



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
聚合稻瘟病、白叶枯病和褐飞虱抗性基因的三系恢复系改良效果的评价
[楼珏1 , 杨文清1 , 李仲惺2 , 罗天宽1 , 谢永楚1 , 郑国楚1 , 岳高红1 , 徐建龙3 , 卢华金1, *  ]
]
]
|
|


第一作者联系方式: E-mail: yyljlu@wzvcst.edu.cn
结合分子标记辅助轮回选择和田间鉴定的方法, 将三黄占2号的抗稻瘟病基因 Pi-GD-1(t)和 Pi-GD-2(t)(分别简称G1和G2)、CBB23中的抗白叶枯病基因 Xa23 (简称X)和IR65482-7-216-1-2-B (简称IR65482)的抗褐飞虱基因 Bph18(t)(简称B)导入温恢845、温恢117和温恢143等3个中籼恢复系, 获得了8个兼抗稻瘟病和褐飞虱聚合系, 温恢845-G1-G2-B-4、温恢845-G1-G2-B-5、温恢117-G1-G2-X-B-3、温恢143-G1-G2-B-3、温恢143-G2-X-B-9、温恢143-G2-X-B-10、温恢143-G1-G2-B-11和温恢143-G1-G2-B-37。这些聚合系及其与不育系五丰A的测交种, 对稻瘟病和褐飞虱的抗性水平接近或略低于稻瘟病抗性亲本三黄占2号和稻飞虱抗性亲本IR65482。部分改良恢复系如温恢117-G1-G2-X-B-3、温恢143-G2-X-B-9和温恢143-G2-X-B-10及其测交种对白叶枯病表现为抗病或中抗。改良恢复系及其测交种在正常条件下的农艺性状与原始恢复系及其测交种相仿或更优, 具有生产应用价值。研究结果表明, Xa23在不同恢复系背景下抗性表达完全, 而 Pi-GD-1(t)、 Pi-GD-2(t)和 Bph18(t)对稻瘟病和褐飞虱抗性的改良效果与恢复系的遗传背景有关。
The novel blast resistance genes, Pi-GD-1(t) and Pi-GD-2(t)(abbreviated as G1 and G2, respectively) from rice cultivar Sanhuangzhan2, bacterial blight resistance gene Xa23 (abbreviated as X) from CBB23 and brown planthopper resistance gene Bph18(t)(abbreviated as B) from IR65482-7-216-1-2-B (abbreviated as IR65482) were introgressed into three rice restorer lines, Wenhui 845, Wenhui 117 and Wenhui 143, by marker-assisted backcross and pyramiding breeding methods integrated with artificial inoculation, natural induction and phenotypic selections. The eight resistant restorer lines pyramiding blast and brown planthopper resistance genes, i.e. Wenhui 845-G1-G2-B-4, Wenhui 845-G1-G2-B-5, Wenhui 117-G1-G2-X-B-3, Wenhui 143-G1-G2-B-3, Wenhui 143-G2-X-B-9, Wenhui 143-G2-X-B-10, Wenhui 143-G1-G2-B-11, and Wenhui 143-G1-G2-B-37, and their tester lines crossed with the sterile line Wufeng A demonstrated the similar or slight lower level of resistance against rice blast and brown planthopper as compared with the resistant donor parents Sanhuangzhan 2 or IR65482. Part of developed restorer lines (Wenhui 117-G1-G2-X-B-3, Wenhui 143-G2-X-B-9, and Wenhui 143-G2-X-B-10) and their tester lines showed resistance or moderate resistance to bacterial blight. The newly developed restorers resistant to blast, bacterial blight and brown planthopper were similar or superior to their respective original types in agronomic traits under normal condition, implying these resistant restorer lines can be useful in hybrid rice breeding and production. The results indicated that Xa23 could completely express its dominant resistance against different restorer genetic backgrounds whereas effects of resistance improvement in Pi-GD-1(t), Pi-GD-2(t), and Bph18(t) depend on their genetic background.
