{kind=link}
{kind=link}
普通小麦DH155抗白粉病基因的分子作图及应用分子标记辅助选择将其转移
[管昌英* , 郭军* , 薛凤博, 张广旭, 王宏伟, 李安飞, 孔令让*  ]
]
]
|
|


普通小麦品系DH155对白粉病菌表现高抗。为明确DH155所携带抗白粉病基因的遗传方式及与抗病基因连锁SSR标记, 利用DH155与高感小麦品系SN2890杂交获得的F2和F2:3群体进行接种鉴定和遗传分析, 发现DH155对白粉菌菌株E09的抗性受1对显性基因控制, 暂命名为 MlDH155。BSA和分子标记分析结果显示, MlDH155与SSR标记 Xcfd81和 Xcfd18连锁。利用已发表的中国春和粗山羊草D基因组序列开发新标记, 进一步将 MlDH155定位于标记 XsdauK525和 XsdauK527之间, 其遗传距离分别为0.2 cM和0.8 cM。将DH155与感白粉病优良品系HB133-4和旱10杂交, 在F2~F4代, 结合优良农艺性状选择、分子标记辅助选择和抗白粉病鉴定, 获得3个高抗白粉病且农艺性状优异的株系(SDAU2100、SDAU2101和SDAU2102)。利用14个白粉菌菌株对DH155进行苗期接种鉴定表明, DH155对13个菌株表现抗病反应型。这些菌株对DH155的毒力谱与已知抗白粉病基因 Pm2相似, 但DH155对Bg78-3和Bg44-5菌株的反应型与携带 Pm2的Ulka/8*Cc不同。结合本试验结果和 Pm2基因的相关报道, 推测 MlDH155可能是 Pm2或其等位基因。
Hexaploid wheat ( Triticum aestivum L.) line DH155 is highly resistant to wheat powdery mildew caused by Blumeria graminisf. sp. tritici( Bgt). To identify the Bgtresistance gene(s) in DH155, we developed an F2 population and its derived F2:3families by crossing the resistant line DH155 with the susceptible line SN2890. The segregation ratios indicated that the seedling resistance to BgtE09 in DH155 was controlled by a single dominant gene, which was tentatively designated MlDH155. By bulked segregation analysis, two codominant SSR markers, Xcfd81 and Xcfd18, were identified to be linked to MlDH155. To identify the closely linked markers to the targeted gene, we developed five new molecular markers based on the published D genome sequences of Chinese Spring and Aegilops tauschii, which permitted mapping of MlDH155 within an interval of 1.0 cM, flanked by XsdauK525 and XsdauK527. The Pm resistant line DH155 was crossed with two elite wheat lines (HB133-4 and Han 10) but susceptible to powdery mildew. Subsequently, two powdery mildew resistant lines with the genetic background of HB133-4 and one resistant line with Han 10 background were developed by genotypic and phenotypic selection, which were designated by the name of SDAU2100, SDAU2101 and SDAU2102, respectively. Among the 14 Bgtisolates tested at the seedling stage, DH155 was resistant to 13 and susceptible to 1 isolates. The virulence pattern of these Bgtisolates on DH155 was similar to that of the known powdery mildew resistance gene Pm2, but the reactions of DH155 to two Bgtisolates differed from those of Ulka/8*Cc carrying Pm2. Compared to previous studies about Pm2, MlDH155was most likely to be either the same as or an allele of Pm2.
