
{kind=link}
{kind=link}
结薯数差异显著的甘薯品种生长前期根系特性及根叶糖组分比较
[王翠娟1  , 史春余
, 史春余1, * , 刘娜2 , 刘双荣1 , 余新地1 ]
, 史春余, 刘娜|
|


* 通讯作者(Corresponding author): 史春余, E-mail: scyu@sdau.edu.cn, Tel: 0538-8246259 第一作者联系方式: E-mail: cuijuanwangwang@126.com
选用结薯数差异显著的甘薯品种商薯19和济徐23, 于2013年和2014年分别在大田和盆栽条件下调查生长前期根系生长发育情况, 测定根叶糖组分, 分析其变化规律及其与单株有效薯块数形成的关系。结果表明, 与济徐23相比较, 商薯19发根缓苗迅速, 整个生长前期均较为侧重吸收根系建成, 且秧苗栽后15 d和30 d地上部和整个根系生物量比值(T/TR)相似; 而济徐23较早完成不定根的发生(秧苗栽后15 d)和不定根向块根的分化建成(秧苗栽后30 d), 封垄期(秧苗栽后45 d)具有较小的地上部和块根生物量比值(T/SR)。2个品种在生长前期根叶中蔗糖、己糖代谢和贮藏糖类多聚物的形成具有显著差异, 其中商薯19整个生长前期比济徐23的根系蔗糖/己糖比率显著低, 在块根分化建成过程中根叶间具有较高的蔗糖浓度梯度, 且根系中存在2种低果聚糖(蔗果三糖和蔗果四糖), 秧苗栽后30、45 d叶片淀粉含量显著低; 而济徐23在块根分化建成过程中根系仅有蔗果四糖, 且与商薯19比, 封垄期根系蔗糖/可溶性总糖比率相似而根系蔗糖/淀粉比率显著低。2013年和2014年的大田试验均表明, 封垄期商薯19具有显著多的单株有效薯块数和显著高的单株有效薯块鲜重( F=10.71, P=0.0170; F=13.97, P=0.0212), 而收获时商薯19具有显著多的单株有效薯块数和显著高的块根产量( F=353.89, P<0.0001; F=88.94, P<0.0001), 济徐23具有显著高的平均单薯鲜重( F=10.32, P=0.0124)。
Starch sweet potato varieties Shangshu 19 and Jixu 23 differing in valid storage root number per plant significantly were used to investigate root characteristics, sugar components in root and leaf at early growth phase and their relationship with the formation of storage root per plant at top cover stage. The results showed that, Shangshu 19 with higher valid storage root number regrew quickly with more new roots after seedling transplanting, developed fiberous roots mainly in the whole early growth phase, and formed a stable rate of top biomass/total root system biomass (T/TR) at 15 and 30 days after planting. Jixu 23 achieved the steady number of adventitious root and valid storage root at 15 and 30 days after planting, respectively. Meanwhile, Jixu 23 showed the lower rate of top biomass/storage root biomass (T/SR) at top cover stage (45 days after planting). On the other hand, in point of metabolism of sucrose and hexoses, and the formation of stored polysaccharide polymers, Shangshu 19 had significantly lower rate of sucrose/hexoses in the whole early growth phase, formed the greater sucrose concentration gradient between leaves and roots, and had 1-kestose and nystose in roots at 15 and 30 days after planting. Its starch content in leaf was significantly lower than that of Jixu 23 at 30 and 45 days after planting. Meanwhile, Jixu 23 only had Nystose in roots before storage root formation, with significantly lowest rate of sucrose/starch when the rate of sucrose/total soluble sugar was similar to that of Shangshu 19 at 45 days after planting. A two-year field trials (2013-2014) were performed to investigate yield-contributing traits and the fresh storage root yield at top cover stage and harvest period, in which, Shangshu19 showed the more valid storage roots per plant, higher valid storage root fresh weight per plant or storage root at top cover stage and harvest period, meanwhile, Jixu23 had significantly higher average fresh weight per storage root.
