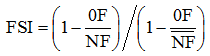
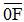


两种施肥环境下冬小麦京411及其衍生系产量和生理性状的遗传分析
[肖永贵1  , 李思敏
, 李思敏1 , 李法计1 , 张宏燕2 , 陈新民1 , 王德森1 , 夏先春1 , 何中虎1, 3, * ]
, 李思敏]
|
|


第一作者联系方式: E-mail:xiaoyonggui@caas.cn
明确高产广适性与肥料高效利用相关性状的遗传特性对培育优良新品种具有重要意义。本研究以京411及其衍生后代共15个品种(系)为材料, 在4个正常施肥和1个常年不施肥环境下研究品种(系)的产量构成因素和生理性状, 结合90K SNP芯片, 解析骨干亲本携带的产量和生理性状等位基因信息, 探讨优异基因对高产品种的贡献。结果表明, 正常施肥环境下, 京411衍生后代的产量和收获指数均随世代增加呈逐渐上升趋势, 其中收获指数增加较为显著( P < 0.05)。冠层温度对籽粒产量有重要贡献( P < 0.05), 而叶面积指数和光合速率对收获指数有显著贡献( P < 0.05)。两种施肥条件下中麦175具有较高且稳定的产量和生物学产量, 主要与其较高的肥料吸收效率有关。控制产量和生理性状的遗传区段主要分布在A和B基因组上, 2B、3A和5A染色体分别携带控制穗粒数、叶面积指数和光合速率的位点。京411携带31个对产量和生理性状为正向效应的等位基因, 衍生品种CA0958和中麦175携带的正向效应区段最多, 分别占正向效应位点总数的53.85%和51.35%。中麦175的高产潜力和广适应性可能与其携带较多有利等位基因有关。
, LI Si-Min
Understanding the genetics of the traits related to yield potential, stability and high fertilizer-use efficiency is important for breeding new varieties. The elite parent Jing 411 has been widely used in wheat breeding programs in Northern Winter Wheat Region. In this study, Jing 411 and its 14 derivatives were sown in normal fertilized and no-fertilized environments to assess the changes in yield stability and fertilizer-use efficiency. Grain yield and harvest index increased following the generation advance, particularly harvest index increased significantly ( P < 0.05). Zhongmai 175 showed high and stable grain yield and above ground biomass under two fertilized environments, mainly due to its high fertilizer-uptake efficiency. Canopy temperature was significantly associated with grain yield ( P < 0.05), and leaf area index, and photosynthetic rate were significantly correlated with harvest index ( P < 0.05). Several important loci associated with yield and physiological traits were found in A and B genomes. Three new loci for grains per spike, leaf area index, and photosynthetic rate were detected on chromosomes 2B, 3A, and 5A, respectively. Furthermore, 31 positive genetic loci for yield and physiological traits were detected in elite parent Jing 411, and two derivatives, CA0958 and Zhongmai 175, had the most positive loci than others, accounting for 53.85% and 51.35%, respectively. Zhongmai 175 had higher yield potential and stability under different fertilizer environments, possibly due to the presence of positive loci for yield and physiological traits.
