


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
河南小麦新育成品种(系)白粉病抗性鉴定与分子标记检测
[曹廷杰1, 2 , 陈永兴1 , 李丹1 , 张艳1 , 王西成2 , 赵虹2 , 刘志勇1, *  ]
]
]
|
|


利用华北地区流行的白粉菌菌株E09和E20, 分别对河南省小麦新品种(系)区域和预备试验参试材料908份(2009—2013年度)和412份(2009—2012年度)进行苗期白粉病抗性鉴定, 同时利用与 Pm2、 Pm4a、 Pm8和 Pm21基因连锁的分子标记检测相关抗病基因的分布。结果显示, 抗E09的材料占21.9% (199/908), 抗E20的材料占9.5% (39/412), 同时抗E09和E20的材料仅占3.6% (15/412)。在908份供试材料中, 580份含有1BL/1RS, 占63.9%, 含 Pm8或新的1RS来源抗白粉病基因; 另有2份材料含6AL/6VS来源广谱抗白粉病基因 Pm21, 8份可能携带 Pm2, 2份可能含有 Pm4a; 有6份材料可能含有多个抗白粉病基因。表明河南省近年育成的小麦新品种(系)依然含有对我国白粉菌菌系有效的抗白粉病基因, 但抗源遗传基础较窄, 部分已经或正在丧失抗性, 应加快引进和利用新的多样化抗病基因资源。
The powdery mildew resistance of 809 (2009-2013) and 412 (2009-2012) new bred wheat varieties (lines) from Henan provincial regional trials was tested using Blumeria graminis f. sp. tritici( Bgt) isolates E09 and E20, respectively. Molecular markers linked to powdery mildew resistance genes Pm2, Pm4a, Pm8, and Pm21were used to detect the presence of the individual resistance gene. Among the 908 varieties (lines) inoculated by E09, 199 (21.9%) exhibited resistance. Among the 412 varieties (lines) inoculated by E20, 39 (9.5%) exhibited resistance. Only 15 (3.6%) varieties (lines) showed resistance to both E09 and E20. The 1RS chromatin was detected in 580 out of 908 (63.9%) varieties (lines), indicating the common use of Pm8 or newly developed 1BL/1RS translocation lines in wheat breeding program. Two varieties carried the board spectrum powdery mildew