稻瘟病、白叶枯病、褐飞虱是水稻生产上主要病虫害, 分别由稻瘟病菌(Pyricularia grisea Sacc.有性世代为Magnaporthe grisea)、水稻黄单胞菌水稻致病变种(Xanthomonas oryzae pv. oryzae, Xoo)和Nilaparvala lugens Stal.引起, 每年在全球各个水稻生产国或地区都有不同程度的发生, 严重时甚至会导致绝收[1, 2, 3, 4], 极大地威胁着粮食安全, 还会导致稻米品质下降。
根据农业部全国农业技术推广服务中心提供的杂交水稻的统计资料[5], 2011年全国年推广面积在6.7× 103 hm2以上的杂交稻组合499个, 有抗性表现现的杂交稻组合共计485个, 其中抗稻瘟病杂交稻组合266个, 占全国大面积种植杂交稻的54.85%; 抗白叶枯病杂交稻组合198个, 占40.82%; 抗褐飞虱杂交稻组合18个, 占3.71%; 抗白背飞虱杂交稻组合6个, 占1.24%; 兼抗稻瘟病和白叶枯病的102个, 占21.30%; 兼抗稻瘟病和稻飞虱的7个, 占1.44%; 兼抗白叶枯病和稻飞虱的12个, 占2.47%; 兼抗3种抗性的3个, 仅占0.62%。可见, 目前我国种植的杂交稻组合抗病虫性普遍不高。由于稻瘟病和白叶枯病生理小种的变异, 褐飞虱生物型的改变, 导致抗病品种的小种专化抗性容易丧失[6, 7, 8], 引起病虫害的暴发流行。水稻生产因病虫危害造成大幅减产的事件时有报道, 如何提高水稻品种抗性的持久性和广谱性, 延长品种使用寿命是育种工作者急需解决的问题。
目前, 已报道80多个稻瘟病抗性基因, 并鉴定出众多广谱或持久抗稻瘟病的抗源, 如Moroberekan、Dourode Perecose、0S6、IR36、IR42、Tetep、三黄占2号、地谷、谷梅2号和湘资3150等[9]。广东省农业科学院育成的三黄占2号不仅对国内稻瘟病菌株表现现广谱、稳定、持久的抗性, 而且对菲律宾稻瘟病病菌也达到98%的抗性频率[10]。经鉴定, 三黄占2号携带Pi-GD1(t)、Pi-GD2(t)和Pi-GD3(t) 3个稻瘟病抗性主基因, 以及位于第2和第7染色体上的3个抗稻瘟病QTL[11]。多个质量和数量抗性基因的聚集确保了三黄占2号在生产上十多年稳定持久的稻瘟病抗性。至少有35个水稻抗白叶枯病基因被发掘和鉴定[12], 其中Xa23是从普通野生稻中鉴定的一个在全生育期表现现广谱、高抗白叶枯病的基因, 位于第11染色体SSR标记RM206和RAPD标记RpdH5的区间内, 与2个标记间的遗传距离分别为1.9 cM和7.0 cM, 该抗性基因转移性强[13], 具有广阔的应用前景。已发掘和鉴定了27个抗褐飞虱的主效基因[14, 15], 其中Bph18(t)基因是一个广谱、高抗的显性主效基因, 高抗我国褐飞虱生物型I和生物型II种群[16], 该基因已被Jena等定位在水稻第12染色体上, 并设计了与之紧密连锁的分子标记[17]。
本研究通过分子标记辅助选择, 回交育种和田间鉴定, 将抗稻瘟病基因Pi-GD-1(t)、Pi-GD-2(t)和Pi-GD-3(t), 抗白叶枯病基因Xa23以及抗褐飞虱基因Bph18(t)聚合到中籼恢复系温恢845、温恢117和温恢143中, 选育带有抗性基因且农艺性状优良的改良恢复系, 评价各抗性基因的抗性改良效果, 以期为杂交稻抗病虫育种提供理论依据。
