

{kind=link}
{kind=link}
{kind=link}
谷子核心种质表型遗传多样性分析及综合评价
[王海岗1 , 贾冠清2 , 智慧2 , 温琪汾1 , 董俊丽1 , 陈凌1 , 王君杰1 , 曹晓宁1 , 刘思辰1 , 王纶1 , 乔治军1, *  , 刁现民
, 刁现民2, * ]
, 刁现民]
|
|


第一作者联系方式: E-mail: nkywhg@126.com, Tel: 0351-7639230
通过遗传种质的多样性评估, 可以指导深入研究资源和育种中优异互补亲本的选择, 进而提高优异基因的交流累加和新品种培育的效率。本研究选用了来自世界各地的878份谷子核心种质通过15个表型性状的综合鉴定, 评估遗传多样性和筛选优异种质资源, 结果表明: (1)我国谷子资源的表型遗传多样性丰富, 单穗粒重、穗长、穗粗、株高、茎节数和生育期均表现了丰富的变异; 谷子育成品种遗传多样性相比农家品种下降明显, 育种的遗传增益主要体现在株高和穗长的适度减低, 以及茎粗、茎节数、穗粗、单穗粒重、单穗重及生育期的适度增加; (2)系统聚类分析将谷子资源分成3类, 第I类以来源为东北欧国家的品种为主, 第II类以北美和非洲的品种为主, 第III类以东亚、南亚的品种为主; 我国种质可划分为春播型、春夏兼播型和南方型3类型; (3)采用主成分分析法和逐步回归分析法综合评判表明, 叶鞘色、刚毛长度、粒色、米色、株高、穗长、茎粗和单穗粒重8个性状可作为谷子表型鉴定的主要指标。
, DIAO Xian-Min
Evaluation of genetic diversity could benefit the identification of optimal parental combinations for obtaining segregating offspring with maximum genetic variability, and facilitate the introgression of favorable genes from various germplasm into commercial cultivars. In this study, foxtail millet core collections including 878 world-wide accessions were evaluated through phenotypic analysis of 15 agronomic traits. Main conclusions are as follows: (1) Chinese accessions present higher level of phenotypic diversity, especially in grain weight per main stem, panicle length, panicle diameter, plant height, stem node number and growth period; phenotypic diversity is lower in elite cultivars than in traditional landraces; gains of breeding progress in foxtail millet are expressed in reduced plant height and panicle length, coupling with increased diameter of main stem, stem node number, panicle diameter, grain weight per main stem, panicle weight per main stem and growth period; (2) from the cluster analysis, foreign resources are divided into three categories according to the geographical origins, the first category mainly includes accessions from northeast European countries, the second cluster is mainly collected from North America and Africa, and the third group includes varieties mainly from East Asia, South Asia, African, and European countries; Chinese collections is able to be divided into three groups of spring-sowing, summer-sowing and southern ecotypes; (3) comprehensive assessment of phenotypic traits based on principal component analysis (PCA) and step regression analysis demonstrates that leaf sheath color, bristle length, hull color, grain color, plant height, spike length, stem diameter and grain weight per plant can be used as main identification indicators of foxtail millet phenotypic variations. This research will benefit the utilization of foxtail millet resources in variety breeding practices in the future.