resistance gene Pm21 originating from 6AL/6VS translocation. Eight and two lines might contain the Pm2and Pm4aloci, respectively. Six varieties (lines) seemed to carry at least two powdery mildew resistance genes. Our results indicated that the newly developed wheat varieties (lines) in Henan province are important powdery mildew resistance resources. However, the common use of 1BL/1RS translocations and a few powdery mildew resistance genes has revealed a very narrow genetic diversity of the resistant resource. It is very urgent to introduce new diversified and broad spectrum powdery mildew resistance genes into the commercial wheat breeding program.
小麦在中国是仅次于水稻和玉米的第三大粮食作物。河南省位于我国冬小麦主产地黄淮平原区, 是我国小麦的最适生态区, 小麦播种面积和总产量均居全国首位, 河南省小麦生产对全国粮食生产和国家粮食安全起着重要作用。
由Blumeria graminisf. sp. tritici引起的小麦白粉病是一种世界性的小麦真菌病害, 造成的产量损失可达5%~34% [1]。1927年, 首先在我国江苏省发现小麦白粉病, 其后逐渐扩展到西南各省和部分沿海地区。20世纪70年代以后, 由于耕作制度的改革和生产条件的改善, 尤其是种植密度的提高和水肥使用量的增加, 致使小麦白粉病不断扩展蔓延北移, 而且发病程度不断加重, 河南省也成为小麦白粉病的重发区[2]。特别是近年来, 由于矮秆品种、半矮秆品种的大面积种植以及主栽品种的抗源单一化和白粉病菌系的变化等原因, 小麦白粉病危害日趋严重, 目前已成为河南省小麦生产上急待解决的重大灾害性问题之一。喷施农药虽然可以起到一定的防治效果, 但增加成本且污染生态环境, 而选育和利用抗病品种是防治小麦白粉病最经济、有效和环境安全的方法。然而, 小麦白粉病菌具有高度的变异性, 品种抗性容易被新的毒性突变体克服而导致抗性丧失和病害流行[3]。因此, 不断发掘新的抗病基因并应用于抗病品种培育, 才可以持续有效地控制小麦白粉病的危害。
自1969年Sears和Briggle [4]将抗白粉病基因Pm1定位在7AL染色体上以来, 迄今已在小麦基因组49个位点(Pm1~Pm53)发掘出正式命名的60多个主效抗白粉病基因[5, 6], 但这些抗病基因主要来源于小麦的近缘种属, 绝大多数抗白粉病基因由于与不良性状紧密连锁或者抗性已经丧失, 未能在育种中直接利用。从抗病基因利用角度来看, 来自小麦品种中的抗病基因是育种家的首选, 因为其具有优良的遗传背景, 更容易直接在育种和生产上得到应用。因此, 发掘和鉴定普通小麦品种中的抗白粉病基因, 对抗病育种具有重要意义。
