



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
基于生物量的油菜主茎叶片形态参数模拟研究
[张文宇, 张伟欣, 葛道阔, 曹宏鑫*  , 刘岩, 宣守丽, 傅坤亚, 冯春焕, 陈魏涛]
, 刘岩, 宣守丽, 傅坤亚, 冯春焕, 陈魏涛]
, 刘岩, 宣守丽, 傅坤亚, 冯春焕, 陈魏涛]
|
|


为了定量油菜主茎叶片形态参数与生物量间的关系, 本研究基于2011—2012年和2012—2013年不同品种、移栽密度及施肥水平油菜田间试验, 通过观测不同品种和处理油菜叶片长、最大叶宽和叶柄长等形态参数, 并分析了上述参数与叶片生物量的关系, 构建了基于生物量的油菜叶长、最大叶宽和叶柄长模型。结果表明, 在全生育期, 不同品种和处理下油菜主茎叶长和最大叶宽均与叶片生物量的平方根成正比, 而叶柄长与叶长成正比。所建模型利用截距为0的线性函数描述叶长和最大叶宽随生物量平方根的变化, 用直线式描述叶柄长随叶长的变化。经独立试验资料检验, 除宁油16叶柄长模型误差较大外, 所建模型对其余形态参数均具有较好预测性, 为通过生物量将油菜生长模型与形态结构模型结合提供了机理性较强的方法, 为建立油菜功能-结构模型奠定了基础。
, LIU Yan, XUAN Shou-Li, FU Kun-Ya, FENG Chun-Huan, CHEN Wei-Tao
To quantify the relationships between main stem leaf morphological parameters for rapeseed and the corresponding leaf biomass, we conducted field experiments on varieties, transplanting densities, and fertilizer in 2011 to 2012, and 2012 to 2013 seasons. The biomass-based leaf morphological parameter models for rapeseed on main stem were constructed through observing leaf length, maximum leaf width, and leaf petiole length under various treatments, and analyzing the relationships between leaf morphological parameters and the corresponding leaf biomass. Because both of the leaf length and leaf width were positively proportional to the square root of the corresponding leaf biomass, meanwhile the leaf petiole length was positively proportional to the corresponding leaf length, the changes in the leaf length, the maximum leaf width, and the leaf petiole length with the