谷子(Setaria italica Beauv.)是起源于中国的古老农作物, 具有抗旱、耐瘠、适应性强等特点, 籽粒和谷草均有较高的营养价值、粮饲兼用, 是我国北方干旱半干旱地区重要的农作物。我国谷子总产约占世界的80%, 其次为印度约占10%, 其他国家如澳大利亚、美国、加拿大、法国、朝鲜、日本、匈牙利等国家也有少量种植[1]。我国的谷子生产主要分布在北方干旱、半干旱地区, 其中2/3分布在干旱最严重的华北地区。目前, 全国谷子年种植面积约200万公顷, 年总产350万吨左右, 种植面积较大的省区依次是河北、山西、内蒙古、陕西、辽宁、河南、山东、黑龙江、甘肃和吉林等; 同时, 谷子作为优良饲草在农牧交接区广泛栽培[2]。谷子中含有丰富的蛋白质、叶酸、维生素E、类胡萝卜素及硒, 作为营养均衡作物对维持人体健康具有重要作用[3, 4, 5]。我国谷子栽培历史悠久, 最新的考古研究表明谷子的驯化栽培可追溯至11 500年前, 并在距今6000年左右的仰韶文化时期发展成为主要粮食作物和中国农耕文化的代表作物[6]。谷子作为中国北方主栽作物的历史悠久, 在漫长的自然选择和人工选择条件下, 形成了丰富的遗传变异, 传播到世界各地后, 也形成了多样的类型[7]。
作物种质资源是种质创新和生物学研究的物质基础, 因此种质资源的收集和保存长期以来得到了广泛重视。但随着种质资源数量的不断增长, 如何开展对海量资源的研究利用是个重要命题。核心种质采用最小的资源样本数量最大程度上代表资源的遗传多样性[8], 为海量资源提供了高效利用的方式。水稻[9, 10]、小麦[11, 12]、玉米[13, 14]、大豆[15, 16]等都已构建了核心种质, 有的作物在此基础上建立了比核心种质样本量稍大, 更接近育种实践的应用核心种质[17], 并以应用核心种质为基础开展表型、基因型等方面的精准鉴定和相关的深入研究, 这对加深种质资源的科学认识和遗传育种具有重要意义。谷子基因组小、二倍体、生育期短、单株种子量大, 正在发展成为包括玉米(Zea mays)、高粱(Sorghum bicolor)、甘蔗(Saccharum officinarum)、珍珠粟(Pennisetum glaucum)、黍稷(Panicum miliaceum)等作物在内的禾本科黍亚科和C4光合作用研究的模式作物[18], 谷子在作物遗传育种研究中的地位日趋重要。目前, 国家中长期种质库中保存有各类谷子资源27 000余份[19], 前期研究已构建了一个涵盖各类谷子资源的应用核心种质[7], 但对应用核心种质在不同环境下的综合农艺性状鉴定依然缺乏, 影响了该基因库在我国不同谷子生态区的高效利用。
已有的研究[20, 21, 22, 23, 24, 25, 26]表明, 我国谷子种质资源主要分为地方品种、育成品种以及国外资源3种类型, 通过形态学、细胞学、生化及分子标记等不同研究手段[27], 已经完成部分谷子遗传资源的品质[3, 4, 5]、抗旱性[28]、抗病性[29]等性状的评价及优异种质的鉴定筛选。随着我国谷子遗传育种工作的不断深入, 已有研究结果在资源代表性和数量上依然十分有限, 亟需开展对我国大量谷子种质资源表型的统一鉴定。本研究旨在对我国谷子应用核心种质进行综合表型鉴定, 为深入认识谷子应用核心种质的表型多样性以及为谷子资源的遗传与育种的深入应用奠定基础。
