{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
谷子转录因子基因 SiNAC45在拟南芥中对低钾及ABA的响应
[王二辉1, 2, *  , 胡利芹
, 胡利芹2, * , 薛飞洋1, 2 , 李微微2 , 徐兆师2 , 李连城2 , 周永斌2 , 马有志2 , 刁现民2 , 贾冠清2 , 陈明2, * , 闵东红1, * ]
, 胡利芹, 闵东红]
|
|


第一作者联系方式: E-mail:wangerhui191132@163.com, Tel: 13366320737 **同等贡献(Contributed equally to this work)
NAC (nascent polypeptide-associated complex)转录因子在植物生长发育和非生物胁迫响应等过程中发挥重要的调控作用, 目前, 关于NAC转录因子参与耐低钾胁迫的研究报道很少。本研究在前期工作对低钾胁迫的转录组测序基础上, 对筛选出的一个低钾胁迫下表达量上调的NAC类转录因子基因 SiNAC45进行了深入研究。结果表明, SiNAC45基因全长1383 bp, 编码461个氨基酸, 分子量为50.7 kD, 等电点为6.92。 SiNAC45在20~100个氨基酸之间有一段NAM保守结构域。系统进化树结果显示, SiNAC45位于NAC基因家族的第1亚族。基因表达谱分析显示, SiNAC45主要在谷子根部表达, 并且能够被低钾和ABA诱导表达。亚细胞定位结果显示SiNAC45定位于细胞核。基因功能分析结果显示, 在不同浓度低钾处理下, 和野生型拟南芥相比, SiNAC45转基因拟南芥的根长和植株鲜重显著增加, 说明过表达 SiNAC45可以提高转基因植物对低钾胁迫的抗性。下游基因表达分析结果显示, 在 SiNAC45转基因植物中两种重要的钾离子转运体基因 AKT1和 HAK1的表达显著提高, 证明 SiNAC45通过调控植物钾离子转运体基因的表达影响植物对低钾胁迫的耐性。种子萌发试验结果显示, SiNAC45转基因拟南芥与野生型拟南芥相比对ABA的敏感性降低, 说明 SiNAC45可能负调ABA信号。
, HU Li-Qin, MIN Dong-Hong
NAC (nascent polypeptide-associated complex) like transcription factors play important role in plant growth and development, abiotic stress response, and other processes. Currently, few researches reported NAC like transcription factors involving in tolerance to low potassium stress. In this study, we found and researched a NAC like transcription factor gene SiNAC45 on the basis of transcriptome sequence of millet under low potassium stress which had been completed in previous work. The result show that the full-length of SiNAC45 is 1383 bp, encoding 461 amino acids, with molecular weight and isoelectric point of 50.7 kD and 6.92, respectively. There is a conserved NAM domain between 20-100 amino acids of SiNAC45. The phylogenetic tree showed that SiNAC45 belonged to the first subfamily of NAC gene family. The gene expression profile results indicated SiNAC45 mainly expressed in roots and was induced by ABA and low potassium treatment. The protein subcellular localization results of SiNAC45 revealed that it was localized in the nucleus. Gene functional analysis showed that under treatment with different concentrations of potassium, root length and fresh weight of SiNAC45 transgenic Arabidopsis significantly increased compared with those of wild-type Arabidopsis, and there was no significant difference in the number of lateral roots between transgenic and wild-type Arabidopsis, indicating that overexpressing of SiNAC45 in transgenic plants can enhance tolerance to low potassium stress. Expression analysis of downstream gene showed that expression of two important potassium transporter genes AKT1 and HAK1 increased significantly in SiNAC45 transgenic plants, indicating that SiNAC45 affects the tolerance to low potassium stress of plants by regulating the expression of potassium transporter gene. Seed germination test results showed that Arabidopsis carrying SiNAC45 decreased the sensitivity to ABA compared with wild-type Arabidopsis, indicating that SiNAC45 maybe negatively regulate ABA signal pathway.
