

{kind=link}
{kind=link}
{kind=link}
{kind=link}
DTA-6对大豆花荚脱落纤维素酶和 GmAC基因表达的调控
[崔洪秋1, 2, *  , 冯乃杰
, 冯乃杰1, * , 孙福东1 , 刘涛1 , 李建英2 , 杜吉到1 , 韩毅强1 , 郑殿峰1, * ]
, 冯乃杰]
|
|


第一作者联系方式: 崔洪秋, E-mail: cuihongqiu@126.com *同等贡献(Contributed equally to this work)
叶面喷施生长调节剂可以调控大豆花荚脱落。2012—2013年在大庆林甸县黑龙江八一农垦大学试验基地采用大田小区试验, R1期对大豆绥农28、垦丰16和合丰50分别叶面喷施DTA-6调节剂, 对调控花荚离区脱落纤维素酶( GmAC)基因相对表达量、荚脱落纤维素酶(AC)活性、花荚脱落率和产量进行了研究。结果表明: DTA-6抑制大豆花荚离区 GmAC基因相对表达量, 最大可达51% (±CK%); 大豆荚的AC活性能够在喷药后阶段性地被降低, 在不同品种上增加和降低幅度存在差异; 能够显著降低花荚脱落率( P<0.05), 显著增加产量( P<0.05)。植物生长调节剂DTA-6是通过降低花荚离区 GmAC基因相对表达量, 调节AC活性, 从而减少植株的花荚脱落率, 以实现对产量的调控。
, FENG Nai-Jie
It is effective to regulate abscission of soybean flowers and pods by spraying plant growth regulators (PGRs). This study was carried out to determine the effect of DTA-6 on abscission cellulase (AC) activity, abscission cellulase gene ( GmAC) expressive, abscission of flowers and pods, and yield in soybean. DTA-6 was foliage sprayed at R1 stage on three varieties of Suinong 28, Kenfeng 16, and Hefeng 50 in 2012 and 2013. The results demonstrated that DTA-6 treatment inhibited GmAC gene expression in abscission zone of flower and pod, with the maximum reduction of 51% among three varieties compared with the control. The abscission cellulase activity was periodically decreased by DTA-6, with