谷子应用核心种质878份中, 农家品种491份, 育成品种294份和国外种质93份。该应用核心种质由中国农业科学院作物科学研究所构建, 取样综合了中国国家种质库中谷子种质资源在地理来源、多样性中心、农家品种和育成品种不同类型, 以及国外品种等方面的代表性, 最大程度地涵盖了我国国家种质库中谷子的遗传多样性类型[7]。
2012— 2013年, 连续2年在山西省农业科学院东阳试验基地(37.6° N, 112.7° E)对878份谷子资源15个表型性状进行调查。每份材料种2行, 行长2.5 m。按照《谷子种质资源描述规范和数据标准》[30]记录各指标。其中, 6个质量性状, 包括叶鞘色、穗型、刚毛长度、穗颈形状、粒色、米色; 9个数量性状, 包括株高、穗长、穗下节间长度、茎粗、主茎节数、主穗直径、单穗重、单穗粒重, 生育期。性状的选取综合考虑了资源和遗传育种研究的需要, 以及性状精准调查的稳定可靠性等, 从而使结果更可靠和实用。
采用Microsoft Excel 2003和Statistic 6.0整理和分析数据。采用DPS7.05进行聚类分析、相关分析和主成分分析, 并计算各主成分得分及综合得分, 再结合逐步回归分析筛选谷子资源综合性状评价指标。采用模糊隶属函数计算出各性状的隶属函数值, μ (xi)=(xi-ximin)/(ximax-ximin) (i=1, 2, 3, …, 878), 即将各性状定义到[0, 1]闭区间。式中, μ (xi)为某种质材料第i个性状的隶属函数值, xi为某种质材料的第i个性状值, ximax、ximin分别为所有种质资源中第i个性状的最大值和最小值。采用Shannon-Wiener’ s多样性指数(Shannon-Wiener diversity index, H')进行遗传多样性评价。
2.1.1 质量性状
谷子应用核心种质中叶鞘有56.09%为绿色, 24.07%为紫色, 16.41%为淡紫色; 穗型以纺锤形最多, 占69.05%, 其次是圆锥和圆筒形, 分别占10.28%和9.12%; 刚毛长度多数在2~4 mm, 占65.47%; 穗颈形状以弯曲型为主, 占84.87%; 77.46%的种质粒色为黄色, 其次为红色(8.27%)、白色(5.64%)和橙色(4.20%); 应用核心种质中92.09%的米色为黄色, 其他为白色, 表明富含类胡萝卜素的黄色种质是谷子米色的主要类型。
2.1.2 数量性状
由表1可知, 9个数量性状的变异系数中, 生育期最小, 为7.94, 单穗粒重最大, 为43.41; 表型性状遗传多样性指数, 主穗直径最小, 为0.67, 单穗重最大为1.84。9个数量性状的变异均符合正态分布, 株高平均值为139.17± 20.94 cm, 变幅为45.00~195.00 cm, 极差为150.00; 其中, 来自印度的种质Y-859最高, 为195.00 cm, 中国河北育成品种衡早1号最矮, 为45 cm。
茎粗平均值为0.68± 0.12 cm, 变幅为1.00~63.50 cm, 极差为0.95; 其中, 来自美国的Y-806和黎巴嫩的Y-812最小为0.25 cm, 中国河南育成品种安矮8号最大为1.20 cm。
主茎节数平均值, 为11.73± 1.84, 变幅为4.00~ 17.00, 极差为13; 其中, 来自河北的育成品种衡早1号主茎节数最, 少为4个, 沧555最多, 为17个。
穗下节间长度平均值为10.84± 4.47 cm, 变幅在1.00~36.25之间, 极差为35.25; 其中, 来自中国河南的育成品种安矮9号和安矮17最短, 为1 cm, 来自法国的Y-759最长, 为36.25 cm。