由禾本科布氏白粉菌小麦专化型[Blumeria graminis (DC.) E. O. Speer f. sp. tritici, Bgt]引起的小麦白粉病, 是世界小麦产区的主要病害之一[1]。目前, 白粉病是我国北部冬麦区和黄淮海冬麦区最重要的小麦病害, 在西南和长江中下游麦区, 是仅次于条锈病或赤霉病的第二大病害[2]。小麦白粉病大流行, 不仅造成严重的产量损失, 而且还会严重降低小麦的加工品质和烘烤品质[3, 4]。实践证明, 利用优良抗源培育抗病新品种是防治小麦白粉病最安全、经济和有效的方法[5, 6]。因此, 研究、挖掘和利用抗白粉病基因资源, 以拓宽抗性抗源多样化, 具有非常重要的理论和生产意义。迄今, 已在小麦基因组46个位点上发现70多个抗白粉病基因或等位基因, 然而随着白粉病流行菌种的不断变化和病菌毒力的提高, 很多抗病基因的抗性逐渐丧失[7, 8]。例如, 来源于黑麦1RS的小麦抗白粉病基因Pm8曾经在20世纪70年代至80年代中期广泛用于育种, 但到80年代中期以后对该基因的毒力菌株频率迅速上升, 使其抗性在全国各地迅速丧失[9]。另外, 一些品种或品系携带某些抗性基因但农艺性状较差, 不适合直接做育种亲本, 如Pm12、Pm16和Pm20[10]。因此, 继续发掘优异新抗源, 培育抗白粉病新品种具有十分重要的意义。
DH155是国家小麦改良中心泰安分中心保存的小麦遗传材料, 系谱为济麦19/鲁麦21 [11]。本实验室经过多年的种植鉴定表明, DH155无论在苗期还是成株期都对白粉病表现高抗, 农艺性状良好, 并用作抗源进行抗病品种的选育。本研究主要是对DH155苗期抗E09基因进行遗传分析, 利用SSR分子标记技术对其抗病基因进行遗传定位, 并利用已发表的中国春和粗山羊草D基因组序列开发与抗病基因紧密连锁的SSR标记, 构建出紧密的连锁图谱; 同时, 利用紧密连锁标记将抗白粉病基因MlDH155辅助导入优良小麦品系中, 为今后开展抗性育种提供分子和材料基础。
高抗白粉病小麦品系DH155, 高感白粉病小麦品系山农2890 (SN2890)均系本实验室保存。以抗白粉病小麦品系DH155为父本, 以高感白粉病小麦品系SN2890为母本, 配制杂交组合。其杂种F1自交得到F2, F2单株自交得到143株F2:3家系, 对F1、F2和F2:3进行抗病性鉴定, 以高感普通小麦品种辉县红及农艺性状优良但感白粉病的品系HB133-4和旱10为感病对照材料。Ulka/8* Cc和Tabasco分别为Pm2和Pm46的载体品种[12, 13]。
将抗病亲本DH155、感病亲本SN2890、感病对照辉县红及抗、感双亲后代(每一个F2:3家系选20个F3单株)均匀种在花盆里, 放在光照培养箱内, 生长条件控制为白天21℃, 18 h; 夜间温度19℃, 6 h, 湿度100%, 光照40 µ mol m-2 s-1。用普通小麦辉县红扩繁白粉病菌种, 待鉴定材料于一叶一心期用抖拂法接种白粉菌菌株E09 (由中国农业科学院作物科学研究所李洪杰研究员提供)。为保证接种充分, 重复多次接种。接种后9~14 d (即感病亲本和感病对照材料充分发病后), 观察记录表型。采用6级分类方法[14]逐棵鉴定单株抗病性, 其中抗病等级0~2型归为抗病类型, 3~4型归为感病类型。根据F2:3家系表型来验证F2基因型, 根据F1和F2代中的抗感分离比例推断抗病基因数目和遗传类型。
采用CTAB法提取小麦基因组DNA[15]。结合F2:3家系表型鉴定结果, 分别选取F2代10个纯合抗病单株的DNA和10个纯合感病单株的DNA并等量混合成抗病池(BR)和感病池(BS)。用集群分离分析法(BSA)检测抗病基因与标记的连锁程度。
根据已经筛选到的与抗病基因连锁的标记Xcfd81序列(http://wheat.pw.usda.gov/cmap/), 与中国春[16]和粗山羊草D基因组序列[17]比对, 选择同源性大于96%的scaffold序列。通过SSR Hunter 1.3软件寻找序列中的SSR, 并用Primer Premier 5.0设计引物。所开发引物均由生工生物工程(上海)有限公司合成。