抗性供体亲本为携带抗稻瘟病主基因Pi-GD-1(t)、Pi-GD-2(t)和Pi-GD-3(t)(分别简称G1、G2和G3)的三黄占2号, 携带抗白叶枯病基因Xa23(简称X)的CBB23, 以及携带抗褐飞虱基因Bph18(t)(简称B)的IR65482-7-216-1-2-B (简称IR65482)。受体亲本温恢845、温恢117和温恢143是温州市农业科学研究院育成的中籼三系恢复系。选择细胞质雄性不育系五丰A与原始恢复系及抗性改良恢复系配制测交种, 进行杂种优势和抗性评价。原丰早、IR24和台中本地1号(TN1)分别作为稻瘟病、白叶枯病和褐飞虱接种评价的对照品种。
水稻苗瘟分菌株接种和抗性评价的稻瘟病菌株共有16个, 包括7群15个小种(2012-2B1, 2005-20- 1B1, 2011-44B3, 2009-30-1B11, 2012-43B13, 2010-45B25, 2011-91B29, 2007-90C13, 2011-213C15, 2009-312A49, 2012-105-1D1, 2009-104-2D3, 2007-034-4E1, 2007-234E3, 2013-13F1, 2012-067G1), 叶苗瘟混合接种的稻瘟病菌株为13-98 (ZA1)、13-46 (ZA9)、12-217 (ZB9)、10- 151 (ZC1)、13-76 (ZC11), 皆是我国主要的致病菌株。
水稻白叶枯病接种和抗性评价的菌株C5 (GD1358)是我国V型致病力较强的菌株。
三黄占2号、CBB23和IR65482分别与温恢845、温恢117和温恢143杂交得到F1, 然后选择供体亲本不同而受体亲本相同的F1彼此复交, 复交F1植株与第3个供体亲本和相同受体亲本的F1再次复交, 复交后代选株与轮回亲本回交, 通过连续回交与自交, 聚合抗稻瘟病、抗白叶枯病和抗褐飞虱基因于同一恢复系中(图1)。在每轮复交和回交世代, 苗期分单株提取DNA, 利用与抗性基因紧密连锁的分子标记(表现1), 进行PCR扩增检测抗性基因, 结合表现型目测, 筛选携带目标抗性基因且与轮回亲本表现型接近的个体进行后续的回交或自交。结合综合农艺性状选择和分子标记辅助选择, 最终选育出农艺性状与轮回亲本相似的稳定抗性改良恢复系。
 | 图1 分子改良恢复系稻瘟病、白叶枯病和褐飞虱抗性的图示Fig. 1 Sketch of molecular improvement of blast, bacterial leaf blight, and brown planthopper resistance for restorer lines |
| 表1 稻瘟病、白叶枯病和褐飞虱抗性基因紧密连锁的分子标记信息 Table 1 Information of molecular markers associated with resistance genes for rice blast, bacterial leaf blight, and brown planthopper |