changed of square root of the corresponding leaf biomass for different treatments could be described with linear function. The biomass-based leaf morphological parameter models were validated using independent experiment data, and the results showed that the model revealed satisfactory predictions of leaf length, leaf width, and leaf petiole length, except the simulation for leaf petiole length of Ningyou 16. The research provides a mechanistic method for linking the rapeseed growth model with the morphological model using organ biomass, and lays a foundation for the establishment of functional-structural plant models of rapeseed.
油菜是世界重要的油料作物[1], 近十年常年种植面积2300~3300万公顷, 总产量5000~6000万吨[2]; 在中国油菜是唯一的冬季油料作物[3], 常年种植面积600~700万公顷, 总产量1000~1300万吨[2]; 油菜也是清洁的新型能源— — 生物柴油的主要原料之一[4]。因此, 促进油菜生产发展对保障国家食物安全具有重要意义。良种良法配套是实现油菜高产、优质、高效、生态与安全生产的根本保证, 而功能-结构油菜模型研究, 既可为油菜栽培的株型调控提供新手段, 也可为油菜育种的株型设计和筛选提供理论方法。
功能-结构植物模型(Functional-Structural Plant Models, FSPMs)是国际上作物形态模型与可视化研究的新趋势[5]。近年来, 以LIGNUM[6]、GREENLAB[7]和L-PEACH[8]等为代表的功能-结构植物模型, 已在水稻[9, 10, 11, 12]、小麦[13]、玉米[14]、棉花[15]等作物上实现。但目前关于油菜模型的研究多为单独的生长模型或形态模型, 如国外的EPIC-Rape[16]、DAISY- Rape[17]、LINTUL-BRASNAP[18]、CERES-Rape[19]及APSIM-Canola[20]模型; 国内张春雷等[21]、刘洪等[22]、廖桂平等[23]、刘铁梅等[24]、曹宏鑫等[25]和汤亮等[26]构建的油菜生长模型, 以及廖桂平等[27]、岳延滨等[28]、赵丽丽等[29]构建的油菜形态模型, 而有关功能-结构油菜模型的报道较少。Groer等[30]利用基于L-System的XL建模语言[31], 并借鉴LEAFC3-N模型[32]的光合部分和GREENLAB模型的源-库系统, 构建了油菜动态3D模型, 使形态模型可响应不同氮素水平, 但仅考虑了氮素的影响, 且不能从出苗开始模拟。