NAC是一类植物中的重要转录因子基因家族[1, 2], 首次报道的NAC转录因子与矮牵牛顶端分生组织和胚胎形成有关[3], 随后在不同植物中报道了越来越多的NAC转录因子, 到目前为止, 已经发现100多种NAC转录因子, 除拟南芥和水稻外, 在大豆[4]、马铃薯[5]、柑橘[6]、谷子[7]、苹果[8]、葡萄[9]等其他植物中也发现NAC转录因子。但是在动物中还未发现NAC家族基因[10]。在拟南芥和水稻中, NAC基因家族有一百多个成员[11], 但是这些成员中功能得到证明的却很少。NAC家族基因在植物的发育、形态建成、次级细胞壁、次生根、胚胎和花的形成以及抵抗非生物胁迫过程中具有很重要的作用。NAC家族蛋白的共同特征是在N端有一个高度保守的NAM结构域, 而在C端有一个序列和长度多样化的转录激活域[12], 但是有些NAC家族的基因没有这种特殊的结构域(NAM, CUC)[13]。NAC家族转录因子能够通过这种结构域特异地结合顺式作用元件CGT(G/A)和CACG来调节下游靶基因的表达[14, 15]。NAC基因家族可以分为5个亚家族[16], 其中第3亚家族成员在非生物胁迫响应过程中发挥重要作用[17, 18]。虽然对于NAC转录因子基因在生物胁迫和非生物胁迫响应中的作用有很多报道, 但是到目前为止少见对其参与植物耐低钾胁迫的报道。另外, 有些NAC家族成员被报道参与植物对ABA的反应, 例如拟南芥的ANAC019和ANAC055基因, 对于其参与ABA信号途径的分子机制仍然未知[19]。
钾元素在植物生长发育和代谢过程中有很重要的作用, 植物根系从土壤中吸收钾来维持正常的生命活动。当土壤中缺乏钾营养时会激活植物体内两种重要的营养吸收机制, 一种是植物体内营养的储存和分配机制[20], 比如转运蛋白和转运通道。另一种是植物根系生长发育进程的改变, 包括主根生长、侧根形成、根毛伸长[21]。文献报道, 植物体内缺乏K+时会激活K+转运蛋白来促进植物根系对K+的吸收[20]。拟南芥中的K+转运体有整流K+通道AKT1和低钾诱导K+转运体HAK5两种, 它们主要在低钾胁迫时促进植物对K+的吸收[22]。在拟南芥和其他物种中HAK5在低钾胁迫下能够被快速激活响应钾胁迫[23]。拟南芥根系吸收K+有高亲和K+吸收和与低亲和K+吸收2种方式, KUP/HAK/KT家族的转运体为高亲和性K+转运体[24], AKT1可以与其亚族中的成员形成异源多聚体来调节植物根系对K+的吸收。KAT1主要参与植物叶片气孔打开时对K+的吸收[25], GORK主要控制叶片气孔关闭时K+的流出[26]。KUP/HAK/KT可能对维持植物体内K+平衡有重要作用。KUP4/TINY ROOT HAIR1(TRH1) 突变后会损害拟南芥根毛的伸长[27]。有些转录因子可以在低钾胁迫下激活钾离子转运体和钾离子运输通道基因转录, 例如AtMYB35在低钾胁迫下可以激活KPU转运体促进植物对钾的吸收[27]。
谷子具有抗旱、耐贫瘠等特点, 是作物抗逆研究的理想材料。目前植物中关于NAC类转录因子的研究主要集中在拟南芥、水稻等模式植物中, 关于谷子NAC类转录因子参与植物逆境胁迫应答的分子机制尚缺乏了解。为揭示谷子中NAC类转录因子的特性及功能, 本研究分析低钾胁迫条件下表达量逐渐上升的SiNAC45基因的特性及生物学功能, 旨在为研究谷子抗逆分子机制提供证据, 也为作物耐低钾遗传改良提供新的基因资源。
1.1.1 植物材料 拟南芥野生型(Columbia生态型, WT)由本实验室保存, 谷子品种H214由中国农业科学院作物科学研究所刁现民课题组提供。