作物产量是生长发育、器官建成、物质生产积累过程的最终结果[1, 2], 甘薯在生长中后期(封垄期至收获期)茎叶生长过旺与块根膨大缓慢, 物质生产分配的不平衡限制了块根产量的进一步提高。目前针对该限制的研究主要集中于在甘薯生长中后期块根膨大时促进物质生产积累, 并更大程度地向块根运转的调控措施及产量形成机理, 例如增施钾肥、喷施多效唑等[3, 4]。但甘薯高产不仅要求较强的物质生产积累、运转能力, 还应具有与其相适应的块根库器官。有研究表明, 促进甘薯生长前期根系生长发育和块根库器官分化建成, 在单株有效薯块数基本稳定的封垄期形成较多的薯块, 可以提高块根库的生长潜力, 促进块根产量提高, 如覆膜栽培等[5, 6, 7]。但目前甘薯单株有效薯块数差异的生理原因及块根分化建成的生理机制尚待进一步研究。
现有研究表明, 甘薯单株有效薯块数与最终的块根产量呈显著正相关[8], 且生长前期的根系特性与块根的分化建成密切相关[9, 10, 11, 12]; 作物根叶器官糖组分的规律性变化与植株生长速率、生长模式及目标产量器官的竞争能力密切相关[13, 14, 15], 影响最终产量[16]。本研究通过系统分析结薯数差异显著的2个甘薯品种生长前期根系生长发育特性与根叶糖组分变化规律, 进而研究甘薯块根分化建成的相关差异机制, 并通过大田试验封垄期的单株有效薯块数、收获期块根产量及产量构成因素进行验证, 旨在为大田生产中通过促块根分化建成提高块根产量的栽培技术途径提供理论依据。
试验于2013年和2014年在山东农业大学农学实验站(山东泰安)进行。供试品种为单株结薯数差异显著的淀粉型甘薯品种商薯19和济徐23, 其中, 商薯19薯块多而匀, 结薯数一般是6~7个, 而济徐23大中薯率较高, 结薯数一般是3~4个。供试土壤质地为沙壤土。2013年0~20 cm土层土壤含有机质1.44%、碱解氮67.65 mg kg-1、速效磷16.00 mg kg-1和速效钾77.83 mg kg-1; 2014年0~20 cm土层土壤含有机质1.07%、碱解氮42.11 mg kg-1、速效磷20.44 mg kg-1和速效钾125.33 mg kg-1。
本试验包括大田试验和盆栽辅助试验两部分。其中, 大田试验分别于2013年5月8日栽植、10月18日收获和2014年5月15日栽植、10月22日收获。小区面积16 m2, 行距0.80 m, 株距0.25 m, 栽植密度为5株 m-2, 基施钾肥(K2O) 24 g m-2, 以甘薯品种作为处理水平, 4次重复, 随机排列; 盆栽试验于2014年进行, 栽植时间与2014年大田试验一致, 选用高0.25 m, 上下内径分别是0.23 m和0.20 m的硬塑料盆, 取大田耕层土壤, 每盆装土10 kg, 栽植秧苗1株, 每个处理各栽20盆, 其余处理与大田试验一致。
大田试验中设置专门的取样区, 分别于封垄期(秧苗栽后45 d)和收获期取样。其中, 封垄期从每个处理选择长势一致的代表性植株5株, 将所有根径≥ 0.5 cm的根系挖出, 计算单株有效薯块数和平均单薯鲜重; 收获期计算单株有效薯块数和平均单薯鲜重, 并测定各小区块根产量。
盆栽试验为研究甘薯生长前期根系特性及块根分化建成的辅助试验, 分别于块根分化建成初期(秧苗栽后15 d)、块根分化建成后期(秧苗栽后30 d)和茎叶封垄期(秧苗栽后45 d)冲根取样, 每个处理冲根4盆、共取4株, 调查根系的根尖数目、不定根数目、不定根根长、不定根根径范围, 并对吸收根系、单株有效薯块和地上部鲜样称重, 计算生长前期地上部和总根系鲜重比值(T/TR)和封垄期地上部和块根的鲜重比值(T/SR)。同时, 将各取样时期样株全部的叶片和块根分化建成前期全部根系、块根分化建成后期和封垄期有效薯块切片装袋, 105℃杀青, 60℃烘干、磨碎, 干燥器内保存, 用于可溶性总糖、淀粉和糖组分的测定。