分子标记已经广泛应用于抗病基因的检测。利用RFLP技术, Pm1首次被定位在Xwhs178附近[7], 此后陆续报道了30多个抗白粉病基因的共分离或紧密连锁分子标记。Morris等[8]研究表明位于5D染色体上的标记Xcfd81与Pm2连锁, 遗传距离为2.0 cM; Hartl等[7]将Pm2定位在标记位点Xwhs295附近(2.7± 2.6 cM)。Ma等[9]通过建立Pm2和Pm4的RFLP标记发现, Pm2与Xbcd1871相距3.5 cM, Pm4a两侧分别与Xbcd1231-2A(1)和Xbcd292-2A连锁, 遗传距离均为3.5 cM; 之后又发现位于2A染色体的标记位点Xgwm356与Pm4a相距2.0 cM[12]。Qi等[10]报道标记OPH171900与Pm21共分离; Liu等[11]进一步将OPH171900转化为可靠的SCAR1400和SCAR1265。这些标记的开发为检测育种材料中的对应抗白粉病基因提供了极大的便利。
本研究通过对河南省2009— 2013年度育成的908个小麦新品种(系)进行白粉菌苗期接种鉴定, 了解其抗性表现; 同时利用与已知抗白粉病基因连锁的分子标记检测抗白粉病基因, 并对这些品种(系)的抗白粉病基因遗传基础进行分析, 为河南省小麦抗白粉病新品种的选育提供材料和技术支撑。
908份材料是来自2009— 2013年度参加河南省区域试验和预备试验的小麦新品种(系), 分别由参试单位提供, 感病对照品种为薛早。用于接种鉴定的病原菌为华北地区流行的小麦白粉菌菌系E09和E20, 由中国农业科学院植物保护研究所周益林研究员惠赠。本试验用E09鉴定了全部908份材料, 用E20鉴定了2009— 2012年度的412份材料。
在中国农业大学小麦遗传育种温室进行人工接种鉴定, 首先用感病品种薛早大量繁殖白粉菌, 用其成熟孢子作为接种体。将小麦种子穴播于塑料培养盘中, 每穴一个品种(系), 约20粒, 每盘种植一穴薛早作为感病对照。在温室中培养至一叶一心, 将已充分发病的薛早繁菌盆置于待鉴定幼苗培养盘的四周, 定期用掸子扫动。待幼苗长到二叶一心(接种15 d左右), 感病对照充分发病时第一次记载抗病性, 3 d后复查一次。参照吴全安[13]的方法, 将寄主侵染型(IT)分为6种类型, 分别是免疫(IT=0)、过敏性坏死(IT=0; )、高抗(IT=1)、中抗(IT=2)、中感(IT=3)和高感(IT=4)。
二叶一心时, 取健康叶片按CTAB法[14]提取基因组DNA。用于检测1BL/1RS易位、Pm2、Pm4a和Pm21基因的分子标记信息见表1。在Applied Biosystems GeneAmp PCR System 9700上进行扩增反应。反应体系为10 µ L, 含有l µ L 10× buffer, l µ L MgCl2 (15 mmol L-1), 0.2 µ L dNTP Mixture (10 mmol L-1), l µ L引物(2 µ mol L-1), 0.1 µ L Taq DNA聚合酶(5 U µ L-1), 2 µ L基因组DNA (20 ng µ L-1), 4.7 µ L去离子水。扩增程序为94℃ 5 min; 94℃ 45 s, 退火45 s, 72℃ 1.5 min, 35个循环; 最后72℃延伸10 min。PCR产物在4℃保存。Xcfd81、Xgwm356和Xcau127扩增产物经8%非变性聚丙烯酰胺凝胶电泳检测。AF1/AF4扩增产物通过1%琼脂糖凝胶电泳检测。