1.1.2 载体和菌株 大肠杆菌、农杆菌GV3101、pBI121载体、GFP载体的质粒都由本实验室保存, pZeroBack载体购于北京天根公司。
1.1.3 试验试剂 限制性内切酶、T4 DNA连接酶购于Promega公司; 质粒提取试剂盒、DNA凝胶回收试剂盒、RT-PCR试剂盒购于天根公司; 由奥科生物技术科技有限公司合成引物和测序; 其他化学药品为国产分析纯试剂。
谷子数据来源于Phytozome (http://www.phytozome. net/search.php), 根据谷子SiNAC45蛋白序列在Phytozome数据库中进行同源性搜索, 下载与谷子SiNAC45蛋白序列相似度较高的其他物种的蛋白序列。用MEGA5软件对谷子SiNAC45蛋白序列及其同源蛋白序列进行多序列比对分析, 用邻接法生成系统进化树, 设BootStrap值为1000。利用MEME (Multiple Em for Motif Elicitation)在线工具分析蛋白序列。利用SMART (http://smart.embl-heidelberg.de/)在线工具分析谷子SiNAC45蛋白的结构域。
从谷子基因组数据库Phytozome (http://phytozome. jgi.doe.gov/pz/portal.html)搜索SiNAC45序列并截取起始密码子ATG上游2000 bp作为SiNAC45启动子。用PLACE (http://www.dna.affrc.go.jp/PLACE/)数据库对SiNAC45启动子中的顺式作用元件进行分析。
根据谷子基因SiNAC45的CDS序列设计基因引物45-F和45-R (表1), 用TRIzol试剂盒(TIANGEN, 北京)提取谷子H214植株总RNA, 用Primer Script反转录试剂盒(TaKaRa)反转录成cDNA, 以cDNA为模板扩增SiNAC45, 并将其回收纯化后连接到pZeroBack载体上。以pZeroBack-SiNAC45质粒为模板扩增SiNAC45, 引物为45-F3和45-R3。采用In-Fusion试剂盒(TaKaRa)将SiNAC45从连接的pZeroBack载体上扩增并插入带有CaMV35S启动子的pBI121表达载体, 采用农杆菌转化法[28]转化拟南芥。
谷子幼苗在营养土中正常生长(温度22℃、相对湿度65%、光照周期16 h/8 h) 3周, 然后将幼苗分别移至低钾(20 μ mol L-1)、干旱(6% PEG)、ABA (100 μ mol L-1)、NaCl (100 mmol L-1)、低氮(0.2 mmol L-1氮)、低磷(5 μ mol L-1)的水培营养液中胁迫处理, 并于处理后的0、1、3、6、12、24和48 h分别取样, 用TRIzol (TianGen, 北京)提取谷子植株总RNA; 将低钾(20 μ mol L-1)处理的部分幼苗分成根、茎、叶, 用TRIzol (TIANGEN, 北京)提取其RNA; 分别取低钾(MS+20 μ mol L-1)处理转基因和野生型拟南芥植株, 提取总RNA检测下游基因。分别用6种胁迫下的谷子总RNA、低钾处理下的谷子根、茎、叶RNA和低钾处理下的转基因和野生型拟南芥RNA反转录产物作为模板, 以SYBR Green染料法, 在ABI Prism 7500上进行实时荧光定量PCR。RT-PCR体系含2× SuperReal PreMix Plus (含荧光染料)(天根公司) 12.5 μ L、10 μ mol L-1正向引物和反向引物各0.5 μ L、50× ROX Reference Dye∆ 0.5 μ L、RNase-free ddH2O 9.5 μ L。