穗长平均值为23.98± 5.62 cm, 变幅在5.25~43.00之间, 极差为37.75; 其中, 俄罗斯资源Y-816最短, 为5.25 cm, 来自中国甘肃的农家种昆昌谷及西班牙种质Y-837最长, 为43.00 cm。
主穗直径平均值为2.54± 0.61 cm, 变幅为0.95~ 5.15 cm, 极差为4.2; 其中, 来自美国的Y-778最小, 0.95 cm, 来自中国河南的农家品种龙爪粘谷最大, 为5.15 cm。
单穗重的平均值为15.03± 5.95 cm, 变幅为0.20~ 39.50 g, 极差为39.30; 单穗粒重的平均值为11.85± 5.14 cm, 变幅为0.15~34.20 g, 极差为34.05; 单穗重和单穗粒重是谷子产量的构成因子, 中国甘肃农家品种红金转单穗重和单穗粒重最高, 分别为39.50 g和34.20 g。
| 表1 878份谷子资源的数量性状变化及分布特征 Table 1 Variation and distribution of quantitative traits in 878 foxtail millet accessions |
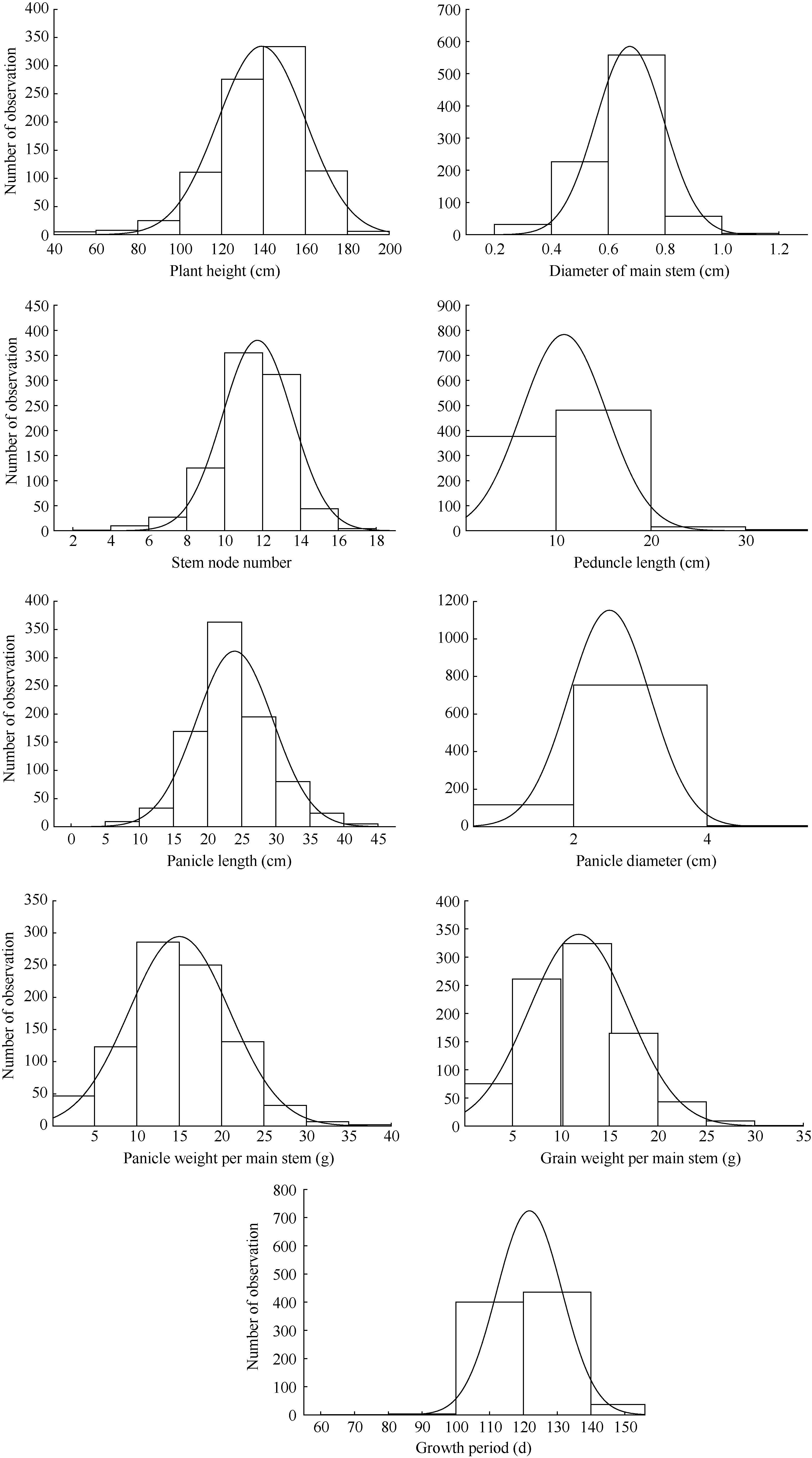 | 图1 878份谷子资源主要数量性状指标的分布情况Fig. 1 Distribution diagram of quantitative traits of 878 foxtail millet accessions |
生育期作为种质适应性的重要指标, 平均值为121.79± 9.67 d, 变幅为56~155 d; 其中, 来自印度的种质Y-859生育期最长, 为155 d。
我国种质库的谷子资源从整体上可分为国外引进品种、20世纪50至80年代搜集整理的农家品种、以及从20世纪70年代后期通过系统选择、杂交育种和诱变育种培育的新品种3种类型。从表2的不同品种类型分析结果可以清楚地看出, 国外引进品种9个性状的变异系数均大于我国的农家品种和育成品种, 如国外种质的单穗粒重变异系数高达65.17, 而我国的农家品种和育成品种单穗粒重的变异系数分别为41.78和32.24; 我国的农家品种的变异系数总体上高于育成品种, 9个性状中穗长、单穗重、单穗粒重等7个性状高于育成品种, 株高和茎粗2个性状略低于育成品种; 国外品种和农家品种存在大量变异, 是品种持续改良的基因基础。
从农家品种和育成品种的比较来看, 育成品种的平均株高降低了4.32 cm, 整体上向变矮发展, 但株高减低的幅度并不明显; 育成品种的穗长较农家品种减少了1.60 cm, 这与育成品种多采用高密度栽培依靠群体增产的方式有关; 育成品种较农家品种的穗下节间长度的变异系数减少了7.48, 这与育成品种的抗倒伏性增强和高密度栽培有关; 茎粗在农家品种和育成品种之间没有发生大的变异; 主穗直径虽然在农家品种和育成品种之间没有大的改变, 但变异系数减少了10.27, 是所有调查性状中改变最显著的, 这是育种家选择紧穗型谷子品种的结果; 单穗重和单穗粒重2个产量相关性状, 育种品种较农家品种分别提高了3.61 g和3.12 g, 这是育种选择的结果; 从生育期来看, 育成品种较农家品种平均增加2.70 d, 农家品种中一些生育期极短的品种在育种过程中已被淘汰。