different decrements among soybean cultivars. DTA-6 significantly decreased ( P<0.05) abscission rate of soybean flowers and pods, and significantly increased yield ( P<0.05). The relative expression of abscission cellulase gene ( GmAC) in abscission zone of flowers-pods was decreased and the AC activity was regulated by DTA-6, resulting in reduced abscission rate of soybean flowers and pods and promoted yield.
大豆是我国重要的油料和经济作物。大豆花荚脱落率过高是大豆籽粒产量的主要限制因素之一[1, 2], 大豆花荚脱落是大豆生殖生长的一种自我调节现象[3], 与产量密切相关。多数大豆品种的花荚脱落率都较高, 可达40%~70%[4], 个别甚至高达80%以上[5], 品种间存在差异[6]。实验和实践已证明, 通过减少花荚脱落提高大豆籽粒产量的潜力很大[7, 8]。各种农艺措施都将促花保荚、减少脱落作为增加产量的重要途径。试验研究发现植物生长调节剂能够降低花荚脱落率, 化控技术也被作为增产措施, 在大豆生产上越来越受到重视, 不同种类的植物生长调节剂调控花荚脱落率的机制存在差异, 由于不同的大豆品种对调节剂种类和浓度要求较高, 研究DTA-6对大豆花荚增产的调控机制显得尤为重要。
一些学者认为, 大豆花荚的形成或败育主要与光合同化物供应的有效性[9]和植物内源激素的有效性[10, 11]这两个内部生理因素有关。近年来, 关于植物器官脱落的研究已深入到分子生物学水平[12, 13], 有研究认为, 植物器官脱落的生理生化变化主要集中在器官基部的离层区内[14, 15], 而调控脱落的水解酶之一— — 脱落纤维素酶(abscission cellulose, AC), 在植物器官脱落中发挥着重要作用, 其变化主要受脱落纤维素酶基因表达的调控。在调控该基因表达的多种因素中, 外源激素效果显著, 以往关于这方面的报道很多, 但主要集中在番茄、柑橘、菜豆等植物上。关于大豆花荚脱落方面, 柴国华等[16]、李辉亮等[17]、Cheng等[18]学者做过很多基础性研究工作, 主要集中在逆境胁迫及激素对大豆花荚离层细胞基因表达的调节方面, 试验结果多未结合田间产量。冯乃杰等[19]通过几年试验研究, 发现植物生长调节剂能够调控大豆花荚发育、降低花荚脱落率, 对大豆叶片、茎和叶柄的显微结构和亚显微结构均有一定的调节作用, 并且能显著增加产量。Ramesh等[20]也做了生长调节剂对大豆形态和生理生化相关研究, 未见分子方面的研究。目前, 植物器官脱落的研究方向已经从最初的外界环境以及生理生化因素(如内源激素代谢和养分、能量供应等)向分子水平方向(如相关基因表达)转变, 将生理生化指标、分子研究和产量结合在一起的研究较少。