| 表1 用于抗白粉病基因检测的分子标记 Table 1 Molecular markers used for detecting powdery mildew resistance genes |
在E09鉴定的908份材料中, 有199份表现抗病(21.9%), 其中112份免疫, 56份高抗, 31份中抗; 另有682份材料表现感病, 包括129份中感和553份高感; 还有27份材料表现抗感分离(表2)。这表明河南省近年育成的小麦新品种对E09抗性水平整体偏低, 并呈逐年降低的趋势, 以2011— 2012年度品种抗性比率最低, 2012— 2013年度稍有提高(表2)。弱春性小麦新品种对E09的抗性频率均高于半冬性品种(表2)。
| 表2 河南小麦新品种(系)对白粉病菌系E09和E20的苗期抗性反应 Table 2 Seedling reactions of Henan wheat varieties (lines) to Bgt isolates E09 and E20 |
E20接种鉴定结果表明, 39份材料表现抗病(9.5%), 其中6份免疫, 9份过敏性坏死, 14份高抗, 10份中抗。在2009— 2010年度鉴定的38个弱春性材料中, 仅有1份对E20菌株表现抗病; 在2011— 2012年度鉴定的33个半冬性小麦新品种(系)中, 也只有1份表现抗病(表2), 说明河南省现阶段育成的小麦新品种(系)抗白粉病菌系E20的非常少。2009— 2010、2010— 2011和2011— 2012年度抗E20的品种频率分别为6.4%、11.0%和5.8%。
共鉴定出15份材料(国麦301、锦麦8号、郑农01059、郑麦00314、温麦988、兰诱1号、偃亳197、濮科麦9号、大路8号、春丰0021、优抗6号、智超2号、泰麦621、乐麦598和郑麦108)同时对E09和E20表现抗性, 抗病频率仅3.6%。
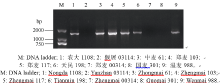
2.2.1 1BL/1RS的分子标记检测及抗性分析 利用黑麦染色质特异引物AF1/AF4, 从908份小麦材料中发现580份(63.9%)能扩增出1.4 kb的特异DNA片段(图1), 表明在新育成的小麦品种(系)中1BL/1RS易位系仍然在大量应用, 且以2009— 2010年度的频率(74.4%)最高, 2012— 2013年度的频率(53.2%)最低(图2)。含有1BL/1RS易位的材料中, 77.1% (447/580)对E09表现感病, 推测这447个品种(系)携带的1BL/1RS是来源于洛夫林等品种的Pm8基因; 另有133份材料对E09菌系表现抗病或抗感分离, 包括51份对E20菌系也表现抗病或抗感分离的材料, 推测这些品种(系)可能含有不同于Pm8的新1BL/1RS易位或其他抗白粉病基因。
 | 图1 利用AF1/AF4引物组合检测1BL/1RS易位结果Fig. 1 1BL/1RS translocation detected by primer pairs AF1/AF4 |