反应条件为95℃预变性10 min; 95℃变性15 s, 60℃退火20 s, 72℃延伸30 s并收集荧光信号, 35个循环, 用2-∆ ∆ Ct法计算该基因表达量。RT-PCR引物序列为45-F4和45-R4 (表1), 用谷子Actin基因(Si001873m.g)作为内参基因, 其引物为SiActin-F和SiActin-R; 下游基因检测引物为AKT1-F和AKT1-R、HAK-F和HAK-R, 内参基因(AT3G15260)引物为AtActin- F和AtActin-R (表1)。
| 表1 SiNAC45基因克隆和Real-time PCR分析所用引物以及引物退火温度 Table 1 Primers used for SiNAC45 gene cloning and Real-time PCR analysis and annealing temperature of the primers |
扩增SiNAC45基因的编码区全长, 引物为45-F1和45-R1 (表1), Bam H I酶切GFP表达载体, 利用In-fusion技术构建phGFP-SiNAC45载体。参考Yoo等[29]的方法制备拟南芥原生质体, 将融合表达的重组质粒phGFP- SiNAC45和作为对照的GFP空载体质粒分别转化制好的原生质体, 黑暗培养18 h以上, 并在激光共聚焦显微镜下观察结果。
野生型和转基因拟南芥的种子经70%酒精处理3 min, 无菌水洗3次, 每次1 min左右; 用0.5%~0.8%的次氯酸钠处理10~15 min, 无菌水清洗3次, 每次1 min; 4℃春化2~3 d, 然后将部分种子分别移到MS、MS+1 μ mol L-1ABA、MS+2 μ mol L-1 ABA和MS+4 μ mol L-1 ABA的培养基中培养(温度22℃、相对湿度65%、光照周期16 h/8 h)一周, 统计种子萌发率; 其余部分种子分别移到MS培养基培养4~5 d, 再移至MS、MS+5 μ mol L-1K+、MS+10 μ mol L-1K+培养基上生长一周, 分别测量不同处理下过表达株系和野生型拟南芥的根长和植株鲜重, 使用根系扫描仪扫描根表面积, 运用方差分析软件分析统计结果。
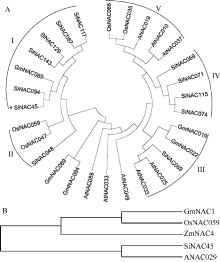
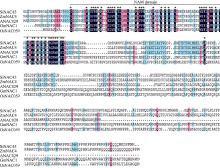
本研究前期对谷子低钾胁迫转录组测序, 从中发现一个低钾诱导下表达上调的NAC类转录因子SiNAC45。在谷子基因组数据库(http://www.phytozome.net/)搜索SiNAC45全长序列, 发现SiNAC45基因全长1383 bp, 有3个外显子, 4个内含子, 编码461个氨基酸, 分子量为50.7 kD。运用MEGA5软件对SiNAC45蛋白及其他物种中的同源蛋白做了系统进化树分析(图1-A, 图1-B), NAC转录因子共分为5个亚家族, 其中I亚族成员最多。SiNAC45位于第I亚族, 与谷子中的SiNAC094进化关系最近。SiNAC45与其他植物中的NAC基因的蛋白序列比对结果(图2)显示, SiNAC45与来自水稻、玉米、大豆等植物的NAC蛋白序列都包含一个NAM保守域, SiNAC45与ANAC029同源性最高, 达到59%。