提高品种的产量潜力及肥料利用效率是作物育种的重要发展方向, 其优势主要体现在提高种植效益, 节约资源投入, 减少环境污染[1, 2]。不同基因型在有效利用土壤潜在养分特性方面存在显著差异[3], 高肥效品种在营养缺乏的条件下, 也可以通过一系列形态、生理和生化机制活化土壤中的营养元素, 从而提高土壤中潜在肥料的生物有效性[2]。小麦高肥效特性主要表现为植株对土壤固有营养的高效利用, 营养元素在植株体内的合理分配和高效转运, 并使机体保持最优的光合势和碳同化效率, 从而获得较高的生物产量和籽粒产量[4]。因此, 品种的高肥效特性可进一步区分为高效吸收型和高效转化型。不同小麦品种的肥料吸收效率和转化效率存在显著差异, 在低肥环境下, 墨西哥和芬兰的春小麦具有较高的肥料吸收效率, 而法国和英国的冬小麦具有较高的肥料转化效率; 在高肥环境下, 英国、墨西哥和芬兰小麦的吸收和转化效率基本一致[3, 5, 6]。Gaju等[2]认为, 土壤肥料供给量与小麦品种存在显著互作关系, 在低肥条件下更能有效鉴定品种的肥料吸收效率。
有关产量性状基因定位的报道很多, 几乎每个遗传背景下都会定位到数十个产量相关性状的基因位点, 而且多数位点不尽相同。已发掘的产量性状基因位点的遗传效应相对较小(2%~10%), 对表型变异的解释率(R2)大于15%的主效基因很少[7, 8, 9, 10]。主要原因有二: 一是环境因素诱导产量及其相关性状之间存在相互协调或此消彼长的关系[11, 12]; 二是影响产量及其相关性状的基因不仅具有主效效应, 而且还受基因× 基因、基因× 环境等复杂网络关系的共同影响[7]。目前, 关于产量性状的遗传研究不足以解析其性状和基因间的关系, 仍需要在多种试验环境下发掘产量性状基因及其协同效应[9, 10]。另外, 理想的基因定位研究应在一个能够造成产量相关性状差异较大的环境(如干旱胁迫)下进行, 尤其是遗传力相对较低的目标性状(如生理性状), 这样可降低基因与环境的互作效应, 更容易鉴定出目标性状的主基因效应及基因间的互作关系[8]。在多种环境下均稳定表达的产量相关性状基因更受育种家的青睐[12], 因为它决定品种的广适性和稳产性。
京411于1991年通过北京市品种审定, 具有高产稳产、适应性广等突出特点, 在北部冬麦区推广近20年, 累计种植74.3万公顷[13]。从该品种育成至21世纪初, 京411一直是北部冬麦区高产广适育种的骨干亲本, 已育成16个衍生品种(系), 包括中麦175、北京0045等本麦区的主栽品种[14]。本研究以冬小麦骨干亲本京411及其14个衍生后代为材料, 在正常施肥环境和常年不施肥环境下研究不同基因型的产量构成因素及相关生理性状的变化, 并结合全基因组SNP标记分析该衍生群体的产量、生理性状遗传区段, 为今后利用骨干亲本培育更优良的新品种提供理论基础。
京411是20世纪90年代初育成的半矮秆小麦品种, 具有抗寒性强、分蘖力强、成穗率高、高产稳产、抗病性好、适应性广等特点, 本课题组针对其植株偏高、灌浆较慢、落黄差等缺点进行改造, 育成的衍生品种(系)多数具有高产广适的特性, 其中北京0045、中麦175等品种已在北部冬麦区生产中发挥重要作用。本研究选用骨干亲本京411及其14个衍生品种(系)为材料(表1), 包括衍生子一代6份、子二代8份, 这些材料基本上反映了当前北部冬麦区小麦品种的产量现状。
| 表1 供试品种(系)的矮秆基因型、抽穗期和株高 Table 1 Rht-B1a/Rht-B1b and Rht-D1a/Rht-D1b genotype, heading date, and plant height of varieties/lines used in this study |