SSR-PCR体系为15 μ L, 包含2× Taq MasterMix 7.5 μ L, Primer-F 1.25 μ L, Primer-R 1.25 μ L, 去离子水3.0 μ L, 基因组DNA 2.0 μ L (50 ng μ L-1)。DNA扩增程序为94℃预变性5 min, 94℃变性30 s, 50~60℃退火30 s (根据引物设置), 72℃延伸30 s, 32个循环; 最后72℃延伸10 min。将扩增产物在8%的聚丙烯酰胺非变性凝胶上电泳2.5 h, 银染显影后用Alpha Innotech V.1.2型凝胶成像系统照相观察并统计带型。
结合F2分离群体的表型进行连锁分析构建遗传图谱。用JoinMap 4.0计算分子标记与抗病基因之间的重组率, 用Kosambi[18]算法将重组率转换成遗传距离, LOD值为3.0[19], 最大连锁交换率为0.5, 采用Mapchart作图。
为了改良普通小麦品系HB133-4和旱10对白粉病的抗性, 加快抗病基因MlDH155在育种中的利用, 本研究配置了2个杂交组合HB133-4 × DH155和旱10× DH155, F1自交得F2, 在F2~F4代, 结合分子标记辅助选择和优异农艺性状选择, 选择纯合稳定的优异株型, 并对其进行苗期抗病性鉴定, 表型鉴定方法同1.2节, 分子标记鉴定方法同1.3节。
抗病亲本DH155对白粉菌菌株E09表现免疫, 感病亲本SN2890高感E09。以抗病材料DH155为父本, 感病材料SN2890为母本杂交得到F1, F1自交得到F2, 在培养箱内用E09接种F1和F2单株进行抗病性鉴定。F1单株对白粉菌病菌E09全部表现免疫; 在143株F2群体中出现抗感分离, 其中有110株表现抗病, 33株表现感病, 经χ 2检验抗感分离比符合孟德尔单显性基因的分离比3∶ 1; 在143个F2:3家系中, 纯合抗病单株35株, 分离单株73株, 纯合感病单株35株, 其分离比符合1∶ 2∶ 1 (表1)。这表明DH155苗期抗E09基因受1对显性基因控制, 暂时将该抗白粉病基因命名为MlDH155。
| 表1 DH155与SN2890及其F2和F2:3家系对白粉菌菌株E09的抗性遗传分析 Table 1 Reactions to Blumeria graminisf. sp. triticiisolate E09 in the parents DH155 and SN2890 and their F2 and F2:3 progenies |
选用均匀分布在小麦基因组DNA上的412对小麦SSR引物, 共获得95对在抗病亲本DH155和感病亲本SN2890之间呈现多态性的引物。利用抗、感混合池筛选在亲本之间有多态性的95对引物, 发现位于小麦5D染色体短臂上的标记Xcfd81和Xcfd18均能够在抗病池和感病池扩增出稳定的多态性片段。进一步将Xcfd81和Xcfd18在10株纯合抗病和10株纯合感病的小群体上验证, 显示, Xcfd81和Xcfd18均与目标基因MlDH155连锁。然后, 将Xcfd81和Xcfd18在F2分离群体进行连锁分析, 发现这2个标记都与MlDH155连锁, 且Xcfd81和Xcfd18与抗MlDH155的遗传距离分别为2.0 cM和36.5 cM。Xcfd81和Xcfd18均为共显性标记, 二者在F2代分离群体中分离比均符合1∶ 2∶ 1 (表2)。由于SSR标记Xcfd81和Xcfd18都位于5D染色体短臂上, 因此, 将抗白粉病基因MlDH155定位在小麦5D染色体的短臂上。
| 表2 SSR标记在DH155× SN2890 F2群体分离情况及其与MlDH155的遗传距离 Table 2 Segregation ratio of SSR markers linked to powdery mildew resistance geneMlDH155 in F2 population derived from DH155 × SN2890 |
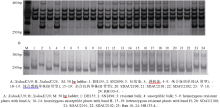
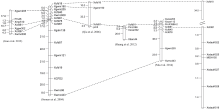