Jullien等[33]利用GREENLAB明确了源-库关系与油菜形态之间的相互作用, 通过叶面积与生物量的关系, 构建了较完整的功能-结构油菜模型, 但其主要考虑叶面积与生物量的关系[34], 对叶片形态描述较简单, 更侧重于分枝描述。曹宏鑫等[35]量化了油菜叶片长宽和角度等指标与叶片干重的关系, 构建了基于生物量的油菜苗期叶片几何参数模型, 张伟欣[36]通过分析油菜生物量与器官形态间的关系, 构建了基于生物量的油菜越冬前植株地上部、分枝各器官形态结构模型, 但其对主茎叶形的模拟都仅适用于苗期。而适用于整个生育期且考虑肥料与移栽密度影响的油菜叶片几何参数模型还未见报道。
本研究旨在通过不同油菜品种、肥料和密度田间试验, 借鉴水稻叶片几何特征模型[10]思路, 分析不同处理油菜主茎叶片叶长和最大叶宽与干重间, 以及叶柄长和叶长间的定量关系, 构建基于生物量的叶片形态参数模型, 为使油菜生长模型与形态模型结合, 进而构建功能-结构油菜模型提供参考。
试验于2011— 2013年在江苏省农业科学院院部试验农场(32.03° N, 118.87° E)进行。前茬为水稻, 供试品种3个, 分别为宁油18 (常规, V1)、宁油16 (常规, V2)和宁杂19 (杂交, V3); 施肥水平分别为不施肥(N0)、施纯氮90 kg hm-2(N1)和施纯氮180 kg hm-2(N2); 移栽密度分别为6× 104株 hm-2(D1)、1.2× 105株 hm-2(D2)和1.8× 105株 hm-2(D3)。土壤含有机质31.4 g kg-1、全氮2.03 g kg-1、速效磷20.3 mg kg-1、速效钾139 mg kg-1, pH 7.31。2011— 2013年两生长季主要气象要素月均值见表1, 2011— 2012年度油菜生长季降水和日照均较2012— 2013年度少。
| 表1 油菜生长季气象要素月均值 Table 1 Monthly mean of meteorological elements during rapeseed growing season |
试验I为品种和肥料试验, 采用裂区设计, 主区为施肥, 设N0和N2两个水平, 副区为品种, 供试品种为V1、V2和V3, 小区面积7.0 m× 5.7 m=39.9 m2, 行距0.4 m, 株距0.17~0.20 m, 随机排列。施肥区于移栽前施入P2O512 kg hm-2、K2O 18 kg hm-2和硼沙15 kg hm-2, 氮肥按基肥:苗肥:腊肥=5:3:2分配。2011年10月15日播种, 11月4日移栽。
试验II为品种试验, 供试品种为V1、V2和V3, 按N1水平施氮、D2密度移栽。
试验III为品种、肥料和密度试验, 采用裂区设计, 主区为施肥, 设3个(N0~N2)水平, 副区为品种和移栽密度, 供试品种为V1和V2, 设3个(D1~D3)移栽密度水平。
试验II和试验III小区面积均为3.99 m× 3.50 m=13.97 m2, 行距0.42 m, 株距按密度计算, 随机排列, 移栽前N1小区施入P2O5和K2O各90 kg hm-2, N2小区施入P2O5和K2O各180 kg hm-2, 并于抽薹后叶面喷施硼沙15 kg hm-2, 氮肥按基肥:腊肥:薹肥=5:3:2分配。2012年10月8日播种, 11月9日移栽。
以上试验均为3次重复, 其他栽培措施同高产大田管理。