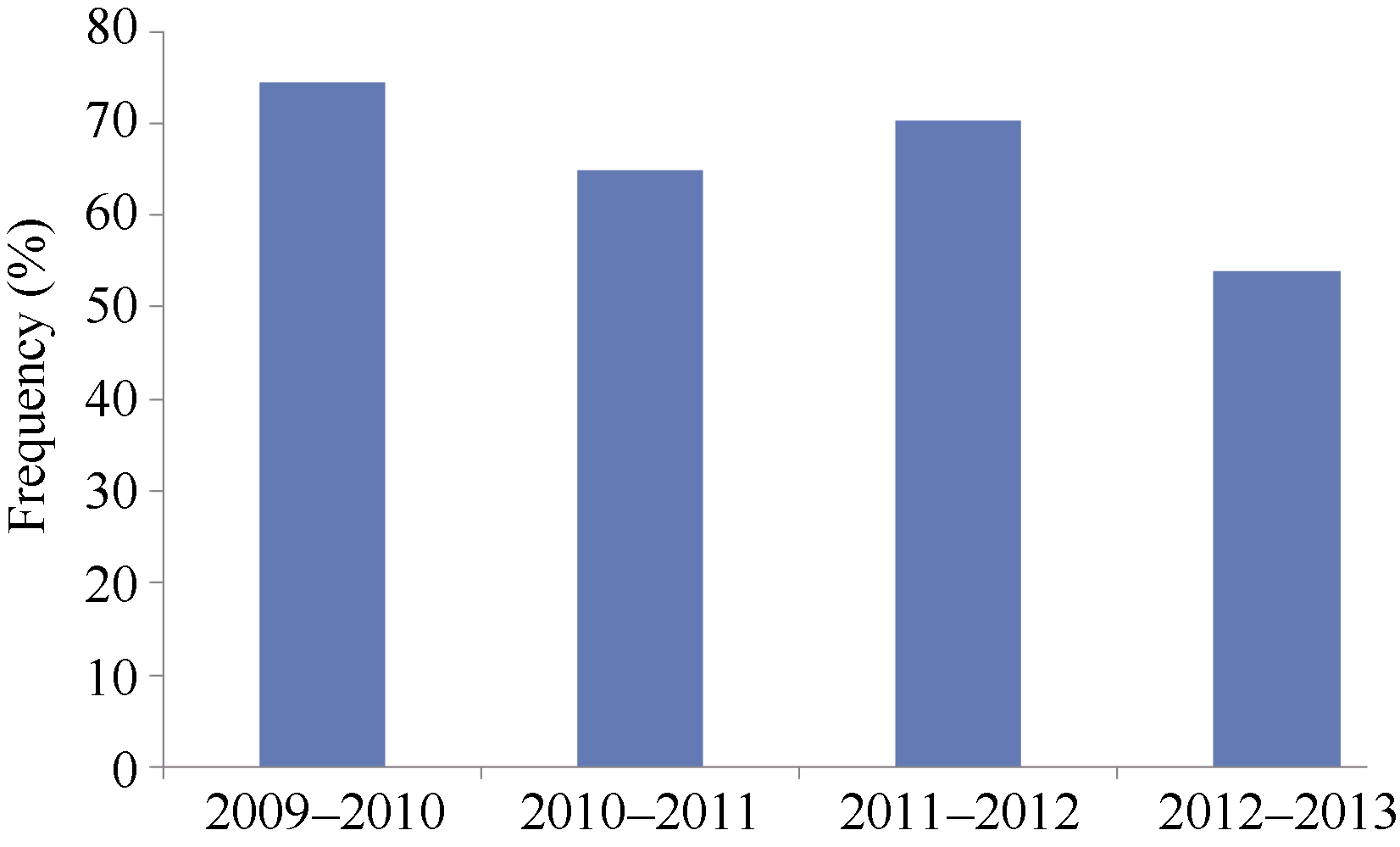 | 图2 不同年份小麦新品系1BL/1RS易位出现频率Fig. 2 Frequency of 1BL/1RS translocation in wheat lines in different years |
2.2.2 抗白粉病基因Pm2分子标记检测及抗性分析
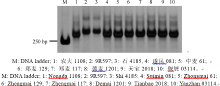
利用与抗白粉病基因Pm2紧密连锁的分子标记Xcfd81对908份材料进行了检测, 在其中8份(遂民081、农大1108、郑麦117、中麦61、德麦1201、郑麦129、偃展03114和天宝2018)中扩增出Pm2连锁的特异条带(图3), 表明这些品种(系)可能含有Pm2基因。接种鉴定结果证实它们对E09均表现抗性, 但都不抗E20。
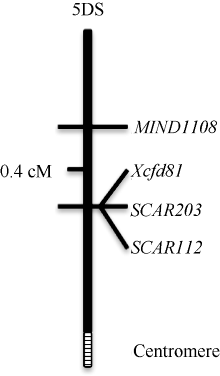
为明确这些品种(系)中是否确实含有Pm2基因, 以本实验室育成的新品种农大1108进行了抗病性遗传分析和分子标记验证。利用E09接种, 农大1108与感白粉病品系9R597的F2代群体分离出105个抗病单株和48个感病单株, 符合显性单基因3∶ 1的遗传模式(χ 2=2.98, P> 0.05); 对其中部分F3代家系进行抗性鉴定, 有32个家系纯合抗病, 60个家系抗感分离, 28个家系纯合感病, 符合显性单基因1∶ 2∶ 1的遗传模式(χ 2=0.68, P> 0.05)。通过分子标记筛选, 找到与农大1108中抗白粉病基因MlND1108连锁的SSR标记Xcfd81及SCAR标记SCAR203和SCAR112, 最终将目标抗病基因定位在5DS染色体臂上(图4), 位于Pm2所在遗传区间[18]。农大1108系谱中有来源于英国的小麦品种Riband, 含有位于5DS染色体上的Pm2基因[5], 因此推测农大1108中的抗白粉病基因很可能是Pm2。
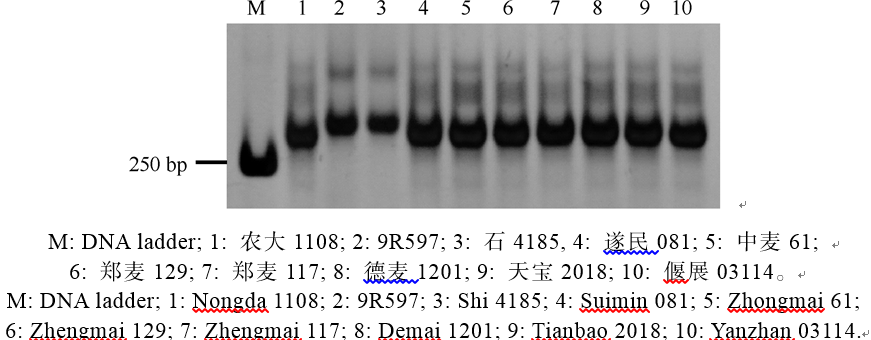 | 图3 利用SSR标记Xcfd81检测抗白粉病基因Pm2Fig. 3 Powdery mildew resistance gene Pm2 detected by SSR marker Xcfd81 |
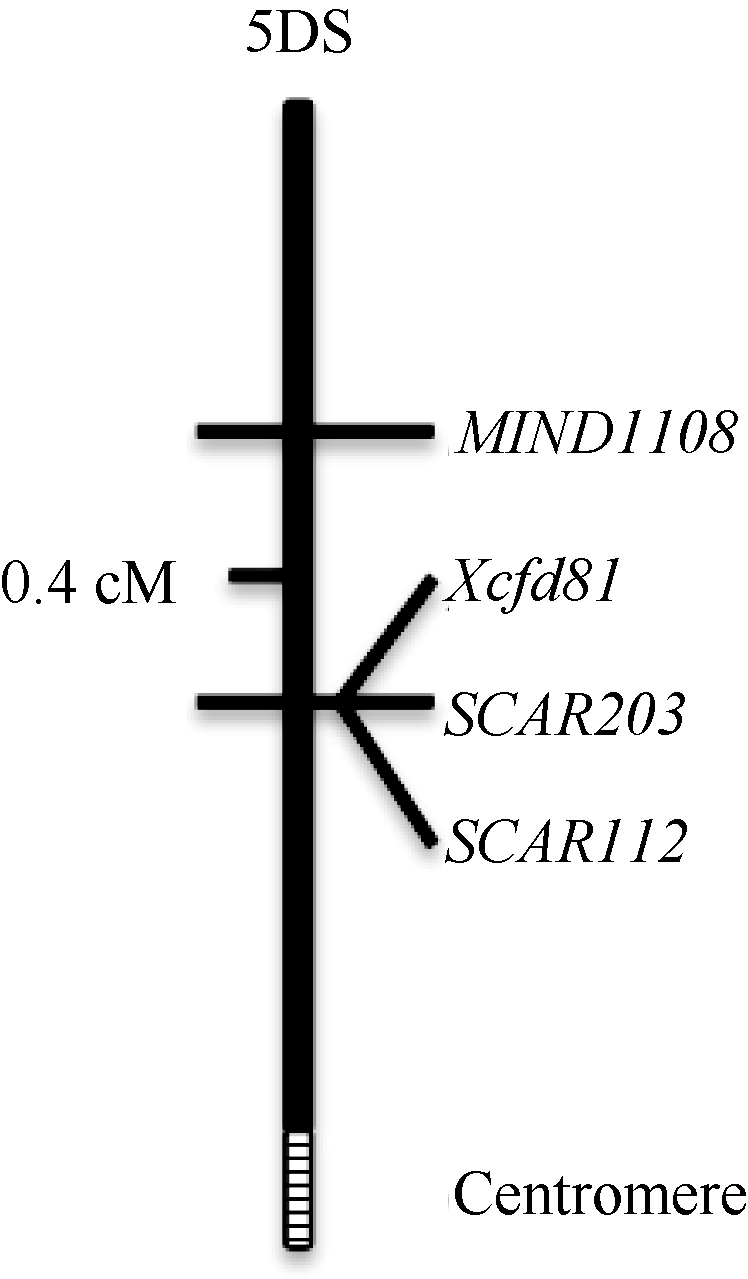 | 图4 小麦品种农大1108抗白粉病基因分子标记图谱Fig. 4 Genetic linkage map of powdery mildew resistance gene in wheat variety Nongda 1108 |
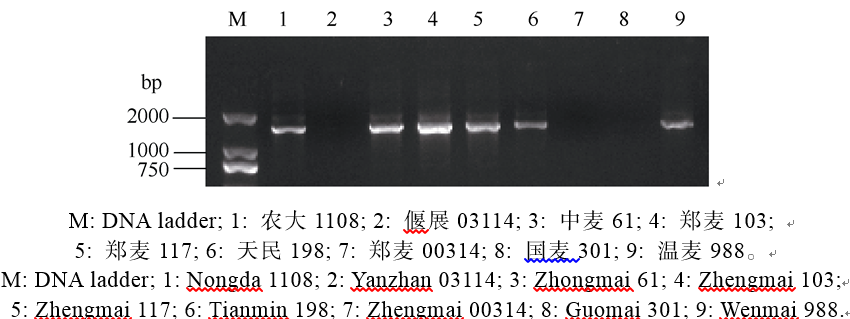
2.2.3 抗白粉病基因Pm4a分子标记检测及抗性分析 利用与Pm4a紧密连锁的分子标记Xgwm356进行PCR扩增分析, 以Khapli (Pm4a)为阳性对照, 在908份材料中发现只有粮丰998和郑麦103扩增出与Pm4a连锁的特异条带(图5)。分子标记分析结果表明粮丰998和郑麦103可能含有Pm4a, 但这2个品系对白粉菌E09和E20均表现感病。
 | 图5 利用SSR标记Xgwm356检测抗白粉病基因Pm4aFig. 5 Powdery mildew resistance gene Pm4a detected by SSR marker Xgwm356 |