1.3.1 根系性状
以DT-SCAN软件分析发根缓苗期和块根分化期的根系根尖数; 人工计数不定根数目, 人工直尺测量不定根根长; 分别参照Noh等[17]和Tanaka等[18]的标准, 利用游标卡尺测量粗根和单株有效薯块的根径。
1.3.2 根叶可溶性总糖、淀粉和糖组分含量
采用蒽酮比色法测定根叶组织的可溶性总糖和淀粉含量; 采用高效液相色谱(HPLC)法测定根叶糖组分含量。称取0.1000 g粉碎的根(叶)干样, 用80%乙醇80℃水浴浸提30 min, 离心3次, 收集所有上清液并蒸干, 以蒸馏水溶解糖, 高速离心, 过C18柱去色素后用0.45 μ m微孔滤膜过滤, 滤液用于HPLC的测定。其中, 采用Waters公司测糖专用的Sugar-PAKI型柱, 流动相为双蒸馏水(0.1 mmol L -1 EDTA Na2-Ca), 流速0.5 mL min-1, 柱温90℃, Waters2414示差折光检测器, 进样量为10 μ L。根据标样的色谱峰面积计算葡萄糖(glucose)、果糖(fructose)、蔗糖(sucrose)、蔗果三糖(1-kestose)、蔗果四糖(nystose)和蔗果五糖(1F-fructofuranosylnystose)的含量。
用SigmaPlot 10.0软件制图, SAS 9.2软件分析数据, 以单因素随机区组法分析收获期块根产量的方差、单因素完全随机法分析其他项目的方差, 利用Duncan’ s新复极差法检验处理间的差异显著性。
表1为2013— 2014年大田统计数据, 其中, 封垄期商薯19的单株有效薯块数、单株有效薯块鲜重均显著高于济徐23 (F=10.71, P=0.0170; F=13.97, P=0.0212); 收获时商薯19具有显著多的单株有效薯块数和显著高的块根产量(F=353.89, P< 0.0001; F=88.94, P< 0.0001), 而济徐23具有显著高的平均单薯鲜重(F=10.32, P=0.0124)。
| 表1 封垄期和收获期与产量有关的性状(大田, 2013-2014) Table 1 Yield traits and fresh yield of storage root in field trials at top cover stage and harvest period in 2013-2014 |
大田收获期产量性状与块根产量的偏相关分析结果(表2)显示, 单株有效薯块数、平均单薯鲜重与块根产量均呈显著正相关, 其中商薯19的单株有效薯块数与块根产量的相关性要大于平均单薯鲜重与块根产量的相关性。
| 表2 收获期产量性状与块根产量间的偏相关系数(大田, 2013-2014) Table 2 Partial correlated coefficients among yield traits in field trials at harvest period in 2013-2014 |
2.2.1 不定根、侧根生长发育