自三叶期起标记叶位。选择每处理3~5株长势相近的油菜, 开春前每隔15~30 d (其中, 移栽前仅考虑品种, 单独用V1和V3表示), 开春后每7~15 d, 观察油菜叶型, 用直尺测量叶长(叶片伸直状态下叶基部到叶尖的距离, 包括了长柄叶的柄长及叶片长)、叶柄长(仅指长柄叶的柄长)和最大叶宽。
采用Microsoft Excel 2013整理数据; 用SPSS v. 21进行统计分析。在分析叶位相关数据时, 采用归一化方法, 即将各处理实测叶位除以该处理主茎总叶数, 使其统一到(0~1]区间内, 消除表观叶位差异, 便于模型构建。
试验II、III的实测数据用于模型构建, 试验I的实测数据用于模型检验。
采用根均方差(root mean square error, RMSE)、平均绝对差值(mean absolute difference, da)、da与观察值平均数的比值[37](dap)及相关系数(r)对模型进行检验; 同时采用1:1关系图, 直观显示模型的精度和可靠性。RMSE、da和dap值越小, r值越大, 表明模拟值与实测值的一致性越好。


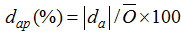
油菜叶片形态参数主要包括叶长、叶宽和叶柄长。由于短柄叶的叶柄和叶片界限较模糊, 因此, 本文模拟的叶长是叶片和叶柄(如果存在)的总长度, 叶柄长仅指长柄叶柄长。叶宽仅指叶片最大叶宽。
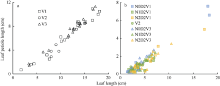
2.1.1 叶长模型 2012— 2013年试验数据表明, 不同处理下, 油菜主茎不同叶位叶片长度与最大宽度均成截距为0的线性关系(图1)。
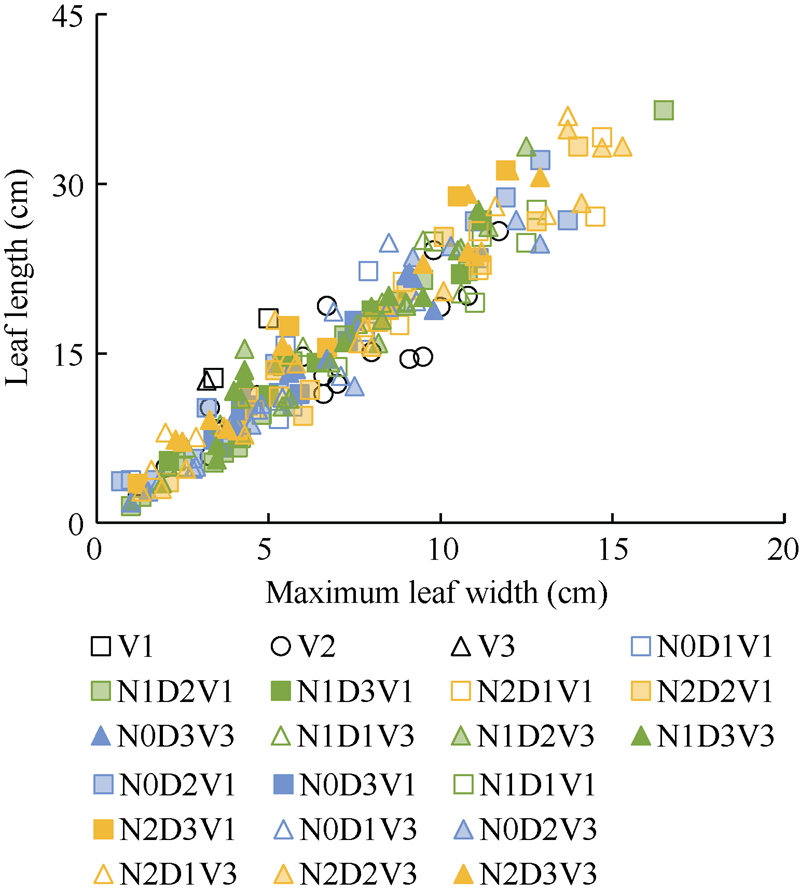 | 图1 2012-2013年不同处理叶长随最大叶宽的变化规律Fig. 1 Relationship between leaf length and maximum leaf width for different treatments from 2012 to 2013 |
根据上述规律以及比叶重和叶面积的计算方法可得(1)至(3)式。
DWLBj(i)=LAj(i)× SLW (1)
LAj(i)=LLj(i)× LWMaxj(i)× LMC (2)
LWMaxj(i)=LLj(i)/SLW (3)
式(1)~(3)中, DWLBj(i)为出苗后第i天主茎第j叶干重(g), 可由油菜生长模型[25]的生物量输出经干物质分配获得; LAj(i)为出苗后第i天主茎第j叶面积(cm2); SLW为比叶重(g cm-2); LLj(i)和LWMaxj(i)分别为出苗后第i天主茎第j叶叶片长度和最大宽度(cm); DWLBj(i); LMC为叶形系数; RLW为叶片长宽比。综合式(1)~(3)可推导叶重与叶片长度的关系为:
DWLBj(i)= LLj(i)2× SLW× LMC/SLW (4)
由于生物量是叶片生长的物质基础, 因此, 根据式(4)可得式(5)。
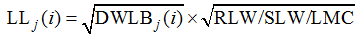
2012— 2013年试验数据也表明, 不同油菜品种、氮素和密度处理下, 主茎叶片在伸长过程中, 叶长随叶干重平方根呈线性上升趋势。结合试验数据和油菜叶形等因素, 将归一化叶位分为(0, 0.185]、(0.185, 0.375]、(0.375, 0.750]和(0.750, 1.000] 4个区间, 如图1所示, 各归一化叶位区间内叶长随叶干重平方根的变化规律均可用截距为0的线性函数拟合(统计参数见表2)。