2012— 2013和2013— 2014年度, 将15份品种(系)种植于中国农业科学院作物科学研究所顺义试验站(40.13° N, 116.57° E)和天津市武清区种子管理站农场(39.50° N, 117.00° E), 北京试验点的播期为2012年9月28日和2013年9月29日, 天津试验点的播期为2012年10月20日(因前茬玉米收获晚, 导致小麦迟播)和2013年9月26日。仅2013— 2014年度顺义试验站设置正常施肥和全生育期不施肥两组试验。各试验点播种时施纯N 180 kg hm-2、P2O5 120 kg hm-2和K2O 60 kg hm-2, 拔节期追施氮肥约 150 kg hm-2。均采取随机区组设计, 3次重复, 小区行距20 cm, 行长4 m, 面积为5.6 m2, 种植密度为300株 m-2。全生育期浇越冬水、拔节水和灌浆水共3次, 并控制病虫草害。
测定了两种试验条件下产量及其相关性状。越冬前, 在小区内出苗均匀且长势旺盛的非边行区域选定2个1 m样段, 调查基本苗。春季拔节前调查样段的最大分蘖, 收获前记录样段穗数, 并折算成单位面积穗数。成熟期在小区内随机选取3点, 量取株高, 计算平均株高; 在非边行区随机选取20穗, 计算穗粒数, 折算成每穗粒数。收获前从根部拔取收获每小区的样段, 剪去根部, 置于80℃烘箱烘干至恒重, 测定生物学产量, 样品穗单独脱粒计算收获指数。田间记载抽穗期和成熟期。小区收获后晾晒干燥至含水量为12%时称重, 测定千粒重。根据小区收获产量和样段的籽粒重量的总合计算供试品种的公顷产量。
本试验仅在正常施肥条件下测定生理性状。采用GreenSeeker冠层光谱仪(Trimble, 美国)分别在越冬前、起身期、拔节期和灌浆期采集植被归一化指数(normalized difference vegetation index, NDVI), 测定时传感器与小麦冠层60 cm高度上方保持平衡, 沿播种方向测定, 在小区非边行区域往返测定2次。用REYTEK ST20XB型手持式红外测温仪(Reytek Corporation, 美国)测定冠层温度, 光谱通带为8~14 μ m, 灰度值ε = 0.95, 选择抽穗期晴朗无风少云的天气, 于午后13:30— 15:00测定一次[15]。开花期和灌浆中后期在小区非边行区域随机选择10片旗叶, 利用SPAD 502Plus叶绿素仪(Konica Minolta Inc., 日本)测定叶绿素相对含量, 10片旗叶的均值作为被测群体的叶绿素相对含量。开花期在每个小区非边行区随机选择10片旗叶, 用YMJ-B叶面积测定仪(浙江托普仪器有限公司)测定叶面积, 其均值作为被测群体的旗叶面积。利用AccuPAR叶面积指数仪(Decagon Devices Inc., 美国)测定群体冠层上下的入射光和透射光强度, 并计算群体的冠层LAI值。另外, 灌浆初期利用Li-6400XT光合仪(LI-COR, 美国), 在恒定光源1200 μ mol m-2 s-1和二氧化碳400 μ mol mol-1条件下, 测定每小区5片旗叶的光合速率, 其均值视为被测群体的旗叶光合速率。
从每份材料选取10个大小和色泽均匀一致的籽粒在培养皿中发芽2周, 混合剪取幼苗, 用澳大利亚Triticarte公司提供的CTAB法提取小麦全基因组DNA (http://www.diversityarrays.com/)。利用UVS-99微量分光光度计(ACTGene, 美国)检测DNA质量和浓度, 将DNA终浓度调为300 ng µ L-1。
利用Illumina SNP基因分型研究平台对京411及其衍生品种(系)DNA进行90 K SNP芯片分型, 包括BS、BobWhite、CAP、D_contig等系列标记, 共计81 587个, 其中38 819个SNP标记在京411衍生群体内存在差异, 已有染色体定位信息的标记为18 011个[16]。本研究仅选取有染色体定位信息的标记进行分析, 并去除掉重复标记, 剩余4846个标记对产量和生理性状进行遗传区段检测分析。
对15个品种的产量和生理性状的方差分析表明(表2), 除越冬前和灌浆期的NDVI之外, 其他性状均受基因型、环境和基因型× 环境的显著影响(P < 0.05)。由于越冬前和灌浆期的NDVI在品种间差异不显著, 不再进行后续分析。冠层温度和旗叶光合速率在相同环境下的重复间存在显著差异(P < 0.05), 可能是这两个性状在测定时受环境的温度和CO2浓度变化影响所致。