2014年夏, 对原始恢复系和改良恢复系分别于浙江省农业科学院植物保护与微生物研究所开展苗叶瘟的混合接种鉴定和16个菌株人工接种鉴定; 于稻瘟病重发区浙江省文成县三源村开展自然诱发穗瘟的抗性鉴定。参照倪大虎等的方法进行苗瘟的混合接种和单个菌株接种[18]。将试验品种(系)和对照原丰早、三黄占2号的种子浸种催芽后, 按顺序均匀播种在带孔、装有细土、穴间隔3 cm的塑料盘中, 每个品种(系) 10~15粒。供试品种(系)长至三叶一心时, 用弥雾喷雾器将浓度约为2× 105个孢子 mL-1的稻瘟病菌(Pyricularia oryzea)分生孢子悬浮液喷洒于秧苗上, 接种量为每百株苗30 mL悬浮液。接种后将供试秧苗置网室内, 用黑色塑料布遮光保湿24 h, 然后去除遮光条件, 在遮阴保湿条件下培养10 d, 待感病对照品种原丰早稻瘟病情稳定时, 根据目测法调查每个品种(系)所有植株的病斑大小, 取平均值。根据国家标准GB/T 15790-2009将抗性分级指标划分为9级, 1~3级分别为HR、R和MR, 4~5级为MS, 6~7级为S, 8~9级为HS[19]。试验设2次重复。苗叶瘟单个菌株接种鉴定中, 按照MR以上为抗病, 其他为感病计算抗性频率。穗瘟鉴定试验以感病对照原丰早作为诱发品种, 以原丰早和三黄占2号为对照, 每个品种(系)材料种植2行, 每行7株, 行株距20 cm × 17 cm, 设2次重复。黄熟期对2行中间的6株进行穗瘟调查, 参照国家标准GB/T 15790-2009评价穗瘟的抗性, 将抗感反应型划分为高抗、抗、中抗、中感、感和高感6级[19]。
委托中国农业科学院作物科学研究所完成原始恢复系和白叶枯抗性改良恢复系及其测交种的白叶枯病抗性鉴定。2014年夏, 将供试材料播种于中国农业科学院作物科学研究所水槽, 每份材料种植2行, 每行种6株, 行株距为20 cm× 17 cm, 设2次重复。待孕穗期采用剪叶法[18], 接种中国强致病力白叶枯病菌系C5, 菌液浓度为5× 108 mL-1, 接种每个品种(系)中间的4株, 每株从叶尖剪去1~2 cm, 每株接种2~3片叶。接种20 d后, 当感病对照IR24发病情况稳定后, 调查植株的抗性表现现。根据4株接种植株上病斑占叶面积百分率的平均值划分抗性级别: 剪口处无明显病斑的为高抗(HR); ≤ 10%为抗(R); 11%~20%为中抗(MR); 21%~40%为中感(MS); 41%~ 75%为感病(S); ≥ 75%为高感(HS)。
在中国水稻研究所开展褐飞虱抗性鉴定, 参照刘光杰等[20]的方法, 将催芽后的原始恢复系和抗性改良恢复系及其测交种, 以及感虫对照品种TN1按照行长20 cm, 行距5 cm, 每行20粒的规格播种在塑料盆内, 设置2次重复。当秧苗长至二至三叶时, 向每株秧苗平均接入5~7头1~2龄的褐飞虱若虫。当感虫对照品种TN1的死苗率达到95%时, 根据各供试材料的死苗率评定抗性级别[20], 即死苗率≤ 1.0%为0级; 1.1%~10.0%为1级; 10.1%~30.0%为3级; 30.1%~ 50.0%为5级; 50.1%~70.0%为7级; > 70.0%为9级。取2次重复的平均值作为鉴定结果。
2014年夏, 在温州市农业科学研究院藤桥基地对原始恢复系和抗性改良恢复系及其测交种开展综合农艺性状评价。采用2次重复的随机区组试验, 每个材料种植3行, 每行10株, 株行距为30.0 cm× 23.3 cm, 外设保护行。待到成熟时取每份材料中间的5株测定株高和有效穗数, 从中间5株上取2个主穗考察穗长、每穗总粒数、每穗实粒数、结实率和千粒重等性状, 混收中间5株计算单株产量[21]。用SPSS软件处理和分析所得数据。
利用与Pi-GD-1(t)、Pi-GD-2(t)、Pi-GD-3(t)、Xa23和Bph18(t)紧密连锁的分子标记(表现1)分析供体亲本与3个恢复系之间的标记多态性, 除与抗稻瘟病基因Pi-GD-3(t)紧密连锁的分子标记RM179在3个受体亲本与抗源供体间均不存在多态性外, 其他标记都具有良好的多态性并易于开展分子标记检测抗性基因。