| 表2 叶长模型及其参数显著性检验 Table 2 Significance test of leaf length model and its parameter values |
因此, 不同处理油菜出苗后第i天主茎第j叶叶片长度LLj(i)(cm)可统一表示如下。
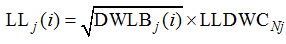
式(6)中, Nj为归一化叶位, 即叶位与主茎总叶数之比; 根据式(5)可知, LLDWCNj是归一化叶位为Nj时RLW/SLW/LMC的平方根, 此处命名为叶长重系数[cm ( 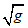
2.1.2 最大叶宽模型 与叶长类似, 由式(1)~(3)也可得出最大叶宽与叶片生物量的平方根成正比。

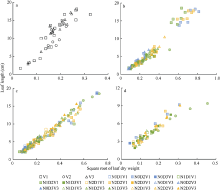
2012— 2013年试验数据也表明, 不同处理水平下, 最大叶宽随叶干重平方根的变化规律与叶长相似, 如图3所示, 在归一化叶位区间(0, 0.185]、(0.185, 0.375]、(0.375, 0.750]和(0.750, 1.000]内, 均可用截距为0的直线拟合(统计参数见表3)。
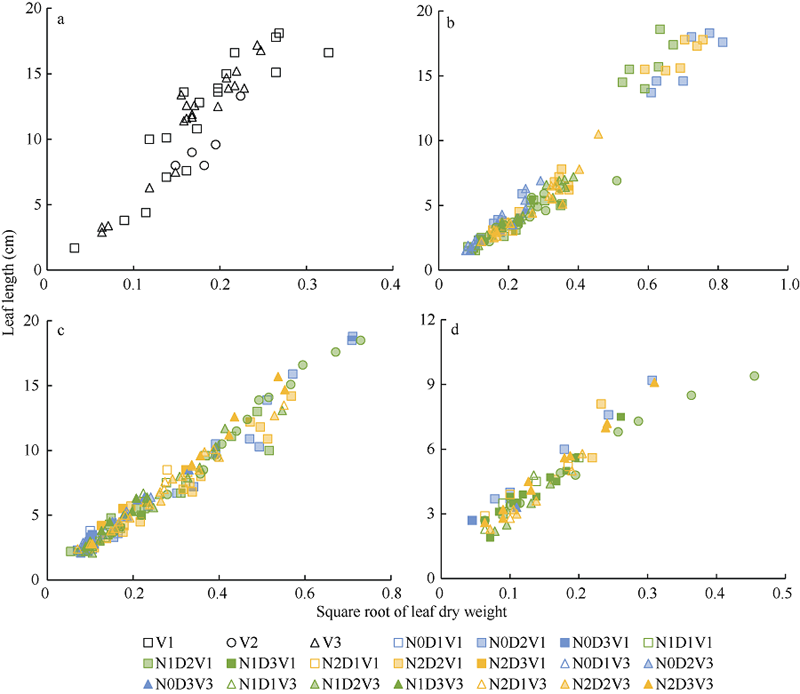 | 图2 2012-2013年不同处理叶长随叶干重平方根的变化规律a、b、c和d分别对应归一化叶位区间(0, 0.185]、(0.185, 0.375]、(0.375, 0.750]和(0.750, 1.000]。Fig. 2 Relationship between leaf length and square root of leaf dry weight for different treatments from 2012 to 2013a, b, c, and d correspond to the interval of normalized leaf ranks (0, 0.185], (0.185, 0.375], (0.375, 0.750], and (0.75, 1.000], respectively. |
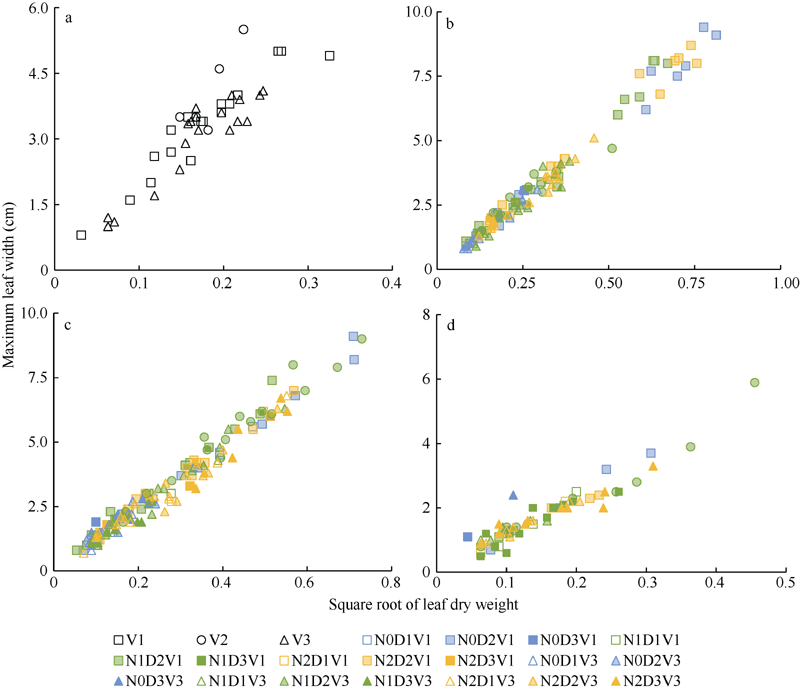 | 图3 2012-2013年不同处理最大叶宽随叶干重平方根的变化规律a、b、c和d分别对应归一化叶位区间(0, 0.185]、(0.185, 0.375]、(0.375, 0.750]和(0.750, 1.000]。Fig. 3 Relationship between maximum leaf width and square root of leaf dry weight for different treatments from 2012 to 2013a, b, c, and d correspond to the interval of normalized leaf ranks (0, 0.185], (0.185, 0.375], (0.375, 0.750], and (0.750, 1.000], respectively. |
| 表3 最大叶宽模型及其参数显著性检验 Table 3 Significance test of maximum leaf width model and its parameter values |