| 表2 正常施肥条件下京411及其衍生品种的产量和生理性状方差分析 Table 2 Analysis of variance for yield and physiological traits in elite parent Jing 411 and its derivatives under normal fertilizer environments |
2012— 2013年度, 供试材料在北京点产量平均为5671.5 kg hm-2, 在天津点的平均产量为5164.8 kg hm-2; 2013— 2014年度, 两点产量介于二者之间, 全生育期不施肥处理的平均产量为2840.7 kg hm-2。2012— 2013年度北部麦区冬季降雨量达108 mm, 播种期土壤墒情好, 气温偏高, 出苗均匀, 有利于分蘖形成; 春季持续低温(≤ 5℃)直至3月中下旬, 抽穗后低温时间长, 生育期较往年推迟5~7 d, 灌浆期相对较长, 因而产量较高。2012— 2013年度天津武清试验点产量偏低的主要原因是播期偏晚。2013— 2014年度的有效积温和降雨量与历年相近, 冬前温度适宜, 春季气温回升较往年快, 但灌浆中后期(5月26日至31日)遇到高温天气, 气温高达42℃, 成熟期较往年提早7 d左右。
正常施肥条件下衍生子代的每平方米穗数与京411接近, 穗粒数随世代更迭略呈增加趋势, 千粒重和株高有所降低(表3), 收获指数和产量随世代演变均呈递增趋势, 其中衍生二代的收获指数和产量显著高于骨干亲本京411 (P< 0.05)。说明在对京411的产量性状改良过程中, 基本保持了单位面积有效穗数, 提高穗粒数, 进而提高收获指数和籽粒产量。
| 表3 两种施肥环境下京411及其衍生品种的产量及其构成因素、株高和肥料敏感性分析 Table 3 Analysis of yield and its components, plant height and fertilizer susceptibility in Jing 411 and its derivatives under two treatments |
在正常施肥条件下, 中麦175、中麦818、CA1055、中麦816和CA1119产量较高(5741.9~5882.6 kg hm-2); 在不施肥条件下, CA1119和中麦175同样具有较高的产量, 分别为3237.8 kg hm-2和3135.1 kg hm-2。在产量较高的品种中, CA1055的产量、穗粒数、千粒重和株高均对肥料的敏感性较高, 敏感指数在1.05~1.57之间, 中麦818和中麦816的产量和穗数对肥料敏感性较高, 敏感指数在1.04~1.34之间(表3)。虽然CA1119的产量对肥料表现不敏感(0.90), 但穗数(1.22)和穗粒数(1.06)对肥料敏感性较高。而中麦175的产量、产量构成因素和株高均对肥料表现不敏感, 敏感指数在0.70~0.97之间, 说明中麦175的产量及其相关性状在不同肥料环境下均具有较好的稳定性。
在正常施肥条件下, 中麦175在苗期和成熟期均具有较高的生物学产量(拔节期NDVI值为0.50, 成熟期生物量为16.94 kg m-2, 穗数为741.25 m-2), 而旗叶面积(17.28 cm2)、叶绿素含量(SPAD值49.04)和冠层温度(19.72℃)其他品种较低。在不施肥环境下, 中麦175仍具有较高的生物量(8.80 kg m-2), 仅次于中麦415 (9.80 kg m-2)。可见, 中麦175在不同肥料条件下均具有较高的肥料吸收效率, 能保持较高的生物量和籽粒产量, 这可能与其株型直立、有效穗数较多、群体蒸腾效率高有关。
在正常施肥条件下, 每平方米穗数与起身期群体最大分蘖数(r = 0.72, P < 0.01)和拔节期NDVI (r = 0.53, P < 0.05)呈显著正相关(表4), 而与叶绿素含量(r = -0.54, P< 0.05)、旗叶面积(r = -0.52, P< 0.05)和光合速率(r = -0.54, P < 0.05)呈显著负相关, 说明苗期群体量对发育后期的单位面积穗数有重要影响, 而随着群体量的增加, 旗叶光合作用能力呈下降趋势。灌浆期旗叶光合速率与穗粒数(r = 0.56, P < 0.05)和收获指数(r = 0.59, P< 0.05)呈显著正相关, 灌浆期旗叶的光合产物主要作用于籽粒灌浆, 进而提高收获指数。此外, 生物量与产量呈极显著正相关(r = 0.70, P < 0.01), 灌浆初期的冠层温度与产量呈显著负相关(r = -0.61, P< 0.05), 冠层温度低意味着植物群体的水分吸收和蒸腾系统高效, 生理代谢旺盛, 具有群体高光效能力, 利于光合产物的积累和转化, 进而影响籽粒产量。