为了构建抗白粉病基因MlDH155更紧密的遗传连锁图谱, 从GrainGenes 2.0 (http://wheat.pw.usda. gov/cmap/)获得Xcfd81序列, 并用其与中国春(http:// plants.ensembl.org/Multi/enasearch)和粗山羊草D基因组(http://ensembl.gramene.org/Aegilopstauschii/Info/ Index)序列进行blastn搜索, 从中选择同源性96%以上的scaffold序列设计SSR引物。在抗、感亲本和抗、感混合池间筛选新开发分子标记的多态性。结果发现, 标记XsdauK519、XsdauK522、XsdauK525、XsdauK527和XsdauK528在抗病亲本和感病亲本及抗病池和感病池间均能扩增出明显的多态性DNA片段(表2、表3和图1)。经F2分离群体验证, 这5个标记均与抗病基因MlDH155存在紧密连锁关系。其中, XsdauK525为显性标记, 位于MlDH155的近端粒侧, 与MlDH155的遗传距离仅为0.2 cM; XsdauK527为共显性标记, 位于MlDH155近着丝粒侧, 与MlDH155的遗传距离为0.8 cM。最终, 将抗病基因MlDH155定位在显性标记XsdauK525和共显性标记XsdauK527之间, 其遗传距离分别为0.2 cM和0.8 cM (表2和图2)。
| 表3 新开发5DS染色体的SSR引物序列 Table 3 Primer sequences of SSR markers on chromosome 5DS developed in this study |
 | 图1 XsdauK519和XsdauK528扩增结果Fig. 1 PCR profile of XsdauK519 and XsdauK528 |
 | 图2 抗白粉病基因MlDH155和5DS上的其他抗白粉病基因的分子标记连锁图谱比较Fig. 2 Linkage map of MlDH155 and its comparison with the known Pmgenes on chromosome 5DS |
苗期活体鉴定结果表示, DH155对14个菌种中的13个菌种表现抗病(反应型0或1), 仅对来自河南新乡的Bg78-3菌株表现中感(反应型3)。Ulka/8* Cc (Pm2)仅对来自山东沾化的Bg44-5菌株表现感病, 而Tabasco (Pm46)除有2个菌种反应型无法确定外, 其余反应型均与Ulka/8* Cc (Pm2)相似。DH155和Ulka/8* Cc (Pm2)相比, 对Bg78-3和Bg44-5菌株反应型不同, DH155对Bg78-3感病, 对Bg44-5表现抗病, 而Ulka/8* Cc (Pm2)则正好相反(表4)。
| 表4 DH155 (MlDH155)与对照Ulka/8* Cc (Pm2)、Tabasco (Pm46)对白粉菌菌株的反应型的苗期鉴定 Table 4 Reaction patterns to Blumeria graminisf. sp. triticiisolates between DH155 (MlDH155) and control genotypes Ulka/8* Cc (Pm2) and Tabasco (Pm46) using seedling test |
以感白粉病普通小麦品系HB133-4和旱10作为母本, 分别与DH155杂交, 结合农艺性状和分子标记辅助选择, 从F2~F4后代中获得3个农艺性状优良的稳定株系。其中SDAU2100和SDAU2101来自HB133-4 × DH155, SDAU2102来自旱10 × DH155。从这3个纯系中分别随机挑选60个单株, 在一叶一心期用E09接种鉴定, 结果显示, 这3个纯系均对E09表现高抗(表5)。这表明在本研究中所开发的标记可以有效用于抗病基因MlDH155的分子标记辅助育种, 大大缩短了育种进程。另外, 主要农艺性状鉴定发现, 3个后代品系均在千粒重方面显著优于亲本HB133-4和旱10。本研究培育的抗白粉病新品系SDAU2100、SDAU2101和SDAU2102可作为培育抗病小麦品种的有效抗源。
| 表5 选育出的抗白粉病株系主要性状表现 Table 5 Main characteristics of the powdery mildew resistance lines selected in this study |