在选育抗性改良恢复系的过程中(图1), 从复交F1世代至BC4F2, 每一代都进行分子标记辅助选择, 筛选携带目标抗性基因且农艺性状接近轮回亲本的单株进行后续的回交或自交(图2)。以温恢845抗性改良为例, 温恢845分别与三黄占2号、CBB23和IR65482杂交获得F1, 将温恢845/三黄占2号F1植株与温恢845/CBB23 F1复交, 得到复交群体620株, 利用与Pi-GD1(t)、Pi-GD2(t)和Xa23紧密连锁的分子标记RM6208、RM3855和RM206检测, 筛选出同时带有3个抗性基因的个体45株, 从中选择农艺性状接近轮回亲本温恢845的个体3株, 分别与温恢845/IR65482 F1植株复交, 得到780个第2次复交群体, 利用分子标记筛选出携带Pi-GD1(t)、Pi-GD2(t)、Xa23和Bph18(t)基因的25个单株, 从中选择综合性状接近温恢845的个体3株, 分别与温恢845回交, 共产生BC1F1群体545株, 后续3次连续回交都针对上述4个抗性基因进行分子检测, 每次选择聚合4个抗性基因且农艺性状近似温恢845的个体3株分别与温恢845回交, 产生下一代回交群体, 其中BC2F1、BC3F1和BC4F1群体分别有446、686和850株, 从BC4F1群体中选择聚合4个抗性基因且农艺性状与温恢845相似的个体3株, 混合种成860株规模的BC4F2群体, 结合抗性基因的分子标记检测, 从中选择不同抗性基因纯合的聚合体种成BC4F3株系, 择优用于与五丰A测定杂种优势。
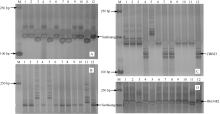
综合分子标记检测结果和各株系的综合农艺性状, 从3个恢复系回交改良后代初步选育出52个BC4F3稳定改良恢复系, 与五丰A配制相应的测交种。进一步对这些改良株系及其测交种进行抗病虫鉴定, 最终选出携带2个或3个抗性基因、农艺性状与轮回亲本相似的改良恢复系8个, 分别命名为温恢845-G1-G2-B-4、温恢845-G1-G2-B-5、温恢117- G1-G2-X-B-3、温恢143-G1-G2-B-3、温恢143-G2-X- B-9、温恢143-G2-X-B-10、温恢143-G1-G2-B-11和温恢143-G1-G2-B-37 (图3)。
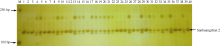
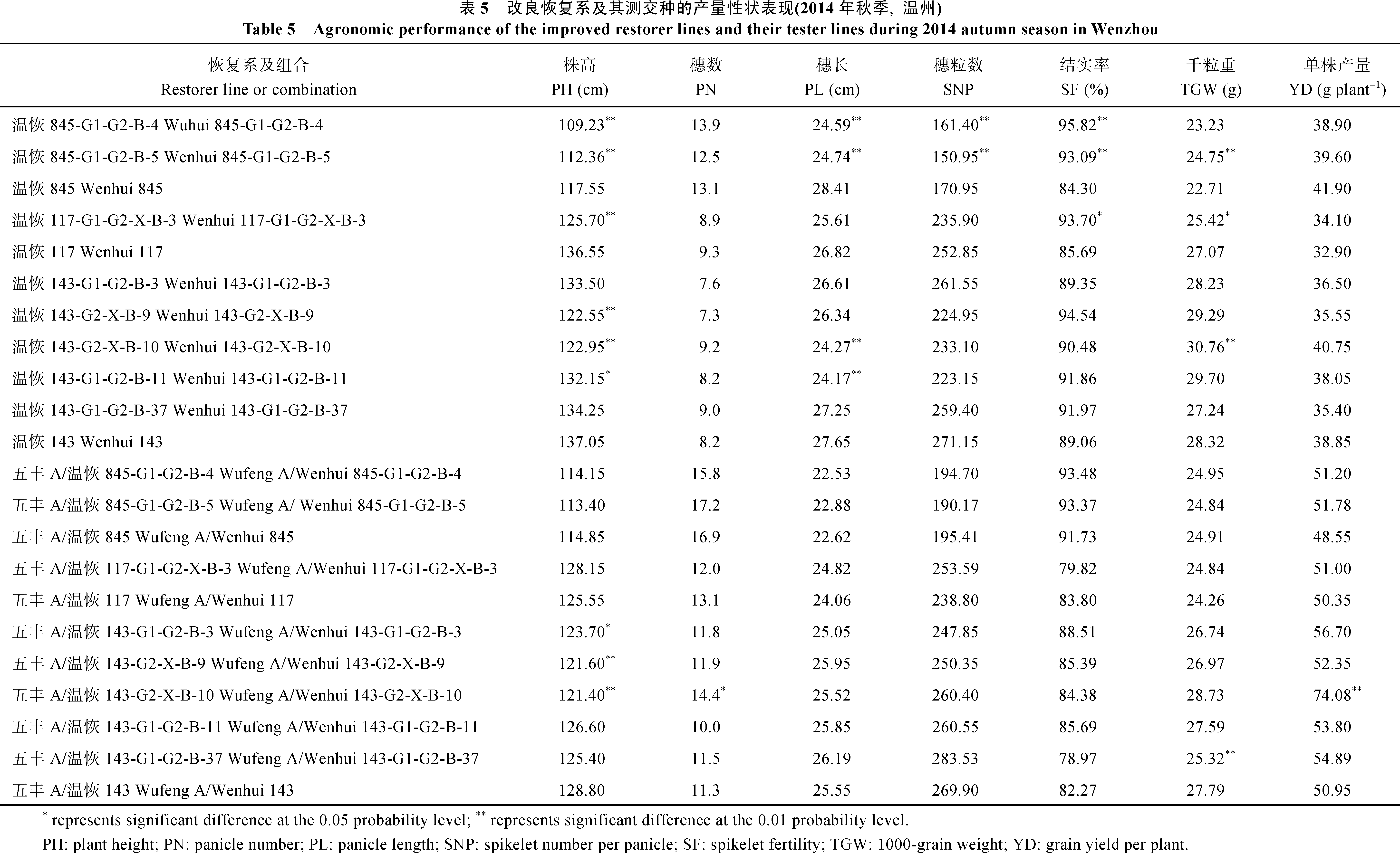
 | 图2 与抗瘟基因Pi-GD-1(t)紧密连锁的RM6208标记在温恢845× 三黄占2号BC4F1群体中PCR扩增结果 M: DL2000 DNA marker; 1~39: BC4F1单株; 40: 三黄占2号。Fig. 2 PCR amplification of the marker RM6208 linked to the blast resistance gene Pi-GD-1(t) in the BC4F1 population derived from Wenhui 845× Sanhuangzhan 2 M: DL2000 DNA marker; 1-39: BC4F1 individual; 40: Sanhuangzhan 2. |