通过课题组几年的试验研究, 发现植物生长调节剂能够调控大豆花荚发育, 降低花荚脱落率, 对大豆叶片、茎和叶柄的显微结构和亚显微结构都有一定的调节作用, 并且能显著增加产量。本研究在R1期叶喷DTA-6, 然后采用分子生物学方法研究主要脱落酶基因表达量, 对生理指标脱落纤维素酶活性进行花后定期动态监测, 再结合田间植株最终花荚脱落结果和产量情况, 分析研究田间环境DTA-6调控花荚、增产的作用机制。大豆花荚器官脱落的分子生物学研究将深化人们对其本质的认识, 控制大豆花荚的脱落程度是植物器官脱落分子生物学基础研究在农业实践中的应用。
试验于2012年和2013年在黑龙江八一农垦大学(大庆市林甸县)大豆试验基地进行, 土壤类型为草甸黑钙土, 土壤肥力中等, 前茬为玉米, 耕层(0~ 20 cm)含有机量3.08%、碱解氮178.50 mg kg-1、有效磷25.4 mg kg-1、速效钾257.4 mg kg-1, pH 7.88, 盐总量 0.1%。
选用东北地区生产上种植面积相对大的和基因型存在差异的主栽亚有限类型大豆品种绥农28 (母本绥农14, 父本绥农10, 生育期120 d, 株高110 cm, 紫花, 长叶, 灰毛), 垦丰16 (母本黑农34× 父本垦农5号, 生育期120 d, 株高65 cm左右, 白花, 披针形叶, 灰毛), 合丰50 (母本合丰35, 父本合95-1101, 生育期120 d, 株高90.3 cm, 紫花, 长叶)。
植物生长调节剂2-N, N-二乙氨基乙基己酸酯(diethyl aminoethyl hexanoate, 简称DTA-6, 为叔胺类活性物质), 由福建浩伦公司生产。
在R1期叶面喷施60 mg L-1 DTA-6 (2012年大豆田间DTA-6浓度筛选确定), 以清水为对照(CK), 药液量为225 L hm-2。2012年5月10日和2013年5月16日机器播种, 出苗后人工间苗, 密度30万株 hm-2, 小区面积为19.50 m2, 4次重复, 随机区组排列。2012年10月7日和2013年10月5日收获。施复合肥(N-P-K, 15-15-15) 450 kg hm-2, 人工除草田间管理同常规。
于R1期喷施调节剂后5 d, 取花荚离区组织。用手术剪刀剥取同处理大豆花荚基部与茎相连离区组织, 长3~4 mm, 迅速放入液氮速冻30 min后, 用锡箔包好置-70℃贮藏, 取样所用器械均经高温灭菌。
采用TRIzol法提取总RNA, 以电泳检测RNA质量后, 利用反转录合成cDNA, 以GmActin为内标进行实时荧光定量PCR, PCR体系(25 μ L)含SYBR Premix Ex Taq II (2× ) 12.5 μ L、上下游引物[21] (表1)各1 μ L、RT产物2 μ L、ddH2O 8.5 μ L, 扩增条件为95℃ 30 s, 95℃ 5 s, 58℃ 60 s, 40个循环; 在72℃延伸时收集荧光信号。药品和仪器均来自与TaKaRa公司。