2.2.4 抗白粉病基因Pm21分子标记检测及抗性分析 利用与Pm21共分离的分子标记Xcau127对供试的908个材料进行检测, 以92R149 (Pm21)为阳性对照。在国麦301和石郑816中扩增出Pm21的特异标记条带(图6), 推测这2个品种携带Pm21基因。接种鉴定结果表明, 国麦301对E09和E20都表现抗病, 而石郑816对E09和E20表现抗感分离。
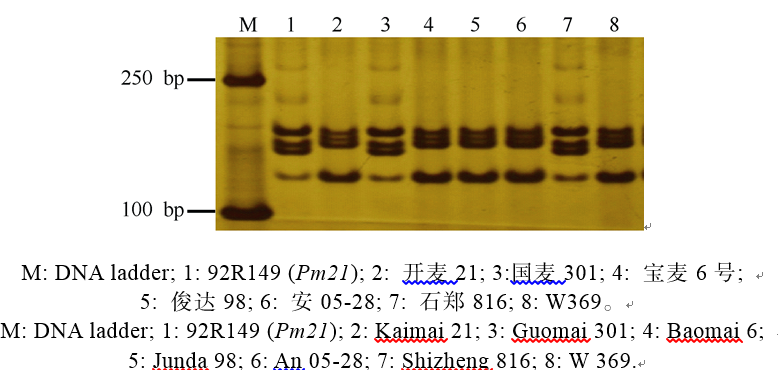 | 图6 利用SSR标记Xcau127检测抗白粉病基因Pm21Fig. 6 Powdery mildew resistance gene Pm21 detected by SSR marker Xcau127 |
2.2.5 聚合多个抗白粉病基因的小麦品种(系)
供试材料中部分品种(系)含有多个抗白粉病基因。遂民081、农大1108、中麦61、郑麦117除了可能含有Pm2以外, 还含有1BL/1RS, 这些品种(系)对E09均具有抗性, 但抗病性可能来源于Pm2, 1BL/1RS上的抗白粉病基因Pm8已经丧失抗性; 郑麦103除含有1BL/1RS外还可能含有Pm4a, 但对白粉菌E09和E20均表现感病; 石郑816可能含有1BL/1RS和Pm21, 对E09和E20均表现抗感分离, 表明在Pm21位点仍然存在分离(表3)。
| 表3 部分可能含有多个抗白粉病基因的小麦品系 Table 3 Partial wheat varieties containing several powdery mildew resistance genes |
近年来, 我国小麦白粉病的发生呈加重趋势。据全国农业技术推广服务中心植物病虫情报统计(http://www.natesc.gov.cn/), 2013和2014连续两年全国小麦白粉病发生面积均在666.7万公顷以上, 其中河南沿黄稻茬麦区和中北部高产区、江苏沿海和沿江麦区偏重发生, 造成不同程度的经济损失。对我国不同麦区不同年代小麦品种及种质材料的抗白粉病鉴定结果表明, 我国近年育成的品种及保存的种质材料抗病性现状不容乐观, 抗性较好的品种和种质材料数量较少, 特别是黄淮麦区的抗病品种的频率更低, 应引起高度重视[19, 20, 21, 22, 23]。本研究利用小麦白粉病菌系E09和E20对河南省2009— 2013年度区试小麦新品种(系)进行了苗期抗病性鉴定, 发现这些新育成品种(系)对华北地区流行白粉病菌系E09的抗性频率在20%左右; 而抗毒性较强的E20的频率更低, 仅有9.5%, 在2009— 2010年度38个弱春性和2011— 2012年度33个半冬性品种(系)中, 分别仅有1个抗E20。这说明近年河南省新育成小麦品种(系)对白粉病菌流行菌系的抗病频率呈下降趋势, 鉴于这些鉴定材料中只有一部分能在未来几年审定后推广, 所以预期生产上使用的抗白粉病品种的比例会较低。因此, 引入多样化的抗白粉病基因, 加强种质创新, 是小麦高产多抗育种的迫切需求, 亟待加强。尽管如此, 目前仍然有一些抗性较好的新品种(系)可供育种选择(表4), 这些材料都具有较好的农艺性状, 可直接用于小麦育种。
| 表4 部分对E09和E20具有较好抗性的小麦新品系抗病性鉴定和分子标记检测结果 Table 4 Molecular marker detections of powdery mildew resistance genes in new bred wheat lines and their reaction to Bgt isolates E09 and E20 |