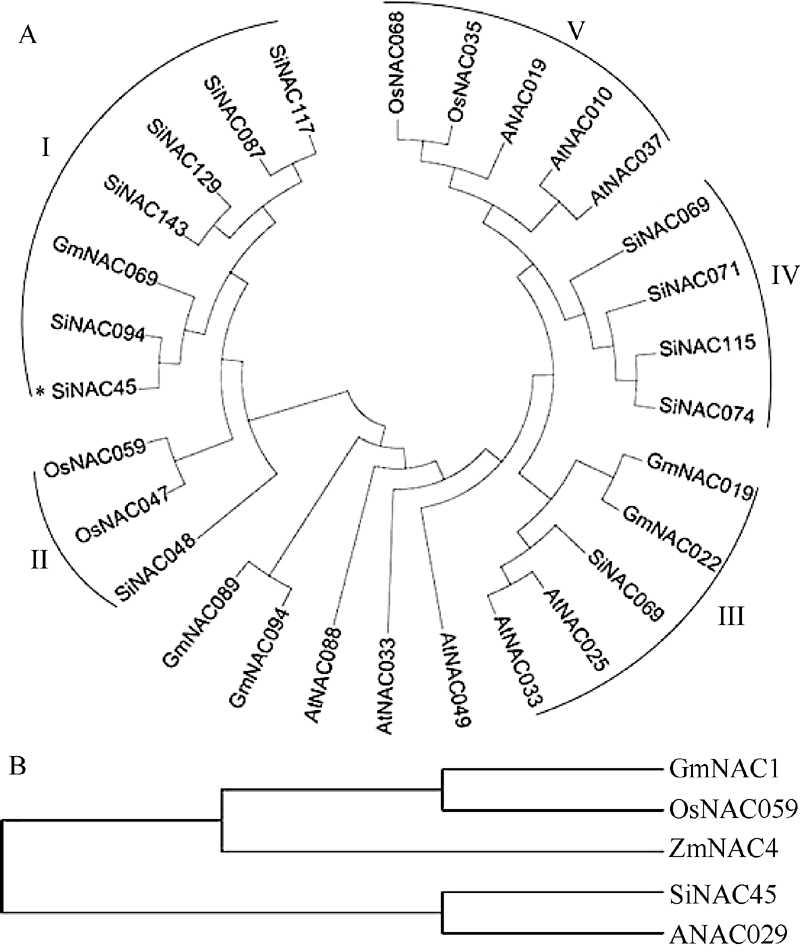 | 图1 谷子与拟南芥、水稻、大豆、玉米NAC蛋白进化树分析结果所示物种的NAC蛋白质序列从数据库Phytozome v10.0.4中获得。Fig. 1 Phylogenetic analysis of NACs protein fromSeteria italica(Si), Oryza sativa (Os), Arabidopsis thaliana (At), Glycine max (Gm), and Zea mays (Zm)The NAC protein sequences in those plants were obtained from the database Phytozome v10.0.4. |
 | 图2 部分物种中NAC家族基因的进化关系Fig. 2 Evolutionary relationship of some NAC family genes |
用PLACE (http://www.dna.affrc.go.jp/PLACE/)在线工具分析SiNAC45上游启动子区的顺式作用元件(表2), 发现SiNAC45启动子区域中包含WARK、W-box (wound responsive element)、LTRE (cold responsive element)、MYB (drought responsive element)、ABRE (ABA responsive element)、MYC (ABA and cold responsive element)、I-box (light responsive element)等逆境相关顺式作用元件, 其中MYC元件最多, 为32个, 其次为W-box, 共24个, MYB元件为16个。据报道, 这些元件在植物干旱、ABA、低温等胁迫响应中发挥作用。
| 表2 SiNAC45启动子逆境相关顺式作用元件分析 Table 2 Analysis of cis-elements in the SiNAC45 promoter |
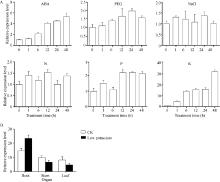
通过荧光定量PCR检测低氮(0.05 mmol L-1)、低钾(0.5 μ mol L-1)、低磷(0.5 μ mol L-1)、干旱、脱落酸(ABA)、NaCl (50 mmol L-1)等不同胁迫处理下的谷子幼苗植株中SiNAC45的表达量(图3-A)表明, 在ABA处理下, SiNAC45的表达量逐渐上升并在48 h时达到峰值, 表达量是处理前的6倍; 在低钾处理下, SiNAC45的表达量逐渐上升, 在48 h时达到最高值, 表达量提高了35倍; 在PEG、NaCl、低氮和低磷处理下, SiNAC45的表达量提高并不显著, 表明SiNAC45对低钾胁迫和ABA有明显响应。
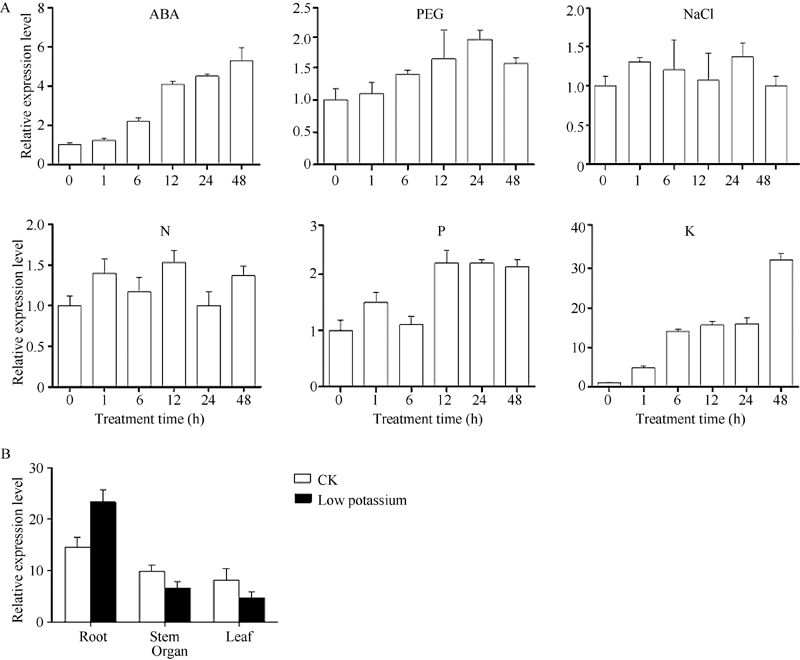 | 图 3 SiNAC45在不同处理下的表达模式A: 不同处理下SiNAC45的表达模式; B: 正常生长和低 K处理条件下SiNAC45在谷子不同组织中的表达。Fig. 3 Expression patterns of the SiNAC45 gene under various treatmentsA: expression pattern of SiNAC045 under different treatments; B: tissues specific expression of SiNAC45 under normal condition and low potassium treatment condition. |
