LEAF1(SRL1)、BR-deficient dwarf1(BRD1)等[10, 20, 21], Roc5编码与GLABRA2 (拟南芥亮氨酸拉链同源域基因第Ⅳ 类) 同源的蛋白, Roc5对泡状细胞的发育起负调控作用, 它结合在表皮因子基因PFL1的启动子区域, 特异性地调控PFL1, 当Roc5被抑制时, PFL1表达减少, 泡状细胞数目和大小增加, 叶片外卷; 而当它被过表达时, PFL1表达增加, 泡状细胞大小和数目减少, 出现内卷。半卷叶基因SRL1也与泡状细胞有关, srl1突变体表型为泡状细胞数目增加, SRL1编码一种糖基磷脂酰肌醇蛋白, 特异性调控编码液泡H+-ATPase亚基和H+-焦磷酸酶基因表达, 抑制液泡的形成, 减少泡状细胞数量; 突变体则增加泡状细胞数量, 引起半卷叶。此外, BRD1基因在控制水稻泡状细胞数量方面有一定作用, brd1突变体叶片的泡状细胞数量比野生型多, 但该突变体叶片并没有发生反卷, 因为该基因除了影响泡状细胞的数量外, 还影响叶片的其他细胞结构, 因此该突变体植株表现为矮化并且叶片严重扭曲。
rl28临近中脉的2个泡状细胞数量减少, 中脉面积显著增大, 这是造成其叶片内卷突变表型的关键原因之一。目前在水稻中, 并没有报道同时影响中脉和泡状细胞发育及气孔发育而导致叶片卷曲的突变体。水稻中脉的发育主要受YABBY基因DL的负向调控[22, 23], 该基因的严重突变导致叶片中脉发育不全, 叶片披垂, 同时花器官也发生变化[22, 23]。DL基因存在复杂的调控模式, 其启动子及第一、二个内含子都是表达调控的重要区域, 部分dl等位突变体在启动子上游区域的突变仅仅影响中脉的形成, 而不影响花器官的发育[24]。rl28的中脉面积增大, 但DL基因在rl28突变体与野生型之间无差异, 中脉增大可能与DL没有关系。另外rl28的单位面积气孔数显著高于野生型, 目前植物中关于叶片气孔数与叶片卷曲度关系的研究并不多。张静懿[25]在拟南芥中发现生长素参与负调控气孔发育, 并揭示了这一过程受核受体介导的生长素信号转导途径介导, 最终通过生长素响应因子ARF5/MP (MONOPTEROS) 直接负调控气孔发育的上游正调控因子编码基因STOMAGEN的表达, 从而负调控气孔发育, 突变体的气孔显著增多。Price等[26]认为在叶片水分不足时, 泡状细胞膨压降低, 叶片卷曲同时气孔关闭, 蒸腾速率降低, 当水分充足时泡状细胞膨压升高, 叶片恢复平展。rl14-1因水分不足导致泡状细胞异常, 气孔复合体面积变小[8]。rl28的单位面积气孔数增多, 气孔导度增大, 蒸腾速率升高, 这可能是导致叶片卷曲的原因之一。
RL28基因定位在第5染色体标记5-43和5-34之间90 kb的区间内。在已定位和克隆的卷叶突变体中, 位于第5染色体上的仅有rl7被定位在RG13和RG57之间[27]、rl8被定位在RM6954和RM6841之间[28], 这两个突变体与rl28位于不同的定位区间, 因此rl28是一个未报道的水稻内卷突变体。在RL28基因的定位区间内的12个开放阅读框中含有1个植物生长素响应因子ARF15, 已有研究发现, 植物生长素响应因子(ARF)在植物根、茎、叶、花、果等中许多生长发育过程, 不管是对单子叶植物还是双子叶植物都起着重要的作用, 且不同的ARF具有各自独特的功能[29]。另一个是C3HC4型锌指蛋白, 锌指蛋白是真核生物基因组中最丰富的一类转录因子, 主要通过与核酸的相互作用来显示不同的功能, 如促进转录、抑制转录、单链DNA结合、IKNA结合或RNA/DNA双向结合[30, 31]。究竟是ARF15还是C3HC4、或者其他基因突变引起叶片卷曲, 需要进一步研究。
通过EMS诱变籼稻缙恢10号获得卷叶突变体rl28, 从拔节期功能叶片开始明显卷曲, 叶夹角显著低于野生型。rl28中脉的面积比野生型显著变大, 中脉与第一个维管束、第一与第二个维管束之间的泡状细胞数量明显减少。rl28的气孔导度和单位面积气孔数量显著高于野生型, 蒸腾速率极显著高于野生型。该突变性状受一对隐性核基因控制, 突变位点被定位在第5染色体SSR标记5-43和5-34之间, 物理距离约90 kb。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|