表3表明, 2个品种随着根系的生长发育, 植株的根尖数、不定根条数、不定根根长密度、粗根数和单株有效薯块数均显著升高。其中, 秧苗栽后15、30 d商薯19均具有较高的不定根条数、不定根根长密度和粗根数; 同时, 商薯19秧苗栽后15 d具有较高根尖数, 形成的不定根数占30 d不定根数的76.91%, 在秧苗栽后30 d所形成的有效薯块数占45 d有效薯块数的52.22%。而济徐23秧苗栽后15 d与30 d不定根数目、30 d和45 d的单株有效薯块数均无显著差异(F=3.375, P=0.1401; F=0.2500, P=0.6433), 说明商薯19在秧苗栽后30~45 d间, 依然有块根的分化建成, 而济徐23在秧苗栽后15 d基本完成了不定根的发生, 秧苗栽后30 d基本完成不定根向块根的分化建成。
| 表3 单株的根系特性(盆栽, 2014) Table 3 Root characteristics at early growth stage in pot trials in 2014 |
2.2.2 有效薯块的形成
从表3还可以看出, 秧苗栽后30 d, 商薯19具有较多根径介于0.5~1.0 cm的幼薯(YSR, 0.5 cm≤ Φ < 1.0 cm), 而济徐23具有较多根径介于1~5 cm的幼薯(YSR, 1 cm≤ Φ < 5 cm); 秧苗栽后45 d, 商薯19具有更多的单株有效薯块数(F=27.00, P=0.0065), 其粗根数和根径介于0.5~1.0 cm的幼薯(YSR, 0.5 cm≤ Φ < 1.0 cm )数目均显著高于济徐23, 而根径大于5 cm的单株有效薯块(MSR, Φ > 5 cm )显著少于济徐23。说明在甘薯生长前期, 商薯19更为侧重于块根的分化建成, 而济徐23更为侧重于块根分化建成后的生长发育。
| 表4 单株的物质生产特性(盆栽, 2014) Table 4 Material production characteristics at early growth phase in pot trials in 2014 |
由表4可见, 秧苗栽后15 d商薯19具有较高地上部、根系鲜重, 较低的地上部与根系鲜重比值(T/TR), 更侧重根系生物量的积累; 而在秧苗栽后30 d, 2个品种间具有相似的地上部、根系鲜重和T/TR值, 其中商薯19的T/TR值与其15 d时相似, 且相较于济徐23具有较大的吸收根系鲜重; 秧苗栽后45 d, 2个品种具有相似的有效薯块鲜重和T/TR值, 其中商薯19具有较大的地上部、根系和吸收根鲜重, 而济徐23具有较低的地上部与块根鲜重比值(T/SR)。
2.3.1 蔗糖含量
由图1可知, 2个品种各取样时期根系蔗糖含量随着根系的生长发育而升高, 秧苗栽后45 d根系的蔗糖含量显著(F=698.31, P< 0.0001)高于栽后30 d, 而秧苗栽后30 d显著(F=19.61, P= 0.0013)高于15 d。其中, 商薯19在整个生长前期(秧苗栽后15、30和45 d)根系的蔗糖含量均显著低于济徐23。
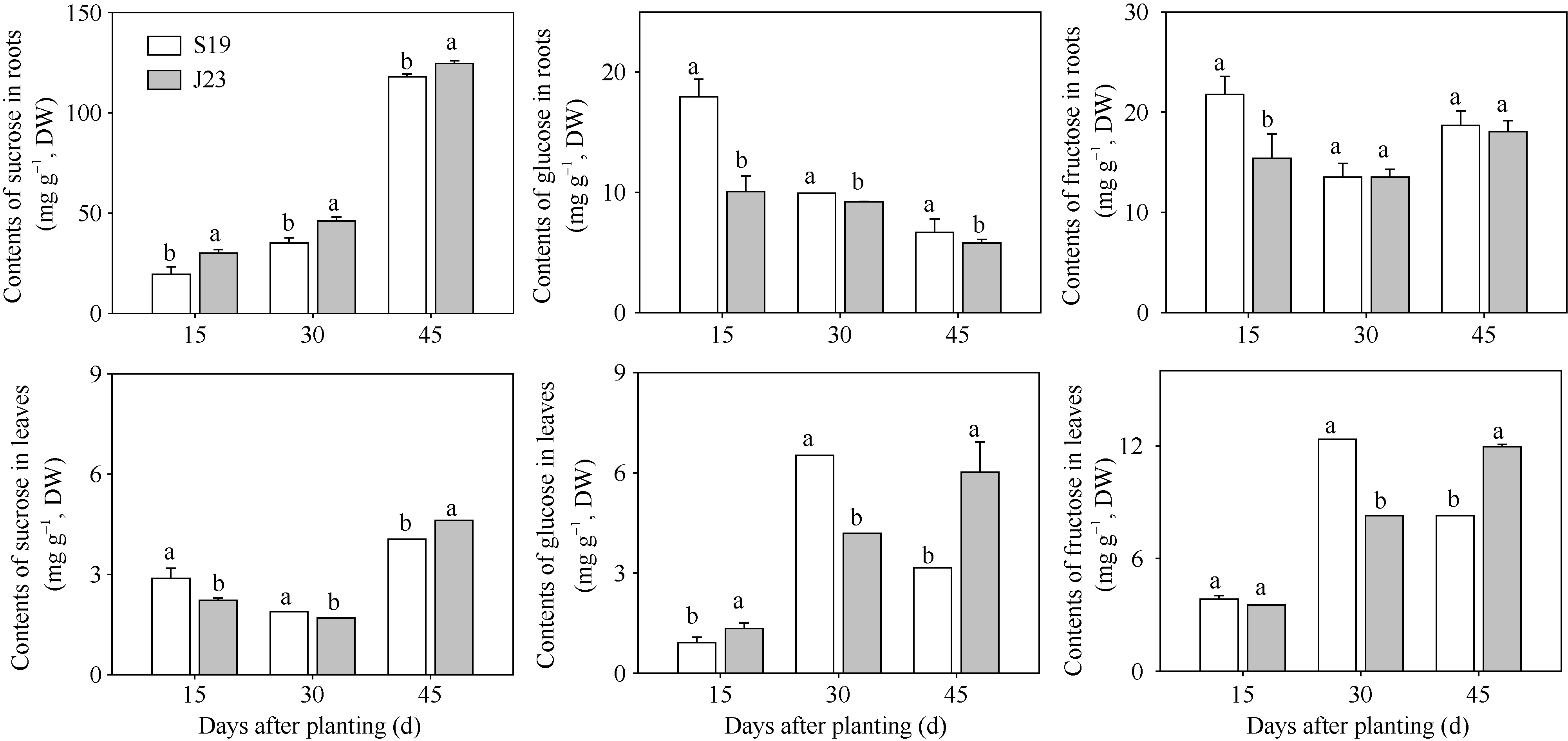 | 图1 秧苗栽后15、30和45 d甘薯根系和叶片蔗糖、葡萄糖、果糖的含量变化(盆栽, 2014) S19: 商薯19; J23: 济徐23。图柱上标以不同小写字母表示糖含量在0.05水平上差异显著。Fig. 1 Changes of contents of sucrose, glucose and fructose at 15, 30, and 45 days after planting in root and leaf in pot trials in 2014 S19: Shangshu 19; J23: Jixu 23. Bars superscripted by different letters are significantly different at the 0.05 probability level among sugar contents. |