因此, 不同处理下, 油菜出苗后第i天主茎第j叶最大叶宽LWMaxj(i)(cm)可统一表示为
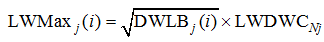
式(8)中, LWDWCNj与LLDWCNj类似, 是归一化叶位为Nj时RLW× SLW× LMC倒数的平方根, 此处命名为叶宽重系数[cm ( 
2.1.3 叶柄长模型 2012— 2013年试验数据表明, 在归一化叶位区间(0, 0.185]和(0.185, 0.375]内, 不同处理油菜叶柄长随叶长的变化呈线性关系(图4), 可用一次函数拟合(统计参数见表4)。
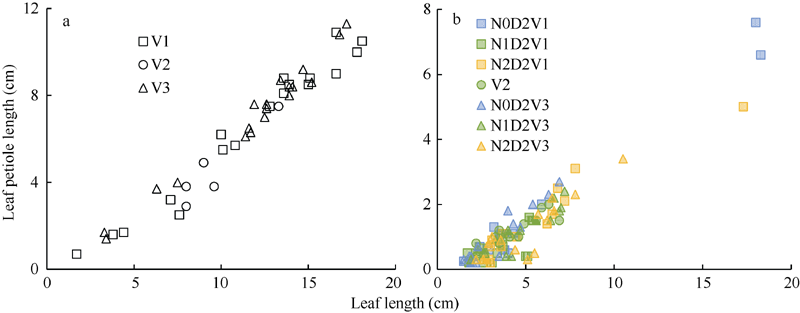 | 图4 2012-2013年不同处理叶柄长随叶长的变化规律a和b分别对应归一化叶长区间(0, 0.185]和(0.185, 0.375]。Fig. 4 Relationship of leaf petiole length with leaf length for different treatments from 2012 to 2013a and b correspond to the interval of normalized leaf ranks (0, 0.185], and (0.185, 0.375], respectively. |
| 表4 叶柄长模型及其参数显著性检验 Table 4 Significance test of petiole length model and its parameter values |
因此, 不同处理下, 油菜出苗后第i天主茎第j叶叶柄长LPLj(i)(cm)可统一表示为
LPLj(i)=LP0+LP1× LLj(i) (9)
式(9)中, LP0与LP1是模型参数, 其取值及显著性测验见表4。
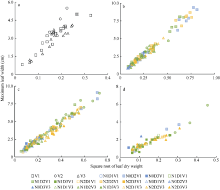
利用试验I的实测值和模拟值以1:1绘图(图5、图6和图7), 统计检验(表5)表明, 各品种叶长、最大叶宽和叶柄长实测值与模拟值的相关系数(r)均达P< 0.001显著水平。除V2叶柄长的da略大于1 cm、dap为18.472%外, 其余da的绝对值均在1 cm以内、dap均在10%以内, 说明除V2叶柄长模型还需要进一步试验数据测试与改进外, 上述模型总体模拟效果较好。
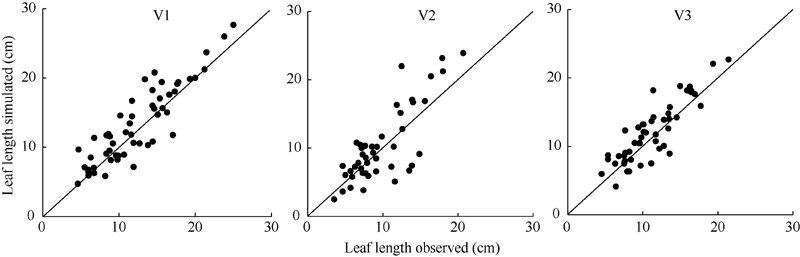 | 图5 2011-2012年3个品种叶长实测值与模拟值比较Fig. 5 Comparison of observation and simulation of leaf length for three cultivars from 2011 to 2012 |
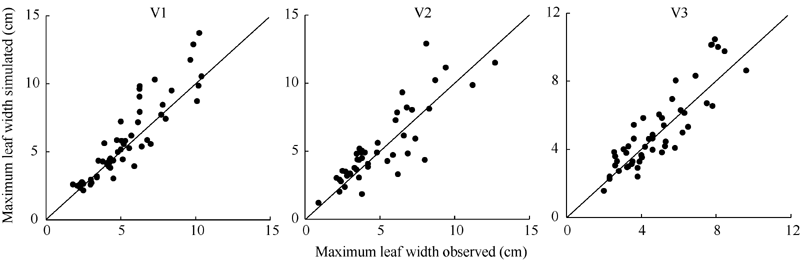 | 图6 2011-2012年3个品种最大叶宽实测值与模拟值比较Fig. 6 Comparison of observation and simulation of maximum leaf width for three cultivars from 2011 to 2012 |
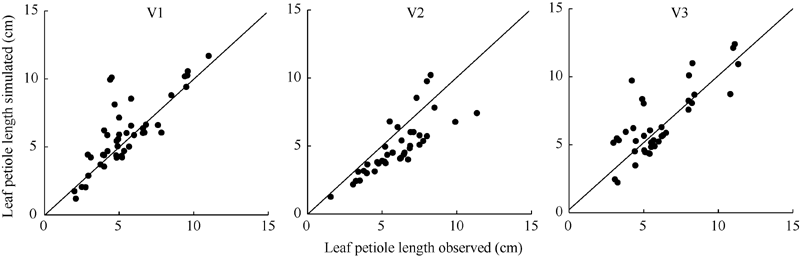 | 图7 2011-2012年3个品种叶柄长实测值与模拟值比较Fig. 7 Comparison of observation and simulation of leaf petiole length for three cultivars from 2011 to 2012 |