| 表2 不同类型谷子资源主要表型性状比较 Table 2 Comparison of quantitative trait of foxtail millet among different types |
稳定性是种质资源的一个重要构成要素(表3), 在同一生态环境下连续两年的谷子资源表型性状平均值除穗下节间长度外, 其他8个数量性状均2012年大于2013年; 两年间9个数量性状的变异系数虽有所浮动, 但表现了相同的趋势, 变化较为明显的是穗下节间长度、主穗直径、单穗重和单穗粒重, 这些指标都与产量构成有着重要关联。
| 表3 2012-2013年主要数量性状年度间变异 Table 3 Genetic variation and distribution of quantitative traits in 2012 and 2013 |
表4表明, 15个表型性状间存在不同程度的相关性。其中, 粒色与米色极显著正相关; 株高与叶鞘色、穂型、刚毛长度、米色和穗下节间长度相关不显著, 与其他性状相关性显著; 穗长与穂型、刚毛长度和穗下节间长度相关不显著, 与其他性状相关极显著; 穗下节间长度与穗颈形状、主茎节数、主穗直径和生育期呈极显著负相关; 主穗直径与叶鞘色相关不显著, 与其他13个性状均显著相关; 单穗重和单穗粒重与穗颈形状呈极显著负相关, 与粒色、米色、株高、穗长、茎粗、主茎节数、主穗直径呈极显著正相关; 生育期与叶鞘色、粒色和米色相关不显著, 与其他性状相关极显著。
前8个主成分的累计贡献率达80%以上。第1主成分贡献率最大, 为26.03%, 其中, 单穗重、单穗粒重和主茎节数分值较高。
由表5可知, 第1主成分贡献率为26.02%, 主茎节数、单穗重和单穗粒重值大于其他性状, 说明第1主成分由主茎节数、单穗重和单穗粒重组成; 第2主成分贡献率为13.15%, 以叶鞘色、粒色和米色为主, 说明第2主成分是叶鞘色、粒色和米色的综合反映; 第3主成分贡献率为10.62%, 穗颈形状和生育期大于其他性状, 说明第3主成分综合了穗颈形状和生育期性状; 第4主成分贡献率为7.96%, 株高和穗长系数大于其他性状, 说明第4主成分是株高和穗长性状; 第5主成分贡献率为6.91%, 穗型和刚毛长度最大, 说明第5主成分由穗型和刚毛长度组成。第6主成分贡献率为6.42%, 穗下节间长特征向量值最高, 表明第6主成分为穗下节间长; 第7主成分贡献率为5.62%, 主穗直径值最大, 说明第7主成分为主穗直径因子; 第8主成分贡献率为5.20%, 茎粗特征向量最大, 说明第8主成分为茎粗因子。
| 表5 谷子应用核心种质表型性状的主成分分析 Table 5 First eight principal components based on 15 phenotypic traits of 878 foxtail millets |
对15个表型性状值进行标准化处理, 求得各种质的8个主成分得分, 将8个主成分得分规一化处理, 计算各主成分权重系数(0.319、0.161、0.130、0.097、0.084、0.078、0.069、0.063), 最后求得每份种质材料的综合得分F值, 进而对各个种质综合评价。应用核心种质的平均F值为0.555, 我国育成品种中排在前10位的是延谷13、长生08、坝谷233、公谷67、长农38、晋谷24、晋谷11、长生07、长生04和冀乡1号; 农家品种中排在前10位的依次是小三变、然谷、昆昌谷、十尖子、盖平白、红耙糯、压塌车、小金俊、赶牛鞭和大谷子。