小麦1BL/1RS易位系因携带抗白粉病基因Pm8、抗条锈病基因Yr9、抗叶锈病基因Lr26和抗秆锈病基因Sr31, 为世界小麦育种广泛应用[5, 24, 25, 26], 我国早期引入的1BL/IRS代表品种有洛夫林10号和高加索等[27]。尽管这些抗病基因已经丧失抗病性, 但大量含有1BL/1RS易位的品种目前仍然在生产和育种中应用[28]。如, 2004年报道1BL/1RS易位系在我国北部冬麦区、黄淮麦区、长江中下游麦区和西南麦区的分布频率分别为48.0%~54.0%、40.0%~ 50.4%、20.0%~6.9%和21.0%~34.6% [29]; 2011年报道Pm8基因在参加国家区域试验的品种中占有相当高的比例, 尤其是在黄淮南部和北部地区培育的品种中[19]。2006— 2010年国家审定的75个小麦品种中1BL/1RS易位利用率仍然较高, 以河北省和河南省育成的品种比率最高[30]。本研究利用黑麦染色质特异引物AF1/AF4检测河南省908个小麦新品种(系), 结果1BL/1RS易位系占参试品种的63.9%, 比率高于以前的报道。值得注意的是, 1BL/1RS易位系的比率已经从2009— 2010年度的74.4%降到2012— 2013年的53.2%, 说明育种工作者已经开始有意识地减少利用1BL/1RS资源。本研究还发现, 这些1BL/1RS易位系有较大一部分可能携带洛夫林10号等来源的Pm8基因, 但部分品种(系)可能携带不同于原来Pm8基因的新1BL/1RS易位。
小麦抗白粉病基因Pm2来源于粗山羊草, 定位于5DS染色体上, 在我国部分地区和欧州中部对小麦白粉病菌优势菌系表现出良好的抗性[31, 32], 但该基因在我国部分麦区已经开始失去抗性或者抗性降低[33, 34]。本研究利用与Pm2基因连锁的分子标记Xcfd81对供试材料进行了分子检测和人工接种苗期抗病性鉴定, 结果发现有8个小麦新品种(系)可能含有Pm2基因, 均对白粉菌系E09表现抗病或免疫; 对农大1108的抗性遗传分析和分子标记定位证实其可能含有Pm2基因; 良星66可能含有Pm2基因或其等位基因, 该品种对17个不同来源的白粉菌菌株均表现出较好的抗性[35]。这些事实表明Pm2或其等位基因在我国部分麦区依然抗性较好且比较稳定, 在小麦抗白粉病育种仍有价值, 应合理利用, 发挥其抗病优势。
王俊美等[36]对已知小麦抗白粉病基因的抗性研究发现, 在河南省已经出现了Pm4a基因的毒性菌系, 而高安礼等[33]报道, 虽然Pm4a已经失去抗性, 但基因聚合体Pm4a+Pm2却表现出较好的抗性。此外, 孟雅宁等[37]通过检测河北省保存的农艺性状较好的65份小麦遗传资源的抗白粉病基因, 发现只有河农7069可能携带抗白粉病基因Pm4a, 对白粉病具有较好的抗性。本研究利用与Pm4a紧密连锁的分子标记Xgwm356, 在908个小麦品种(系)中只检测到2个品系可能含有Pm4a, 均对白粉菌系E09和E20表现高感。由于不能单纯从分子标记检测确认其是否确实含有Pm4a基因, 还需要做进一步的系谱追踪、遗传分析和分子标记定位。