叶片的蔗糖含量在秧苗栽后45 d最高(F=361.38, P< 0.0001), 秧苗栽后30 d最低(F=19.36, P=0.0013)。其中, 商薯19秧苗栽后15 d、30 d叶片的蔗糖含量显著高于济徐23, 使得商薯19在根叶间形成了更大的蔗糖浓度梯度。
2.3.2 葡萄糖含量
在甘薯生长前期根系的葡萄糖含量随着根系的生长发育而逐渐降低, 秧苗栽后45 d根系的葡萄糖含量显著(F=16.61, P=0.0022)低于栽后30 d, 秧苗栽后30 d显著(F=8.16, P=0.0170)低于栽后15 d。其中, 商薯19整个生长前期根系的葡萄糖含量均显著高于济徐23。
叶片的葡萄糖含量在秧苗栽后15 d最低(F= 20.45, P< 0.0001), 而秧苗栽后30 d、45 d间无显著差异(F=0.79, P=0.3936)。其中, 商薯19秧苗栽后15 d、45 d叶片的葡萄糖含量显著低于济徐23, 而秧苗栽后30 d显著高于济徐23。
2.3.3 果糖含量
根系的果糖含量在秧苗栽后30 d最低(F=9.42, P=0.0022), 秧苗栽后15 d、45 d间无显著差异(F=0.02, P=0.8980)。其中, 商薯19秧苗栽后15 d根系果糖含量显著高于济徐23, 而秧苗栽后30 d、45 d, 2个品种间无显著差异。
叶片的果糖含量与叶片葡萄糖含量具有相同变化规律, 在秧苗栽后15 d最低(F=28.38, P< 0.0001), 而秧苗栽后30、45 d间无显著差异(F=0.03, P=0.8679)。其中, 2个品种秧苗栽后15 d叶片的果糖含量无显著差异, 而商薯19秧苗栽后30 d显著高于济徐23, 秧苗栽后45 d显著低于济徐23。
2.4.1 淀粉含量
由图2可知, 2个品种根系、叶片淀粉含量具有相同的变化规律, 秧苗栽后30 d淀粉含量高于15 d (根系F=129.59, P< 0.0001; 叶片F=16.72, P=0.0022), 秧苗栽后45 d高于30 d (根系F=216.59, P< 0.0001; 叶片F=265.75, P< 0.0001)。商薯19秧苗栽后15、45 d根系的淀粉含量均显著的低于济徐23, 栽后30 d无显著差异; 商薯19秧苗栽后15 d叶片的淀粉含量显著高于济徐23, 而栽后30 d、45 d均显著的低于济徐23。
 | 图2 秧苗栽后15、30和45 d甘薯根系、叶片淀粉和可溶性总糖的含量变化(盆栽, 2014) S19: 商薯19; J23: 济徐23。图柱上标以不同小写字母表示糖含量在0.05水平上差异显著。Fig. 2 Changes of contents of starch and total soluble sugar in roots and leaves at 15, 30, and 45 days after planting in pot trials in 2014 S19: Shangshu 19; J23: Jixu 23. Bars superscripted by different letters are significantly different at the 0.05 probability level among sugar contents. |
2.4.2 可溶性总糖含量