相关分析(表6)表明, F值与15个表型性状的相关性均达极显著水平, 与穗颈形状和穗下节间长呈极显著负相关。利用综合得分F值和表型性状构建最优回归方程, 筛选谷子资源综合性状评价指标。通过逐步回归分析构建的最优回归方程为y= (155.030+7.845x1+9.220x3+9.660x5+16.537x6+0.769x7+2.017x8+ 86.518x10+ 2.119x14)× 10-3, 式中x1、x3、x5、x6、x7、x8、x10和x14分别代表叶鞘色、刚毛长度、粒色、米色、株高、穗长、茎粗和单穗粒重8个表型性状, 方程相关系数r和决定系数R2分别为0.994和0.987, 表明这8个自变量可决定F值总变异的97.1%, F值为8364.041, 方程极显著。由回归方程可知, 在15个表型性状中, 叶鞘色、刚毛长度、粒色、米色、株高、穗长、茎粗和单穗粒重8个性状对谷子种质资源表型性状多样性综合值的影响显著, 可以作为核心种质综合评价指标。
| 表6 15个表型性状与表型综合值(F值)间的相关系数 Table 6 Correlation coefficients between 15 phenotypic traits and comprehensive value (F-value) |
图2表明, 欧式距离在11.07时, 将不同国家来源的谷子资源划分为三类; 第I类包含部分欧洲(波兰、德国、俄罗斯、乌克兰、前南斯拉夫、荷兰)、部分亚洲(阿富汗、吉尔吉斯坦、乌兹别克斯坦、黎巴嫩)和非洲(摩洛哥)的品种; 第II类由部分亚洲(印度尼西亚、哈萨克斯坦和伊朗)、部分欧洲(匈牙利、瑞士、保加利亚)、非洲(肯尼亚、南非)、北美洲(美国、加拿大)的品种组成; 第III类有部分亚洲(中国、蒙古、朝鲜、日本、印度、格鲁吉亚、菲律宾、尼泊尔、孟加拉国、土耳其)以及非洲(埃塞俄比亚)和欧洲(法国)的品种组成。
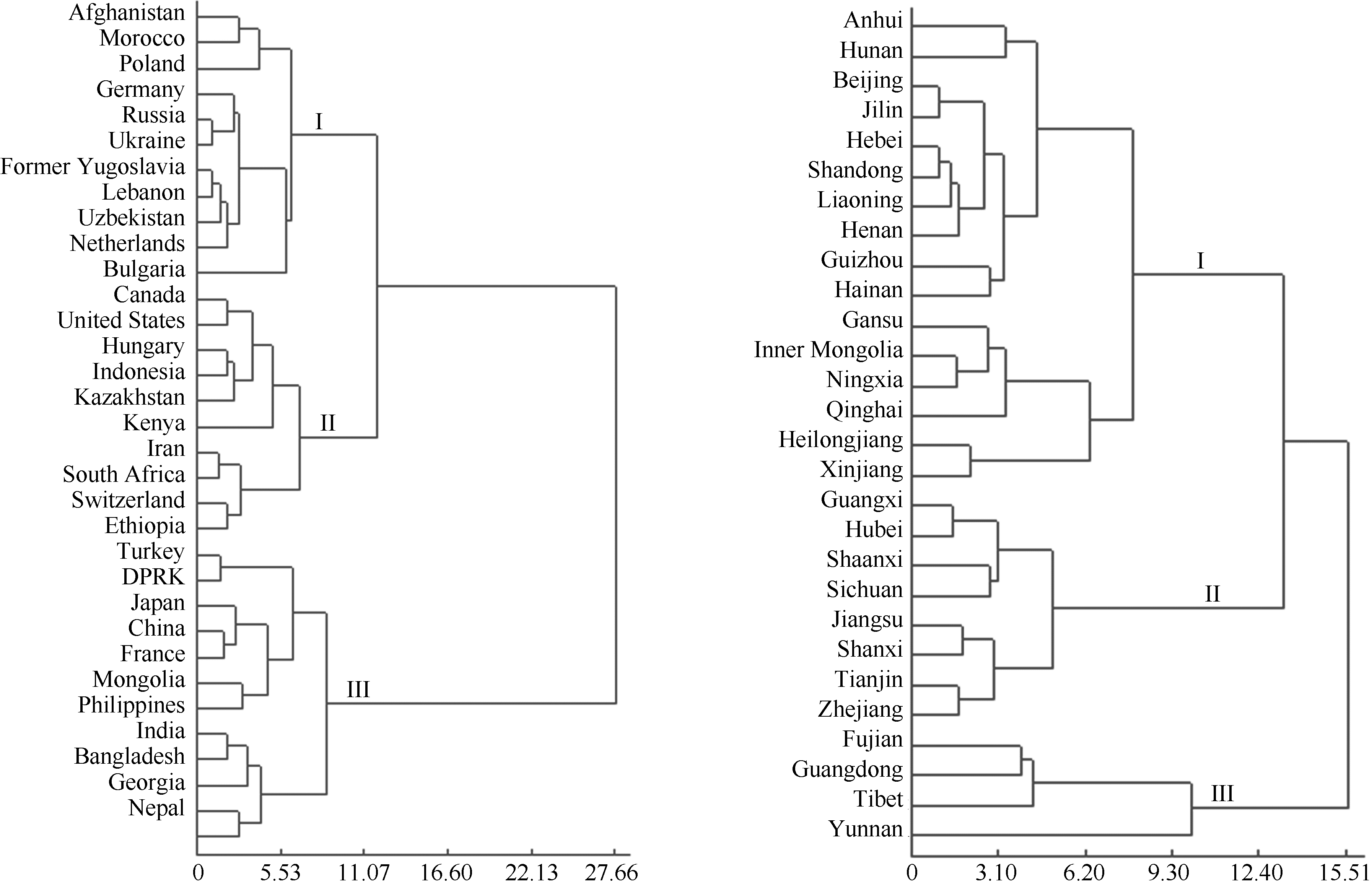 | 图2 不同国家和中国不同省份来源的谷子种质的系统聚类图Fig. 2 Clustering dendrogram of foxtail millet germplasm from different countries and provinces of China |
对我国谷子资源按不同省份来源聚类发现, 欧氏距离在10.35时, 我国谷子资源划分为3个类群; 第I类包括吉林、辽宁、河北、北京、山东、河南、贵州、海南、安徽、湖南、内蒙古、甘肃、宁夏、青海、黑龙江、新疆等省区的品种; 第II类主要包含山西、陕西、天津、四川、广西、湖北、江苏、浙江的品种; 第III类包含广东、福建、西藏和云南的品种。
本研究中核心种质9个数量性状均表现丰富的变异, 各性状的变幅和变异系数均很高, 远高于田伯红等[20]在河北沧州对我国部分农家品种和育成品种表型鉴定的报道, 也高于王晓娟等[21]、孙加梅等[26]和杨天育等[27]对地方品种的遗传多样性的分析结果。其中, 单穗粒重和单穗重2个产量紧密相关性状的变异系数分别为43.41和39.58, 遗传多样性指数分别为1.82和1.84, 其他表型性状也表现了高的变异系数和遗传多样性指数, 说明本研究中的应用核心种质变异丰富, 不仅较好地代表了谷子资源的遗传多样性, 在遗传育种中也有较好的利用价值。稳定性是种质资源的一个重要构成要素, 虽然多数数量性状的平均值2012年的结果大于2013年的结果, 且年度间的变异系数有一定变化, 但表现相同的趋势(表3), 说明在山西东阳生态环境下不同年度之间的结果是稳定的。年度间变化较为明显的是穗下节间长度、主穗直径、单穗粒重和单穗重, 说明这些性状可能受多个微效基因控制, 遗传基础比较复杂。