从R1期喷药后28 d (R4)开始, 每7 d调查一次, 分别在喷药后28 d (R4)、35 d (R5)、42 d (R5)、49 d (R6)、56 d (R6)取样, 直到叶片完全脱落, 样品经液氮速冻后, 于-40℃贮藏。用羧甲基纤维素钠盐(CMC-Na)测定酶活性[22, 23]。
| 表1 实时荧光定量PCR引物序列 Table 1 Primer sequences used in qRT-PCR |
取荚0.5 g, 加入2 mL pH 7.2的磷酸缓冲液, 在0~4℃冰浴中研磨提取, 提取液倒入离心管中, 再分别用1 mL的溶液冲洗研钵2次, 共4 mL, 一并倒入离心管中, 4℃下12 000× g离心20 min, 取上清液4℃保存, 作为纤维素酶粗提液。
取2支试管, 分别加入1 mL酶液, 然后在一支试管中加入1 mL羧甲基纤维素钠(CMC-Na), 在另一支试管中加入1 mL蒸馏水作为对照。将2支试管摇匀后, 在电热恒温培养箱中40℃保温24 h, 采用3, 5-二硝基水杨酸比色法[24], 测定反应体系中还原糖的含量。以每小时生成l mg还原糖作为1个酶活单位[25]。
采用铺纱布网法调查脱落花荚。
花荚脱落率(%) = (脱落花数+脱落荚数)/(脱落花数+脱落荚数+座荚数)× 100
在大豆成熟期, 从每个小区收获0.65 m2 (垄长1.00 m× 垄宽0.65 m), 选长势均匀的10株, 统计株粒数、株有效荚数、单株粒重、百粒重(干)等。
产量(kg hm-2)=单株粒数× 百粒重(g)× 公顷株数/ 100 000。
采用Microsoft Excel 2003制图、SPSS19.0统计分析, 用Duncan’ s法进行多重比较。
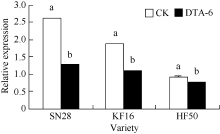
从图1可知, 花荚离区GmAC基因相对表达量, 绥农28 DTA-6处理比CK降低51%, 垦丰16 DTA-6处理比CK降低42%, 合丰50 DTA-6处理比CK降低了17%。说明R1期叶面喷施DTA-6在3个大豆品种中, 均降低了大豆花荚离区GmAC基因相对表达量, 绥农28降低最多, 合丰50降低最少。
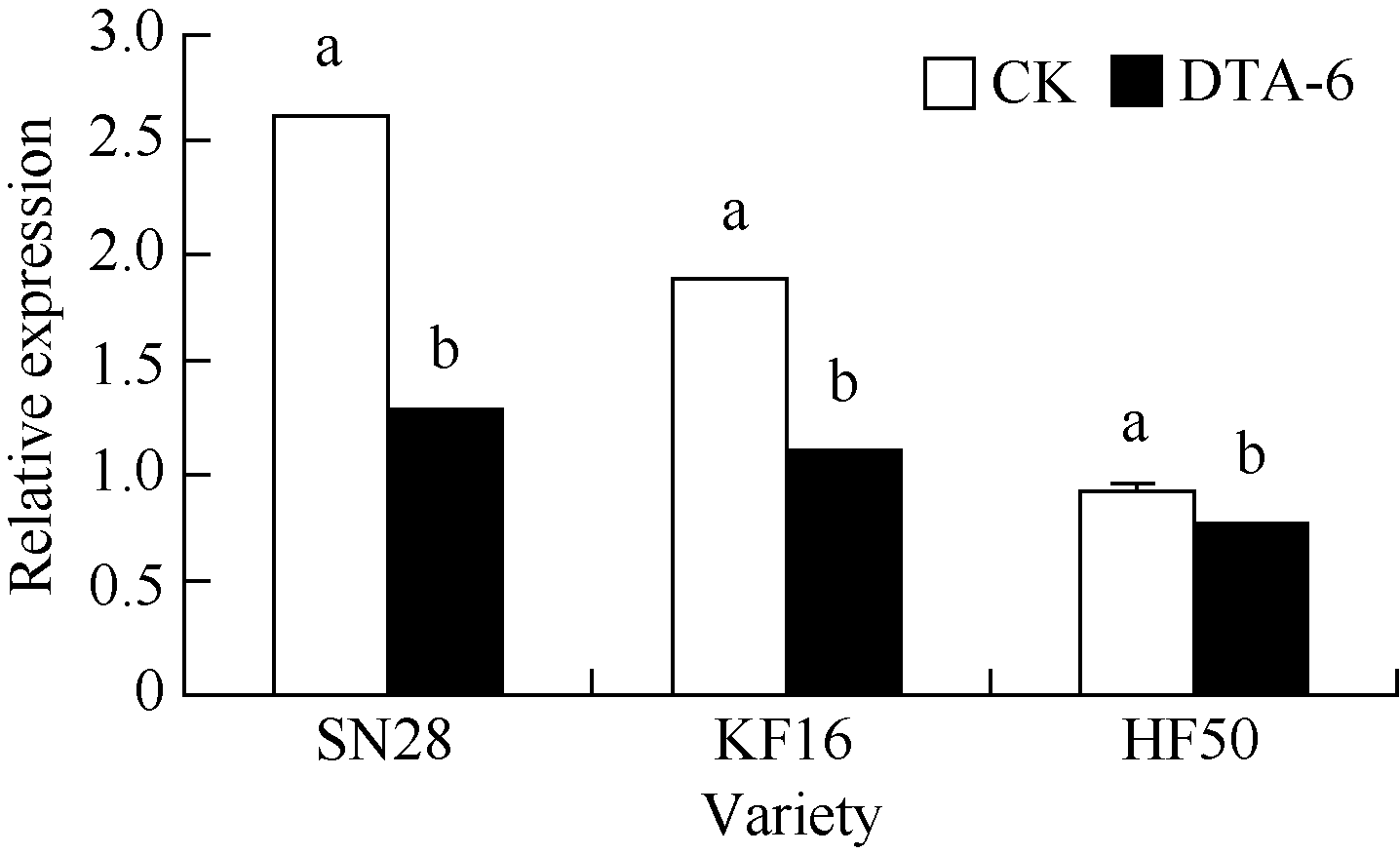 | 图1 R1期叶喷DTA-6对大豆花荚离区GmAC基因相对表达量的影响 图柱顶端不同字母表示处理间在0.05水平上显著。SN28: 绥农28; KF16: 垦丰16; HF50: 合丰50。Fig. 1 Effect of DTA-6 sprayed at R1 stage on GmAC gene expression in soybean Different letters superscripted show significant difference in relative expression between treatments at the 0.05 probability level. SN28: Suinong 28; KF16: Kenfeng 16; HF50: Hefeng 50. |