根系、叶片可溶性总糖含量具有相同的变化规律, 秧苗栽后15 d、30 d间可溶性总糖含量没有显著差异(根系F=0.45, P=0.5182; 叶片F=3.71, P=0.0830), 而秧苗栽后45 d可溶性总糖含量显著升高(根系F=344.46, P< 0.0001; 叶片F=344.46, P< 0.0001)。其中, 商薯19整个生长前期根系、叶片的可溶性总糖含量均不低于济徐23。
随着根系的生长发育(表5), 2个品种根系的蔗糖/己糖比率逐渐升高, 秧苗栽后30 d根系的蔗糖/己糖比率显著(F=29.24, P=0.0003)高于栽后15 d, 栽后45 d显著(F=215.68, P< 0.0001)高于栽后30 d, 其中秧苗栽后15、30和45 d商薯19根系的蔗糖/己糖比率均显著低于济徐23。说明在整个生长前期, 商薯19更为侧重己糖的代谢。
| 表5 秧苗栽后15、30和45 d根系的糖组分比率(盆栽, 2014) Table 5 Changes of content ratio of carbohydrate composition in root at 15, 30, and 45 days after planting in pot trials (2014) |
2个品种根系的蔗糖/可溶性总糖比率也随着根系的生长发育逐渐升高, 秧苗栽后30 d根系的己糖/可溶性总糖比率显著(F=24.93, P=0.0003)高于15 d, 栽后45 d显著(F=120.00, P< 0.0001)高于栽后30 d。其中, 秧苗栽后15 d、30 d济徐23根系的蔗糖/可溶性总糖比率均显著高于商薯19, 而栽后45 d品种间无显著差异。说明在根系可溶性总糖的代谢中, 蔗糖代谢比例随着根系的生长发育而不断升高, 其中秧苗栽后15 d、30 d时, 济徐23在可溶性总糖的代谢中具有更高的蔗糖代谢比例。
2个品种根系的蔗糖和淀粉含量均随着根系的生长发育显著升高(表3和表4), 但栽后15 d和30 d间根系的蔗糖/淀粉比率没有显著差异(F=0.92, P=0.3573), 栽后45 d时根系的蔗糖/淀粉比率才显著升高(F= 215.68, P< 0.0001)。说明在秧苗栽后15 d、30 d, 根系的蔗糖、淀粉含量只在绝对含量上有差异, 二者含量的相对变化比率相似; 而秧苗栽后45 d, 根系蔗糖、淀粉含量显著升高的同时, 蔗糖/淀粉比率也显著升高, 说明蔗糖代谢的变化大于淀粉代谢的变化。其中, 秧苗栽后45 d济徐23更为侧重于淀粉的转化积累。
2.6.1 根系果聚糖含量变化
由表6可知, 整个生长前期在2个品种的根系中均未检测出蔗果五糖, 秧苗栽后15 d、30 d根系的蔗果四糖含量间没有显著差异(F=0.01, P=0.9358), 栽后45 d蔗果四糖含量显著降低(F=24.23, P< 0.0001)。其中, 秧苗栽后15 d、30 d商薯19根系的蔗果四糖含量均显著低于济徐23, 但商薯19根系低果聚糖总含量(蔗果三糖+蔗果四糖)与济徐23根系中低果聚糖总含量(仅蔗果四糖)相似。此外, 45 d可能因蔗果四糖的解聚, 或因蔗果四糖合成反应的弱化, 促使济徐23根系蔗果三糖合成。在秧苗栽后15 d、30 d, 商薯19根系的蔗果三糖含量间没有显著变化差异(F=0.43, P=0.5263), 栽后45 d显著降低(F=5.33, P=0.0178), 且与济徐23的蔗果三糖含量无显著差异。
| 表6 秧苗栽后15、30和45 d根系果聚糖含量(2014) Table 6 Changes of fructosan content in root at 15, 30, and 45 days after planting in pot trials in 2014 (mg g-1 DW) |