本研究中农家品种涉及我国所有生态区, 育成品种主要来自华北、西北和东北谷子主产区。从表2对农家品种和育成品种的比较分析可以看出, 虽然育成品种的变异系数受育种选择瓶颈效应影响而低于农家品种, 但育成品种内的多样性仍很高, 还存在着广泛的变异供育种选择, 如育成品种的单穗重的变幅和变异系数分别为1.50~34.95 g和29.08, 单穗粒重的变幅和变异系数分别为1.20~29.35 g和32.24, 其他几个产量相关的数量性状也存在广泛变异, 这说明谷子育种的选择压力还不够强。从育成品种和农家品种比较来看, 现有育成品种的株高较农家品种降低了4.32 cm, 穗长减少了1.60 cm, 生育期平均增加了2.70 d, 单穗重和单穗粒重2个产量相关性状别提高了3.61 g和3.12 g, 这些育种进步所体现的是遗传上较低的选择强度。本研究中的国外种质表现了丰富的表型变异, 这为利用国外材料改良我国谷子品种提供了基因基础和选择空间。综合这些结果可较清楚看出, 我国谷子的育种水平还较低, 育种选择和进步的空间还很大, 培育出更多有突破性品种的可能性很大, 其难度要远低于水稻、小麦等选择瓶颈已很大的主要农作物。当然, 本研究以全国各生态区的品种在一个环境下种植, 是多种品种类型综合的结果, 对特定生态区的农家品种和育成品种在其自身的环境下的比较分析工作正在进行中。
种质资源评价关系到遗传育种的直接利用。目前, 对作物种质资源表型性状综合评价已有多个报道[10, 31, 32], 其中基于表型数据标准化处理和主成分得分归一化处理的F值分析法应用较多[10, 33, 34]。本研究利用这种方法建立了谷子资源的表型综合性状评价体系, 得到了每份资源的F值, 这为育种和遗传研究提供了参考基础。根据F值的结果, 育成品种中来自山西的长生08、长农38、晋谷24、晋谷11、长生07、长生04、晋谷31、长农36、太选10号、晋谷30、晋谷21、晋谷46等在山西综合表现最好, 说明本方法评价结果稳定可靠。本研究中鉴定出的其他高F值的资源应该引起育种家的关注。
在与F值相关达到极显著的15个性状中, 质量性状粒色和米色与F值的相关系数分别达到了0.6660和0.6924; 而在数量性状中株高、主茎节数、穗长、单穗重和单穗粒重的相关系数相对较高。说明这几个性状对资源评价较为重要, 在谷子资源性状鉴定和育种实践中应注重对这些性状的选择。
本研究的性状相关分析建立在大样本和连续两年数据的基础上, 根据统计学原理, 有的相关系数较小, 但相关仍显著或者极显著(表4)。穗颈形状、穗型、刚毛长度、叶鞘色、粒色和米色6个质量性状中, 米色和粒色的相关极显著, 相关系数达0.7032, 说明通过籽粒色泽的选择可以有效选择米色, 这对谷子品质育种有着重要意义。尽管质量性状与产量关联的数量性状间也存在显著相关, 但相关系数均较小, 实际应用意义还有待实践验证。在9个数量性状之间, 单穗重和单穗粒重之间的相关系数高达0.9837, 是所有相关中最高的; 数量性状之间的相关系数多数要高于该数量性状和质量性状之间的相关系数, 且多数为极显著, 说明了这些性状紧密的关联性, 遗传的复杂性和育种选择的困难性。