来源于小麦-簇毛麦6AL/6VS染色体易位的抗白粉病基因Pm21是目前抗性最突出的抗白粉病基因之一, 表现抗性强、抗谱广[38, 39], 且已经在育种中得到应用, 尤其在西南麦区小麦品种中的利用频率较高[40]。在黄淮麦区Pm21基因的频率还很低, 仅有个别品种携带Pm21, 如河北省的石麦15 [19], 而在山东省227个育成品种和442个地方品种中未发现含有Pm21[41]。本研究利用与Pm21共分离的分子标记Xcau127检测908个小麦品种(系), 只在国麦301和石郑816中扩增出目标带, 其中国麦301对白粉菌系E09和E20均表现免疫, 而石郑816表现为抗感分离。由于Pm21基因白粉病抗性突出, 而这些含有Pm21的新品种已经在黄淮麦区表现出良好的农艺性状和产量特征, 预计很快就会成为重要的小麦育种亲本, 育出衍生新品系。但值得注意的是, 目前已经在西北春麦区的甘肃省[42]、长江中下游冬麦区的湖北省[43]、长江中上游冬麦区的四川省[44]、东北春麦区的黑龙江省和辽宁省[45], 以及黄淮冬麦区的山西中南部[46]、河南省[47]、河北省[48]、陕西省[34]等地区发现对Pm21具有毒性的菌株, 在亲本选配和育种中应加以注意, 以免重蹈Pm8基因的覆辙。
大面积种植含单个抗性基因的品种会对病原菌群体形成强大的选择压力, 加速新的毒性小种的产生和在群体中的频率增加, 从而克服抗病基因的抗性, 导致病害大流行。将多个主效抗病基因聚合到一个品种中, 将有助于拓宽品种的广谱抗性, 提高抗病品种的稳定性和持久性, 延长在生产上的使用寿命。研究表明, 聚合2个及以上抗病基因的品种(育种后代材料)与单个抗病基因相比抗病性均得到极大的改善[33, 49, 50, 51]。在本研究中, 河南省小麦新品种(系)中存在抗病基因聚合现象, 如聚合1BL/1RS和Pm2的品种(系)农大1108、中麦61和郑麦117, 均对白粉菌系E09具有较好的抗病性。由于1BL/1RS易位携带的Pm8基因已经丧失抗性, 推断这些品种的抗性源自Pm2基因, 所以需要尽快引入新的抗白粉病基因。
分子标记是鉴定一个品种是否含有特定基因的高效手段之一, 且具有操作简单、低成本等优点。目前, 利用图位克隆技术仅获得了Pm3基因的功能标记, 可用于分子标记辅助选择, 然而大部分已知的Pm3位点等位基因都对我国白粉病菌菌系不具有抗性, 没有育种利用价值。与抗白粉病基因连锁的分子标记往往由于遗传背景的差异, 在不同的品种中还不能完全确认是否一定携带某个目的基因, 需要通过系谱追溯、遗传分析和连锁分析等予以佐证。除Pm8和Pm21等少数位于外源染色体上的抗病基因可以用染色质特异的分子标记准确追踪外, 其他抗白粉病基因还需要借助分子标记、遗传分析和系谱分析加以推断。
在检测的河南省小麦新品种(系)中, 还有一部分抗性较好的品种未能用上述分子标记检测出含有Pm2、Pm4a、Pm8和Pm21基因, 推测其可能含有其他抗白粉病基因或由于遗传背景差异所致。高通量的分子标记技术为快速明确其含有的抗白粉病基因提供了新的途径, 有待进一步检测分析。
河南省近年来育成的小麦新品种(系)抗病频率偏低, 且呈现出逐年降低的趋势, 抗病基因遗传基础狭窄, 应在育种实践中尽快引进多样化的抗病基因资源, 聚合有效的抗病基因并加快种质创新进程, 避免抗病基因的单一化, 培育具有广谱和持久抗性的新品种。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|