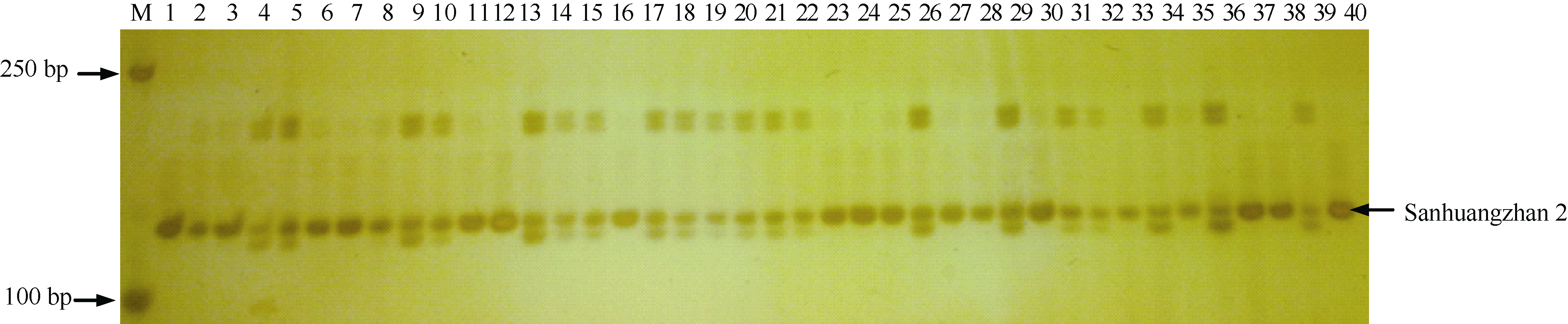
 | 图3 改良恢复系抗性基因分子标记验证 Fig. 3 Confirmation of the resistances for the improved restorer lines using molecular markers |
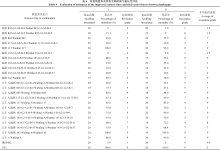
抗性亲本三黄占2号在自然诱发穗瘟和混合接种苗瘟条件下分别表现现抗病和高抗, 分菌株接种除对S14表现现中感外, 其他菌株皆表现现抗性, 抗性频率达到93.75%, 表现现高水平抗性; 感病对照原丰早高感穗瘟, 感苗瘟, 仅对4个稻瘟病菌株有抗性, 抗病频率为25.00%, 表现现高度感病(表现2)。3个恢复系亲本在3种不同稻瘟病鉴定条件下表现现出不同的抗性水平, 温恢117混合接种苗瘟和自然诱发穗瘟分别表现现感病和中抗, 对16个菌株中的15个表现现出抗性, 抗性频率为93.75%; 温恢143中感苗叶瘟, 感穗瘟, 对S2、S7、S10 3个菌株表现现感病, 抗性频率为81.25%; 温恢845感苗叶瘟, 中感穗瘟, 对S1、S2、S6和S15 4个菌株表现现感病, 抗性频率为75.00%。可见, 3个恢复系亲本中, 温恢117的稻瘟病抗性表现现最好, 温恢845最差。
8个多基因聚合恢复系对16个稻瘟病菌株、混合接种以及自然诱发的抗性表现现强于温恢845, 优于或相似于温恢117和温恢143。温恢845的稻瘟病抗性改良效果明显, 苗瘟和穗瘟抗性都得到了较好的改良。8个稻瘟病抗性改良恢复系在自然诱发穗瘟和混合接种苗瘟条件下抗性表现现为中感至抗不等, 其对16种稻瘟病菌株的抗性频率变化范围在75.00%~ 93.75% (表现2)。除温恢117-G1-G2-X-B-3的抗性频率没有变化, 温恢143-G2-X-B-11的抗性频率低于原始亲本温恢143, 苗瘟抗性与亲本相同外, 8个改良恢复系在抗性频率, 苗瘟或穗瘟上都得到了不同程度的改善。
孕穗期接种中国强致病力白叶枯病菌系C5, CBB23表现现抗病, IR24表现现高度感病。温恢117和温恢143表现现感病, 而携带Xa23基因的3个改良恢复系均表现现抗白叶枯病(表现3), 抗性改良效果显著。抗性改良恢复系测交组合的白叶枯病皆表现现为中等抗性, 优于原始亲本测交组合; 其抗性水平之所以达不到抗级, 主要是由于测交的不育系五丰A感白叶枯病。
| 表3 改良恢复系及其测交种孕穗期对白叶枯病菌C5菌系的抗性评价 Table 3 Evaluation of resistance of the improved restorer lines and their tester lines to the virulent Chinese strain C5 of Xanthomonas oryzae pv. oryzae at the booting stage |
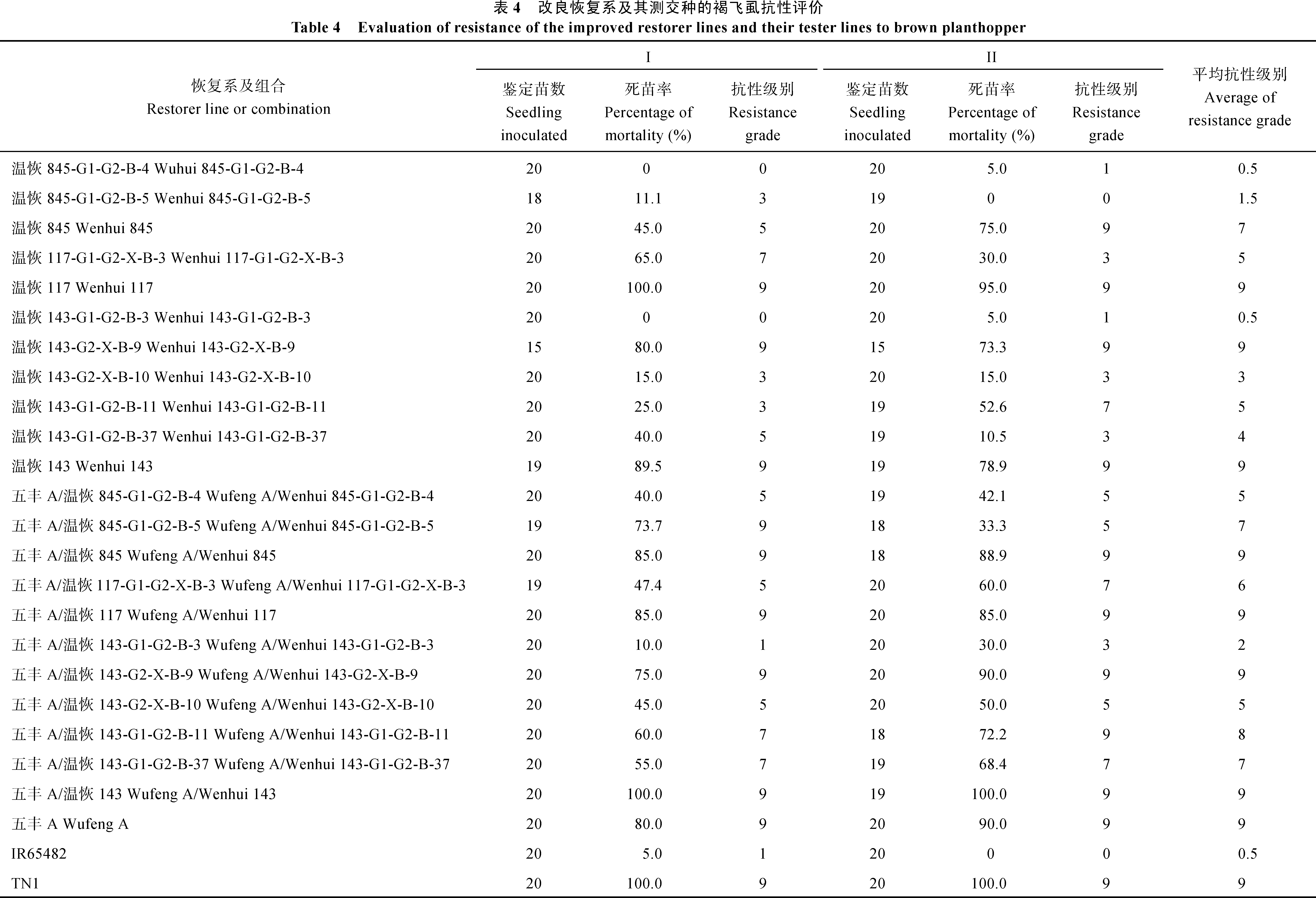
温恢117、温恢143、五丰A和感虫对照品种TN1对褐飞虱的抗性均为9级, 表现现为高感褐飞虱, 温恢845中感褐飞虱, 抗性品种IR65482平均抗性级别为0.5级, 表现现高抗(表现4)。8个抗性改良恢复系中有7个褐飞虱抗性得到了明显提高, 1个与轮回亲本相似, 其中温恢143-G1-G2-B-3抗性最好, 达到0.5级。3个轮回亲本与五丰A配制的测交种的褐飞虱抗性表现现为9级高感, 8个改良恢复系与五丰A的测交种褐飞虱抗性优于或等于原始亲本的测交种但略差于或等于改良恢复系自身, 表现明Bph18(t)基因的导入能提高恢复系的褐飞虱抗性水平, 但改良效果可能与被改良恢复系的遗传背景有关。