| 表4 正常施肥条件下生理性状与产量及其构成因素的相关分析 Table 4 Correlation coefficients of grain yield and its components with physiological traits under normal fertilizer environments |
在4846个SNP标记中筛选出70个与产量和生理性状相关的标记, A、B基因组携带的位点较多, 分别为29个和36个, D基因组仅有5个位点。2B和5A染色体分布最多, 分别为10个和9个, 其中2B携带的位点主要与穗粒数、千粒重和产量等性状相关, 而5A携带的位点多数与生理性状相关, 如旗叶叶绿素含量、光合速率和株高等(表5)。控制小麦NDVI的位点主要与1B、4A、4B和5B染色体有关, 而控制叶绿素含量的位点主要位于4A、4D、5A和5B染色体。在3A染色体的80 cM区段附近和5A染色体的300 cM区段附近分别发掘到3个与叶面积指数和光合速率相关联的SNP位点, 这两个遗传区段可能参与小麦生理性状调控。另外, 在2B染色体发掘到4个与穗粒数相关的SNP位点(表5), 位于染色体短臂近着丝点附近的240 cM区域, 该区域可能携带穗粒数相关基因。
| 表5 京411及其衍生品种的产量和生理性状遗传区段分析 Table 5 Genetic loci for grain yield and physiological traits in Jing 411 and its derivatives |
在发掘的产量和生理性状关联区段中, 衍生一代获得骨干亲本的遗传位点较高, 为51.43%, 衍生二代相对较低, 为43.75%, 正向效应遗传位点也在衍生一代和二代之间存在差异, 分别为46.12%和40.50% (表6)。不同品种间携带与骨干亲本一致的位点存在较大差异, 如中麦415、CA0958和中麦175携带与京411相同位点较多, 所占比例均在50%以上; CA1090与骨干亲本的相同位点最少, 占京411位点数的37.14%。在发掘的70个遗传区段中, 对产量和生理性状起正向效应的区段为31个, 占总区段数的44.29%, 其中CA0958和中麦175携带正向效应区段最多, 分别占骨干亲本贡献位点总数的53.85%和51.35% (表6), 说明这两份基因型获得较多的骨干亲本优异产量和生理性状遗传成分。
| 表6 京411及其衍生品种的多效性标记遗传位点差异 Table 6 Difference of alleles linked with pleiotropic SNP markers among Jing 411 and its derivatives |
理想的小麦基因型应在多种环境下具有较高且稳定的产量, 并表现较高的水肥利用效率[1]。育种过程中通常将世代材料和稳定品系在肥水充足的环境下进行产量鉴定, 但由于基因型与肥料用量存在互作关系, 很难鉴定出品种间肥料利用效率的差异[3, 6]。本研究表明, 生物量对籽粒产量有重要贡献, 尤其在低肥条件下, 生物量高的品种一般籽粒产量较高(r = 0.70, P < 0.01)。在京411衍生后代中, 中麦175在两种肥料条件下均具有较高的肥料吸收能力, 表现较高的生物量和产量, 而骨干亲本京411具有较高的肥料转化能力, 低肥环境下具有最高的收获指数(0.46)。本研究还发现, 低肥环境下的千粒重显著高于正常施肥条件下的, 可能原因是低肥条件下单位面积穗数少, 群体内竞争力小, 光温通透, 植株光合产物能够充分向籽粒转化, 进而提高粒重。