SiNAC45在谷子组织表达特异性分析结果(图3-B)显示, 在正常生长和低钾处理条件下的谷子根、茎和叶均有SiNAC45的表达, 在正常条件下SiNAC45在根部的表达量最高, 在低钾处理条件下, SiNAC45在根部的表达量相对增加(图3-B), 证明SiNAC45主要是在根部响应低钾胁迫。
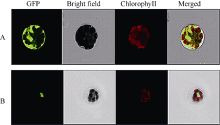
将SiNAC45与GFP蛋白融合的SiNAC45-hGFP融合表达载体和GFP空载体分别转入拟南芥原生质体中, 激光共聚焦显微镜下观察结果显示, 转入对照GFP空载体的GFP蛋白在细胞核、细胞质、细胞膜中均有表达; 而转入SiNAC45-GFP融合表达载体的原生质体只在细胞核中能够发现绿色荧光信号, 表明SiNAC45定位在细胞核中(图4)。
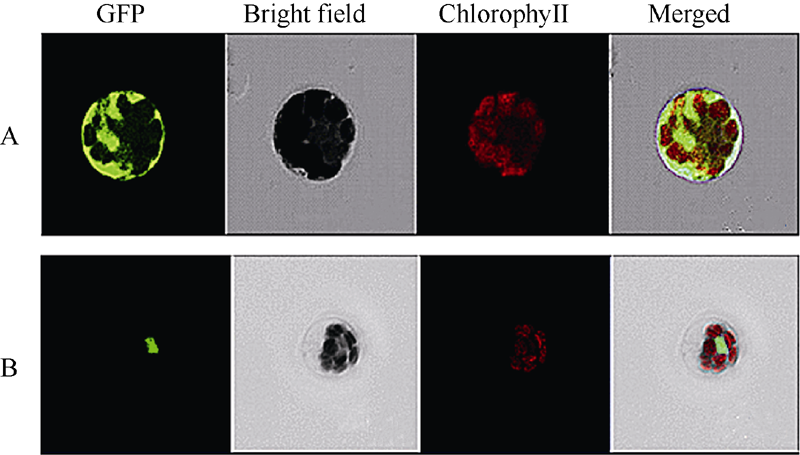 | 图4 SiNAC45蛋白的亚细胞定位分析结果A: 对照GFP定位于整个细胞; B: SiNAC45-GFP定位于细胞核中。Fig. 4 Subcellular localization of SiNAC45 proteinA: control GFP; B: SiNAC45-GFP localized in the nuclear. |
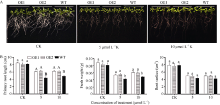
通过在MS培养基中分别添加5 μ mol L-1 K+和10 μ mol L-1 K+, 对2个转SiNAC45基因拟南芥株系和野生型拟南芥进行2种浓度的低钾处理, 结果显示在MS培养基上过表达SiNAC45的转基因拟南芥的根长、植株鲜重和根表面积与野生型拟南芥没有明显差别(图5-A), 而在5 μ mol L-1 K+和10 μ mol L-1 K+的低钾胁迫处理下, 与野生型拟南芥相比, 过表达SiNAC45的转基因拟南芥的根表面积增大(图5-A)但并不明显, 根长增长, 差异达到极显著水平(图5-B), 植株鲜重明显提高, 差异达到显著水平(图5-B)。以上结果显示SiNAC45转基因拟南芥低钾胁迫的抵抗力显著强于野生型拟南芥。
 | 图5 SiNAC45转基因和野生型拟南芥的表型分析A: 对照和低钾处理下的表型, 低钾浓度分别为5 μ mol L-1和10 μ mol L-1。B: 正常条件和低钾处理下的主根长、鲜重和根表面积; 采用单因素方差分析法对数据统计分析, 柱上不同的小写字母代表柱值在0.05水平上差异显著, 不同大写字母代表柱值在0.01水平上差异显著。Fig. 5 Phenotype analysis ofSiNAC45 transgenic and wild-type Arabidopsis to low potassium stressA: phenotype of control and low potassium treatment with 5 μ mol L-1 and 10 μ mol L-1K. B: primary root length, fresh weight and root surface under low potassium treatment. Data statically analysis was made by the means of one-way ANOVA. The values marked with different lower-case letters on the columns are significantly different at the 0.05 probability level; those with different capital letters are significantly different at the 0.01 probability level. |