2.6.2 叶片果聚糖含量变化
由表7可知, 整个生长前期在2个品种的叶片中均未检测出蔗果三糖, 且随着植株的生长, 叶片蔗果四糖、蔗果五糖含量先升高(F=20.89, P=0.01; F=936.52, P=0.0001)后又下降(F=35.96, P=0.004; F=2307.35, P=0.0001), 且栽后15 d时2个品种叶片的蔗果四糖、蔗果五糖含量间均无显著差异(F=2.78, P=0.24; F=0.83, P=0.06), 栽后30 d济徐23的蔗果四糖含量显著最高, 而商薯19的蔗果五糖含量显著最高, 栽后45 d商薯19的蔗果四糖和蔗果五糖均显著最低。
| 表7 秧苗栽后15、30和45 d叶片果聚糖含量(2014) Table 7 Changes of fructosan content in leaf at 15, 30, and 45 days after planting in pot trials in 2014 (mg g -1DW) |
甘薯茎叶封垄期的单株有效薯块数基本稳定[6, 19, 20, 21], 在本研究中单株有效薯块数较多的商薯19在收获期具有更高的块根产量, 且单株有效薯块数与块根产量的正相关性大于平均单薯鲜重与块根产量, 说明促进甘薯块根的分化建成, 形成多的单株有效薯块数, 对于最终块根产量的提高具有重要的意义。大量研究表明, 甘薯生长前期根系特性与块根的分化建成密切相关[9, 10, 11, 12]: 长度大于20 cm的不定根更容易分化建成块根[9, 11], 同时不断发生的侧根, 会扩大吸收根系, 促进土壤水分和营养元素的吸收利用[22], 并提高根系获取碳同化物的能力[23]和其着生不定根向块根分化的能力[11, 12], 且甘薯形成早的根原基多发育成粗根, 易分化为块根[9, 24]。此外, 地上部生物量与总根系生物量比值(T/TR)说明甘薯茎叶和根系生长的动态变化, 地上部与块根生物量比值(T/SR)是判别甘薯茎叶源与块根库关系是否协调的直观指标[] [6]。在本研究中, 商薯19发根缓苗迅速, 在块根分化建成前期具有较高的地上部、根系生物量和较低的T/TR值, 且块根分化建成过程中T/TR值较为稳定, 整个生长前期均较为侧重吸收根系的建成, 促进了粗根的形成及其向块根的分化建成, 在块根分化建成后期至茎叶封垄仍持续分化建成有效薯块, 是商薯19形成较多单株有效薯块数的基础性条件; 而济徐23较早完成不定根的发生(秧苗栽后15 d)和有效薯块的分化建成(秧苗栽后30 d), 封垄期时具有较低的T/SR值, 侧重已形成有效薯块的生长发育。
蔗糖是光合作用的主要产物[25], 同化物运输的主要形式和库器官代谢的主要基质[26, 27], 而源库器官间的蔗糖浓度梯度是蔗糖由源向库的运输驱动力[28, 29], 叶片较高的蔗糖浓度利于蔗糖的向下运输[30, 31], 库器官对蔗糖的利用促进蔗糖浓度梯度的形成[32]。同时, 己糖和蔗糖代谢分别对应库器官不同的生长发育阶段: 高水平的己糖对应于发育器官的细胞分裂分化, 而高浓度蔗糖会触发贮藏物质的积聚[32, 33, 34]。萝卜肉质根较高的蔗糖浓度, 抑制蔗糖卸载阻碍肉质根的膨大, 而较低的蔗糖浓度, 较高的葡萄糖、果糖浓度, 利于肉质根的形成和发育[35, 36]。对蚕豆籽粒、杨梅果实发育的研究表明[37, 38], 当蔗糖/己糖比率较低时, 籽粒或果实具有较高的细胞分裂活性; 蔗糖/己糖比率增大, 籽粒或果实便从细胞分裂阶段过渡到贮藏阶段。