以来自于一个国家的种质资源作为一个整体计算F值, 世界各地的资源按照本研究的表型性状划分3类(图2), 第I类除中亚的阿富汗外均是东北欧国家, 第II类除保加利亚和匈牙利外, 均为北美和非洲的品种; 第III类以东亚、南亚的品种为主, 但包括了个别非洲和欧洲的国家。从总体上来说, 这个分类在一定程度上反映了品种的地理生态类型。
利用SSR分子标记Wang等[35]等将谷子的农家品种清楚地分为早春播型、春播型、春夏兼播型和南方型4种类型; Jia等[36]等采用SSR标记将谷子育成品种清楚地分为春播型和夏播型两类。本研究按照在山西东阳的表型性状将中国谷子资源分为3类, 这3类和地理来源有着较好的吻合。第I类主要是河南、河北、山东等华北夏谷区的品种和来自黑龙江、内蒙古、宁夏等地区的品种, 这类品种和SSR分子标记分析的农家品种的春夏兼播型, 以及育成品种的夏播型相当; 但海南的品种在本研究中也分到此组。第II类来源较乱, 包括江苏以南地区和山西、陕西的品种, 此组品种在SSR分析中相当于部分南方型和部分春播型。第III类是福建、广东、云南和西藏的品种, 与SSR分析的南方型相当。
表型性状受环境影响大, 利用表型性状分类容易存在各种问题。本研究中从整体上来说, 利用表型性状的分类和地理来源有较清楚的吻合, 这一方面说明谷子品种有着较清楚的地理分化, 也说明本研究连续两年的鉴定结果可靠。将SSR分子标记分类和本研究的表型分类进行比较, 分子标记分类可能更清楚些, 但也容易遗漏一些品种多样性信息, 如SSR标记分类的南方型品种在本研究中出现了明显的分化, 福建、广东、云南和西藏的品种与其他南方地区的品种很清楚地在表型上分开。但分子标记的基因组分析未能表明这一点。从资源深入研究的角度出发, 对资源进行表型和分子相结合的分析是必要的。
表型数据的完整是种质资源精准鉴定的基础。2012和2013两年在山西省农业科学院东阳试验基地对878份谷子资源种植, 这些材料尽管来自于世界各地和我国的各个生态环境, 但均能够在东阳正常成熟, 没有发现极端不抽穗或者不能成熟的材料。这说明东阳的生态环境在谷子资源繁种方面是适合的, 也说明在这个生态环境下能够获得较全面且稳定的表型数据。谷子属光温敏感作物, 地理来源差异大的品种在异地不能抽穗和成熟是资源研究中常见的现象, 特别是南方来源的品种在北方条件下不能抽穗或成熟[37]。山西东阳生态环境为不同类型品种提供了均能成熟的条件, 这对谷子资源研究具有重要的意义。
山西省是我国谷子主产区之一, 是谷子起源中心所在地[38], 本研究完成了对谷子应用核心种质资源的综合评价, 其结果首先对山西利用应用核心种质开展谷子资源深入研究和育种实践有直接指导意义, 其次对其他生态区利用谷子核心种质也提供了表型多样性参数, 对促进谷子遗传育种研究起重要作用。
建立了回归方程, 筛选出8个性状作为谷子表型鉴定的主要指标, 能有效反映谷子种质的综合表现。878份谷子的应用核心种质变异丰富, 在遗传育种中有较好的利用价值。我国谷子的育种水平还较低, 育种选择和进步的空间还很大。山西省适合谷子表型精准鉴定, 鉴定数据全面稳定且可为应用核心种质在山西的应用以及其他谷子产区的育种提供参考。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|