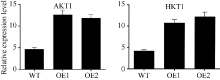
为了研究SiNAC45耐低钾胁迫的机制, 检测了在低钾处理下SiNAC45下游与钾离子吸收有关基因的表达。结果(图6)显示, 在低钾条件下, 在SiNAC45转基因拟南芥中钾离子转运体基因AKT1和HAK1的表达量都明显高于野生型拟南芥, 说明SiNAC45过表达后通过影响AKT1和HAK1的表达来提高植物对低钾胁迫的耐性。
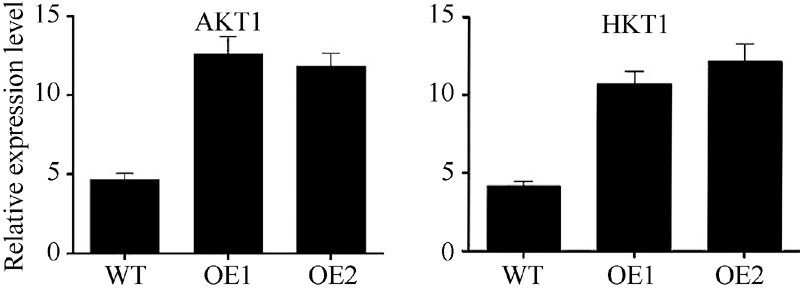 | 图6 低钾处理下转SiNAC45基因和野生型拟南芥中与钾相关下游基因检测Fig. 6 Testing of downstream gene related to potassium inSiNAC45 overexpression and wide-type Arabidopsis |
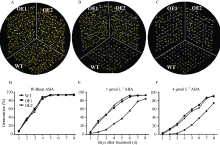
将野生型和过量表达SiNAC45转基因拟南芥种子分别播种于MS、MS+1 μ mol L-1ABA、MS+4 μ mol L-1ABA培养基中的种子萌发试验显示, 在MS培养基中过量表达SiNAC45转基因拟南芥和野生型拟南芥种子萌发速率基本保持一致, 在第4天以后萌发率维持在95%左右(图7-A, D); 在含1 μ mol L-1ABA的培养基中SiNAC45转基因拟南芥的萌发速率明显高于野生型拟南芥(图6-B), 转基因拟南芥在第6天萌发率达到96%, 野生型拟南芥在第8天达到85% (图7-E); 在4 μ mol L-1ABA处理时野生型拟南芥的萌发速率明显比转基因拟南芥慢, 直到第3天才开始萌发, 到第8天达到最高值71%, 而转基因拟南芥的萌发率最高为93% (图7-C, F)。在ABA处理下野生型拟南芥种子的萌发率明显低于SiNAC45转基因植物, 说明SiNAC45过表达降低了对ABA的敏感性, SiNAC45负调植物ABA信号途径。
 | 图7 35S::SiNAC45过表达拟南芥种子在不同ABA浓度处理下的萌发率A和D: 种子在无ABA处理条件下的萌发率; B和E: 种子在1 μ mol L-1 ABA处理条件下的萌发率; C和F: 种子在4 μ mol L-1 ABA处理条件下的萌发率。Fig. 7 Seed germination rates of 35S::SiNAC45 transgenic lines under treatments with different concentrations of ABAA and D: seed germination rates under the condition without ABA; B and E: seed germination rates under the condition with 1 μ mol L-1 ABA; C and F: seed germination rates under the condition with 4 μ mol L-1 ABA. |