本研究发现, 商薯19生长前期具有较低的根系蔗糖含量, 较高的根系可溶性总糖、己糖含量和较大的根叶蔗糖浓度梯度; 同时, 根系的蔗糖/己糖比率随着甘薯根系的生长发育逐渐升高, 而整个生长前期商薯19根系的蔗糖/己糖比率均显著低于济徐23。初步分析认为, 在甘薯生长前期己糖、蔗糖代谢亦会通过影响不同的生理活动, 调控根系的生长发育、块根库器官的分化建成。其中, 商薯19在整个生长前期根叶间具有更强的蔗糖运输驱动力, 且根系具有更为活跃的己糖代谢, 具有较高的块根分化建成潜力。
一般源器官运输到库器官的蔗糖, 或作为碳源、能源分解成己糖等供新生组织的生长发育, 或转变成用于贮藏的糖类多聚物[39]。其中, 淀粉是葡萄糖多聚物, 而果聚糖是在蔗糖的基础上, 链接上一个或多个果糖残基聚合而成。有研究表明, 源叶淀粉的积累会抑制马铃薯块茎的形成[40]; 大麦等作物库器官淀粉的积累要早于其蔗糖/己糖比率的变化[41, 42]; Wang等[43]研究发现, 甘薯根系淀粉、蔗糖含量与薯块大小高度相关。同时, 果聚糖的合成、降解与植物碳素分配, 源库关系调节有密切关系[44], 其对器官的启动生长及最终产量具有重要作用[45, 46, 47, 48]。果聚糖的合成可以提高韧皮部至新生细胞间蔗糖的浓度梯度[49]; 在小麦、大麦籽粒和蒲公英幼根生长早期阶段果聚糖会迅速而相对大量地合成[46, 47, 50]。同时, 淀粉生物合成对蔗糖的竞争作用[51, 52]使小麦、大麦的籽粒淀粉快速积累时[45, 46]与牧草快速生长前蔗糖快速积累时[48, 53]均伴随果聚糖的迅速降解。
本研究中2个甘薯品种的根叶器官在生长前期的不同阶段侧重于不同贮藏糖类的积累, 块根分化建成初期根系低聚合度果聚糖的合成相对旺盛, 2个品种根系低果聚糖总含量相似; 而块根分化建成后, 分化根主要进行蔗果四糖的降解和淀粉的合成积累, 同时茎叶封垄期2个甘薯品种根系的蔗糖/淀粉比率均显著升高, 要晚于蔗糖/己糖比率的变化, 说明甘薯块根分化建成中淀粉积聚的发生要晚于细胞的分裂分化; 在整个生长前期, 叶片淀粉含量持续稳定增长, 而叶片低聚合度果聚糖在块根分化建成后期开始相对大量合成。其中, 蔗果三糖、四糖、五糖的合成在积聚利用等量蔗糖的前提下, 实现了己糖果糖的不同利用。所以在甘薯块根分化建成初期, 2个品种在具有相似根系低果聚糖总量的前提下, 商薯19根系通过蔗果三糖的存在, 相对减少了对己糖果糖的利用, 更利于根系己糖的代谢; 在块根分化建成后期, 商薯19叶片通过合成更多的蔗果五糖, 实现对己糖果糖更多的贮藏积累, 而较低的叶片淀粉含量利于其根系向块根的分化建成; 在2个甘薯品种具有相似根系蔗糖/可溶性总糖比率前提下, 济徐23根系的蔗糖/淀粉比率显著更低, 说明茎叶封垄时济徐23更为侧重块根分化建成后淀粉的转化积累和已形成薯块的生长发育。
商薯19生长前期具有较高的生物量积累且侧重吸收根系的生长, 不定根、侧根的形成发育时间早且持续期长, 利于粗根的形成及其向有效薯块的分化建成; 在根叶蔗糖、己糖代谢和贮藏糖类多聚物的形成中, 糖的组成及其代谢流向影响了甘薯块根库器官早期的发育: 商薯19整个生长前期旺盛的己糖代谢促进不定根、侧根的形成和生长发育, 利于粗根的形成及其向块根的分化建成; 济徐23块根分化建成过程中显著高的根系蔗糖/己糖比率, 使其较早完成不定根的发生和有效薯块的分化建成, 而块根分化建成后显著低的蔗糖/淀粉比率, 利于已形成有效薯块的生长发育。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|