| 表5 2011-2012年油菜主茎叶片形态参数模型观察值与模拟值比较的统计参数 Table 5 Comparison of simulation and observation of rapeseed main stem leaf morphological parameter models from 2011 to 2012 |
功能-结构植物模型将作物生长模型之功能与作物形态模型之结构有机结合[5], 提高了模型的机理性, 加强了品种对环境因子的响应。Groer等[30]和Jullien等[33]都将叶面积与叶片生物量联系起来, 嵌入GREENLAB模型的源-库关系, 具有较强的机理性, 但前者仅考虑了氮素的影响, 且不能从出苗开始模拟, 后者则更侧重于分枝的描述, 两者都未模拟叶宽和叶柄长等具体形态参数。本研究根据比叶重等常用参数的计算方法, 得出叶长和叶宽与叶片干重的定量关系, 并构建了基于生物量的叶长、叶宽模型。叶片干重可由已有的、能响应不同品种、环境及栽培管理措施的油菜生长模型[25]获得, 所建模型理论上也可将各种因素(包括环境条件和栽培管理措施等)的影响通过叶片生物量传递给叶形, 这种传递性使模型表达更为简明。因此, 本研究将随各种因素而变化的叶形系数、叶片长宽比和比叶重等的复杂关系, 统一为具有明确生物学意义的叶长重系数和叶宽重系数2个整合参数, 在不失机理的前提下简化了模型。基于不同品种和施肥水平的独立试验数据检验所建模型, 结果表明模型观察值与模拟值一致性较好, 说明所建模型具有一定精度, 且机理性较强, 能适应不同年份环境条件。
为了统一不同处理叶位的差异, 本研究对叶位进行了归一化处理, 并根据油菜叶形等因素将归一化叶位分为(0, 0.185]、(0.185, 0.375]、(0.375, 0.750]和(0.750, 1.000] 4个区间。按叶片定长时间, 这4个区间大致分别对应于移栽前、移栽后苗期、蕾薹期和开花期及以后; 按叶位, 则较为严格地对应于第1~第4叶、第5叶及其后的长柄叶、短柄叶和无柄叶。曹宏鑫等[35]和张伟欣[36]建立了基于生物量的油菜叶形模型, 但由于时效和数据所限, 他们构建的叶形模型所对应的叶位, 大致属于本研究前两个归一化叶位区间, 且建模时未涉及后期的数据。因此, 本研究所建模型较前人研究具有更强的普适性。但本文用于建模和检验的生物量数据均为单叶实测数据, 限于篇幅, 生长模型干物质分配部分将另文详述。
应当指出, 本研究仍存在一些不足。较之曹宏鑫等[35]和张伟欣[36]的研究, 本文仅考虑了生物量对叶片形态参数的影响, 而未考虑其对其他叶形参数及其他器官的影响; 较之Jullien等[33]实现的较为完整的功能-结构植物模型, 本研究仅停留在功能对结构的影响, 而尚未涉及结构对功能的反馈及其相互影响。这些都有待今后进一步研究。
基于2011— 2012年和2012— 2013年不同品种、移栽密度和施肥水平油菜田间试验, 构建了基于生物量的油菜主茎叶长、最大叶宽和叶柄长模型。提出了具有明确生物学意义的叶长重系数和叶宽重系数概念, 除宁油16叶柄长模型误差较大外, 模型总体预测性较好, 且具有一定机理性和普适性。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|