NAC转录因子家族是植物中最庞大的家族之一, 其家族成员的数量在不同物种中差异较大。许多NAC家族的成员在作物胁迫响应中发挥重要的作用[37, 38]。到目前为止, 在水稻和拟南芥中发现了140多种NAC或NAC类似基因, 被分为5个亚族[39], 所有已知与胁迫相关的NAC类基因被分在第III亚族[40]。例如, 属于NAC第三亚族的SNAC1基因过表达后能提高植物的抗旱性, OsNAC6过表达能提高植物抗旱性和耐盐性[41]。其他亚族与非生物胁迫相关的NAC基因还未见报道。NAC第I亚族的基因大多与植物发育相关[42]。本研表明SiNAC45属于第I亚族(图1-A), 可被多种非生物胁迫诱导表达, SiNAC45在低钾胁迫处理下表达很高(图2-A), 在处理48 h时SiNAC45的表达量提高了大约35倍 (图2-A), 在低钾处理条件下SiNAC45转基因拟南芥的根系明显比野生型拟南芥发达, 鲜重显著大于野生型拟南芥, 表明SiNAC45过表达后提高了植物对低钾胁迫的耐性(图4)。这是第一次报道NAC类基因家族成员参与植物低钾胁迫的响应, 正向调控植物对低钾胁迫的耐性。另一方面, 一些NAC家族的基因包括OsNAC6/SNAC2[43, 44]和其他家族基因OsDREB1A、OsDREB1B、AtDREB1A、AtDREB1B[45]过表达后能够在正常条件下延缓植物的生长, 这一表型限制了这些基因在生产中被应用, 而SiNAC45转基因拟南芥在正常条件下和野生型拟南芥的生长一致, 并没有延缓拟南芥的生长。因此, SiNAC45基因克隆为作物钾高效利用遗传改良提供了新的候选基因。到目前为止, 在拟南芥中已经发现71种钾离子通道和钾离子转运体, 它们被分为6个基因家族, 包括3个钾离子通道家族(Shaker、TPK和Kir-like家族)和3个转运体家族(KUP/HAK/KT、HKT和CPA家族)[46]。AKT1被认为是Shaker家族最重要的钾离子运输通道, 它在钾离子吸收过程中首先在拟南芥根表皮细胞中表达, 进而在钾离子从土壤到根表皮细胞的运输中发挥重要功能[46]。同时, 还有一些K+转运体基因能够在低钾条件下被诱导表达, 例如AtHAK5、AtKEA5、AtKUP3、AtCHX13、AtCHX17[47]。在本研究中, 在低钾条件下, SiNAC45转基因拟南芥中HAK1和AKT1的表达量明显高于野生型拟南芥, 表明SiNAC45通过提高HAK1和AKT1在植物根部的表达来促进植物对钾离子的吸收, 从而在低钾胁迫下提高SiNAC45转基因植株对低钾的耐性。
据报道, 大多数ABA响应基因的启动子区都包含ABRE元件[48]。OsNAC5和OsNAC6启动子区域包含ABA响应元件ABRE (ACGTG/TC), OsNAC5和OsNAC6可以通过ABRE响应ABA[48]。启动子分析结果表明, SiNAC45的启动子区包含7个ABRE元件(表2), 同时, SiNAC45能够被ABA诱导表达, 在48 h时表达量提高了大约6倍(图2-A), 说明SiNAC45也是通过ABRE元件响应ABA处理的(图6)。基因功能分析结果显示, SiNAC45在种子萌发期降低了植物对ABA的敏感性, 而在幼苗期却没有表现这一特点(图6), 说明SiNAC45基因可能在种子萌发期负向调控植物ABA信号途径。关于SiNAC45提高转基因植物对低钾胁迫的耐性是否与该基因对ABA的响应相关还有待进一步分析。本研究初步阐明了谷子SiNAC45在低钾胁迫响应过程中发挥重要作用, 同时负调植物对ABA的反应, 这些研究结果为我们进一步了解谷子的抗逆机制提供了新的证据。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|