中麦175是骨干亲本京411最成功的改造品种, 具有较好的丰产性、稳产性和广适性, 分别通过北部冬麦区水地和黄淮旱肥地品种审定, 也是北部冬麦区国家区域试验的对照品种, 近几年来一直是北部冬麦区种植面积最大的品种[14]。该品种不仅具有较强的耐低磷[18]和氮肥利用效率(童依平, 个人交流), 而且水分利用效率高。2012— 2013和2013— 2014年度对100个国家审定的冬小麦品种在无灌水、灌1水和灌2水条件下进行比较, 中麦175在灌1水的条件下产量最高(郭进考, 个人交流)。该品种的节水性可能是其在河南、陕西和甘肃陇东等旱肥地区大面积推广种植的主要原因。
在京411衍生群体中共发掘70个控制产量和生理性状的遗传区段, 其中2B染色体携带较多控制穗粒数、千粒重和产量的遗传区段(表6), 5A染色体上控制叶绿素含量和光合速率等生理性状位点较多。已有研究证实, 5A染色体携带较多控制小麦环境适应性、抗病性和驯化相关的基因[19, 20], 最重要的2个基因是春化基因(Vrn1)和易脱粒基因(q), Vrn1基因除参与调控小麦春化作用之外, 也可能参与株高、穗数和穗粒数等产量相关性状的调控[20]; q基因同时具有控制穗部形态和易脱粒性的作用[19]。Graziani等[21]发现, 2B染色体携带一个重要的控制千粒重、穗下节长、叶绿素含量和产量的基因, 且在8个试验环境下均稳定表达。另外, 在京411衍生群体中发现3A和5A短臂分别存在控制叶面积指数和光合速率相关基因位点, 这些影响生理性状的位点也可能参与植物发育[22]、叶片衰老相关性状[23]的调控。
此外, 本研究还检测到控制单个产量和生理性状的遗传区段, 分布在1A、1B、2A、2D、3A、3B、3D、4A、4B、4D、5B、6A、6B、7A、7B和7D, 可能这些区段的等位基因仅属于京411衍生种群体特有的遗传变异, 也可能是因群体数量小造成了等位变异的伪关联。然而, 利用全基因组关联分析时, 经常遇到单标记负责调控单一性状的结果, 可能是高密度的标记信息更能精准评估种质的单倍型模式, 并能够准确发掘出单倍型与性状表型的关系[24]。连锁定位表明, 分布在这些遗传区段的多数位点具有多效性, 同时参与产量和生理相关性状的调控。例如, 5B染色体NDVI位点, 不仅在灌溉和干旱胁迫下能够有效作用于苗期NDVI的表达, 而且对植株发育后期的叶片衰老机制有一定的影响[22]。3B染色携带的基因位点同时参与调控千粒重、生物学产量、叶片叶绿素含量和籽粒产量[21]。
骨干亲本京411具有突出的适应性和丰产性, 可能与其携带较多正向遗传效应基因有关(表6) [14, 18]。本研究表明, 京411携带31个产量和生理性状正向关联区段, 其衍生后代CA0958和中麦175携带骨干亲本贡献的正向基因位点最多。通常骨干亲本具备有利基因多、遗传传递力强等特点[14, 25, 26], 育种过程主要是保留其携带的有益等位基因, 并使其在多个环境下稳定表达。这也充分说明传统育种针对优异农艺性状为目标进行高强度选择, 可使优异目标性状基因有效聚集, 使亲本携带的正向遗传位点在衍生材料中增加。对京411衍生群体研究发现, CA0958是携带骨干亲本的根系性状[14]、肥料利用效率[18]、产量和生理性状正向遗传区段(表6)最多的品种, 然而, 在本试验和实际生产情况下, 该品系的产量均不突出, 且稳定性较差。而另外一个携带正向遗传区段较多的品种中麦175, 其产量和适应性明显优于其他品种。由此说明, 正向等位基因的富集是构成优良品种的遗传基础, 而优势等位基因的协同表达可能是提高品种高产和稳产性的重要因素之一。
在京411衍生群体中, 中麦175是改造骨干亲本较成功的范例。该品种在越冬前后幼苗旺盛, 生长势较强, 分蘖成穗率多, 单位面积穗容量多, 株型好, 叶面积指数较高(光截获面积大), 群体光能利用率高, 具有较高的生物学产量。此外, 高效肥料吸收效率和水分利用效率为该品种的稳产和广适性奠定了基础。这些优良的农艺性状表现可能与其携带较多正向遗传位点有关。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|