经过多年接种鉴定, DH155对白粉病流行菌株均表现高抗, 并且农艺性状良好, 适应性广, 是一个很好的抗病育种的抗源。因此, 对其抗病基因进行定位, 开发紧密连锁的分子标记, 并通过分子标记辅助选择实现对其抗病基因遗传转移是十分必要的。目前, 发现在5D染色体上的抗白粉病基因有Pm34[20]、Pm35[21]、Pm-M53[22]、Pm2[23, 24, 25]、Pm46[13]、PmLX66[26]和PmX3986-2[27]等。其中, Pm34、Pm35和Pm-M53均被定位在5D染色体的长臂上, 与其紧密连锁的标记分别为Xbarc177、Xcfd26和Xwmc289[20, 21, 22], 所以MlDH155应该是不同于Pm34、Pm35和Pm-M53的抗白粉病基因(图2)。
来自德国普通小麦Tabasco的抗白粉病基因Pm46被定位在Xgwm205和Xcfd81两个标记之间, 其遗传距离分别为17.6 cM和3.1 cM, 且通过等位点测验证明与Pm2不是同一基因[12]。PmLX66和PmX3986-2均来自我国普通小麦品种或品系, 且都位于5D染色体的短臂, 并都被报道可能与Pm2是等位基因[26, 27]。来源于粗山羊草的Pm2最初是由Pugsley和Carter从德国小麦Ulka中发现的, 并将其定位在5D染色体的短臂上[23, 24], 前人对其进行了大量研究。Qiu等[25]利用中国春与CI14118 (含有Pm2)衍生的814个F2分离群体进行SSR分析, 发现Xcfd81、Xgwm190和Xcfd18均与目标基因连锁, 与目的基因的遗传距离分别为2.0、34.2和44.2 cM。而我们发现SSR标记Xcfd81和Xcfd18与MlDH155连锁, 其遗传距离分别为2.0 cM和36.5 cM, 与Qiu等[25]定位的结果相似, 且Xcfd81能在DH155中扩增出与携带Pm2材料大小相近的片段。此外, 通过多菌株抗性鉴定表明, DH155与Ulka/8* Cc(Pm2)对用于分小种鉴定的其中的11个菌株抗性相似, 仅对Bg78-3和Bg44-5两个菌株反应型不同。MlDH155与Pm2基因对不同白粉菌菌株反应型的差异, 可能是由于不同的遗传背景造成的。因此, 我们推测DH155携带的抗白粉病基因可能与Pm2位于同一个基因座, MlDH155可能与Pm2是一个基因或者是等位基因(图2), 需要通过等位点测验进一步确定。
曾报道的小麦抗白粉病基因很多, 但其中有的已被证实是等位基因。例如, Pm18和Pm22被证实均为Pm1的等位基因, 因而又被命名为Pm1c和Pm1e[28, 29]。目前发现, Pm1共有5个等位基因(Pm1a至Pm1e), Pm3有10个等位基因(Pm3a至Pm3j), Pm4有4个等位基因(Pm4a至Pm4d), Pm5有5个等位基因(Pm5a至Pm5e)[30, 31]。本研究从DH155中发现一个苗期抗白粉病基因位点, 对其进行了遗传定位, 并将其命名为MlDH155。该抗白粉病基因与PmLX66、Pm2、PmX3986-2和Pm46都位于5D染色体短臂, 且遗传距离较近, 它们可能为等位基因或紧密连锁基因。
本研究利用抗白粉病品系DH155与农艺性状优良但感白粉病的品系HB133-4和旱10杂交, 在杂交后代中结合农艺性状选择、分子标记辅助选择和抗白粉病鉴定, 获得3个农艺性状优良且抗白粉病的稳定优良株系(SDAU2100、SDAU2101和SDAU2102), 目前正在对这3个品系进行产量潜力比较试验。这些结果进一步验证了我们所开发的与抗白粉病基因MlDH155紧密连锁的分子标记可以用于小麦抗白粉病分子标记辅助育种。
普通小麦品系DH155苗期对白粉病病菌E09表现高抗, 且对E09菌种的抗性受1对显性基因MlDH155控制。该基因被初步定位在小麦5D染色体的短臂上, 其双侧邻近标记为XsdauK525和XsdauK527, 遗传距离仅为0.2 cM和0.8 cM, 可用于分子标记辅助选择。分子作图及多菌株抗谱分析表明, MlDH155很可能是Pm2或其等位基因。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|