与原始恢复系比, 所有改良恢复系的株高、穗长、穗粒数都有所降低, 结实率都得到了提高, 但差异显著性各不相同, 而单株产量与原始恢复系差异不明显。改良恢复系与五丰A测交种的农艺性状与原恢复系与五丰A的测交种的基本相仿, 仅温恢143-G2-X-B-10测交种的单株产量极显著高于原始恢复系的测交种产量(表现5)。
温恢845、温恢117、温恢143是温州市农业科学研究院育成的强三系恢复系, 其配制的多个组合如II优845、宜优845、K优117等通过了浙江省、湖南省和江西省等多地审定[22, 23, 24]。但是温恢845, 温恢117和温恢143对稻瘟病、白叶枯病和褐飞虱的抗性表现现一般, 配制的杂交稻组合的抗性也相对较差。随着推广面积的扩大, 必然会存在病虫害大规模爆发的潜在威胁。因此, 利用转育抗性强、抗谱广的抗性基因改良恢复系, 提高杂交组合抗击病虫害的能力显得尤为重要。本研究通过利用分子标记辅助选择和田间杂交、回交, 将抗性亲本三黄占2号, CBB23和IR65482中的3个抗稻瘟病基因, 抗白叶枯病基因Xa23和抗褐飞虱基因Bph18(t)导入到中籼恢复系温恢117、温恢845和温恢143中, 获得了抗性水平得到改善且农艺性状与轮回亲本基本一致的多个抗性基因聚合恢复系8个, 其与五丰A配制的测交种的白叶枯病抗性和褐飞虱抗性水平都优于原始恢复系与五丰A配制的组合。因此, 本研究得到的8个改良恢复系既是水稻抗病育种新的种质资源, 又具有良好的生产应用价值。
据报道, 三黄占2号中携带了3个抗稻瘟病基因Pi-GD-1(t)、Pi-GD-2(t)和Pi-GD-3(t), 其紧密连锁的分子标记分别为RM6208、RM3855和RM179[11]。本研究中, RM6208和RM3855在抗源三黄占2号和轮回亲本间有较好的多态性差异, 利用分子标记辅助选择鉴定跟踪抗性基因流向, 确保了抗性基因正常导入的可行性和操作性; 但是RM179在恢复系与抗源供体间未能检测到多态性, 因而不能确定三黄占2号所携带的Pi-GD-3(t)基因是否导入改良系。在陈凯等的研究中也发现同样的情况[21]。由于多个抗性基因的聚合涉及到的亲本较多, 分子标记辅助选择的难度和复杂性要比转育单个抗性基因更高。因此, 进一步加强水稻抗性基因的分子标记开发显得十分重要。通过利用已知籼、粳稻全基因组序列的SNP位点设计开发标记, 或者根据克隆基因序列信息开发位于基因内部的功能标记, 确保所开发的标记在不同的抗性差异材料间具有丰富的多态性, 提高选择效率及准确率, 以便能更好地应用于分子标记辅助选择聚合育种, 为抗病虫育种提供有力的技术支持。
从抗性鉴定结果可知, 导入Xa23基因的改良恢复系的白叶枯病抗性水平与抗源CBB23相同, 都表现现为抗病, 说明该基因的白叶枯病抗性表现达彻底, 并不会因为遗传背景的差异影响其抗性表现现, 是一个非常利于分子标记辅助选择育种的白叶枯病抗性基因, 这与Zhang等[25]的研究结果一致。导入Bph18(t)基因后, 在一定程度上提高了改良恢复系的褐飞虱抗性水平, 但未达到抗源供体IR65482的水平, 表现明该基因的抗性表现达较易受到遗传背景的影响, 这与陈凯等的研究结果一致[21]。三黄占2号抗稻瘟病基因的导入, 对温恢845的抗性改良效果显著, 对温恢143的抗性有不同程度的改善, 对温恢117的改善不明显, 这可能与温恢117自身抗性相对较好, 所携带的抗瘟基因与三黄占2号存在一定程度的同质性有关。说明作为优质稻瘟病抗源的三黄占2号可以较好地改良稻瘟病抗性相对较差恢复系的抗瘟性, 但若利用其改良本身抗性水平较高的恢复系, 由于抗瘟基因同质性等原因, 可能存在改良效果不佳的潜在风险。改良恢复系中温恢845-G1-G2-B-4和温恢143- G2-X-B-10的抗性频率要高于温恢845-G1-G2-B-5和温恢143-G2-X-B-9, 稻瘟病综合抗性表现现也更好, 这可能是由于无法用分子标记检测的Pi-GD-3(t)基因在回交过程中被转入前面两个改良恢复系, 提高了其抗性水平, 也可能是因为温恢845和温恢143遗传背景中未知位点与Pi-GD-1(t)或Pi-GD-2(t)基因产生互作效应进而提高了抗性水平, 这些互作位点的定位和效应还有待进一步发掘和验证。分子标记检测结果显示, 温恢143-G1-G2-B-11携带Pi-GD-1(t)和Pi-GD-2(t)基因, 但其稻瘟病抗性频率低于原始亲本, 这可能是在多代回交选育过程中发生小概率的遗传交换事件, 导致了抗性基因的丧失[26]。因此, 在利用分子标记辅助选择进行抗性选育的过程中要适当减少回交次数, 扩大回交群体的容量, 提高农艺性状和抗性兼顾株系入选的几率。
利用回交选育的方法进行恢复系的抗性改良是行之有效的方法, 但是培育出的株系在综合农艺性状及配合力方面与原始亲本表现现相似, 很难实现突破。正如本研究中筛选出的8个抗性改良恢复系, 虽然抗性在不同程度上得到了改良, 但是农艺性状方面基本上与轮回亲本差异不明显。因此, 为了突破原有恢复系的潜力限制可以尝试开展携带抗性基因不同背景恢复系之间的复交, 在分子标记辅助选择目的抗性基因的基础上, 根据分离世代中所产生重组体的综合农艺性状及配合力表现现, 进行抗性与产量等的同步改良。如潘晓飚等利用浙恢7954/三黄占2号的F1与明恢86/IRBB23的F1的复交后代选育的双抗聚合恢复系明浙-G1-G2-G8-Xa23, 即为一个产量和抗性同步改良的苗头恢复系[27]。
需要指出的是, 由于改良恢复系与五丰A配制测交种的种子量有限, 本研究没有开展其稻瘟病抗性评价, 因而对三黄占2号改良杂交组合的稻瘟病抗性还缺少研究数据, 有待今后进一步完善。
利用分子标记辅助选择和回交育种技术, 将三黄占2号抗稻瘟病基因Pi-GD-1(t)和Pi-GD-2(t)、CBB23抗白叶枯病基因Xa23和IR65482抗褐飞虱基因Bph18(t)聚合到温恢845、温恢117和温恢143三个恢复系中, 得到了8个携带2~3个抗性基因的改良恢复系。这些改良恢复系及其与五丰A测交种的稻瘟病和褐飞虱抗性得到了明显的提高, 抗性水平接近或略低于三黄占2号和IR65482。3个白叶枯病改良恢复系的抗性水平与CBB23相同, 表现现抗病; 测交种表现现中抗。在正常条件下, 改良恢复系及其测交种的农艺性状与原始恢复系及其测交种相似或更优, 具有较好的应用前景。
致谢: 感谢浙江省农业科学院植物保护与微生物研究所柴荣耀和郝中娜帮助稻瘟病抗性评价, 中国水稻研究所胡阳协助褐飞虱抗性评价。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|