从图2可知, 绥农28DTA-6处理AC活性在喷药后28、42和49 d高于CK, 其他2个时间均低于CK, 与CK相比随时间递进呈“ 高-低-高-低” 交替; 垦丰16 DTA-6处理, 除了喷药后42 d略高于CK (没有显著差异)外, 其他时间均低于CK, 差异不显著。合丰50 DTA-6处理在喷药后28~42 d荚脱落纤维素酶活性高于CK, 49 d和56 d显著低于CK。
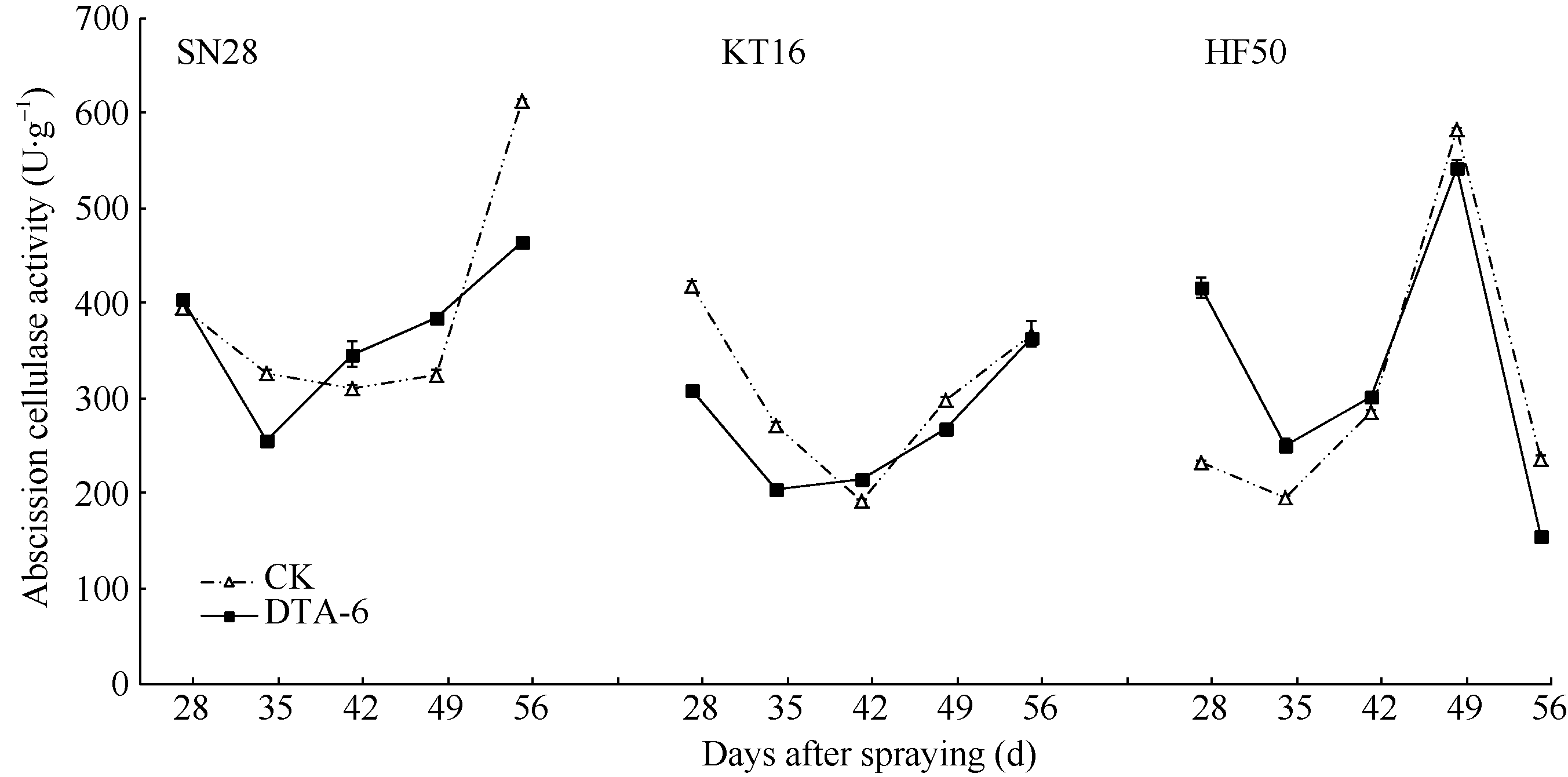 | 图2 R1叶喷DTA-6对大豆荚脱落纤维素酶活性的影响 SN28: 绥农28; KF16: 垦丰16; HF50: 合丰50。Fig. 2 Effect of DTA-6 sprayed at R1 stage on the GmAC activity of soybean pods SN28: Suinong 28; KF16: Kenfeng 16; HF50: Hefeng 50. |
从喷药后42 d为转折点, 之前绥农28、垦丰16两个品种处理高于对照, 之后处理低于对照, 合丰50变化趋势相反。DTA-6对大豆荚AC的调控随着喷药后天数的变化而变化, 品种不同时变化也不尽相同。推测该差异是品种差异造成的。
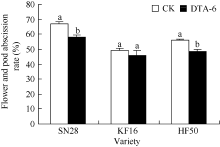
从图3可以看出, 经DTA-6处理的3个品种都表现降低花荚脱落率, 与CK差异显著(P< 0.05)。说明始花期(R1)叶面喷施DTA-6能够有效地降低大豆花荚脱落率。品种间差异与DTA-6对GmAC基因表达的影响相对应, 说明GmAC基因在DTA-6对花荚脱落率的影响中起着重要作用。
 | 图3 R1期叶面喷施DTA-6对花荚脱落率的影响 图柱顶端不同字母表示处理间差异达0.05显著水平。SN28: 绥农28; KF16: 垦丰16; HF50: 合丰50。Fig. 3 Effects of DTA-6 on abscission rate of flowers and pods in soybean Superscripted by different letters are significantly different between treatment at the 0.05 probability level. SN28: Suinong 28; KF16: Kenfeng 16; HF50: Hefeng 50. |
由图4可知, 除2012年垦丰16 DTA-6处理产量没有显著高于CK外, DTA-6处理的3个品种产量均显著(P< 0.05)高于CK, 增产幅度不一致。2年的产量结果均表明, DTA-6对不同基因型亚有限大豆品种产量有显著增产效果。增幅不同是品种差异和年度差异造成的。品种产量变化趋势与DTA-6对GmAC基因表达的影响相对应, 说明GmAC基因虽然对产量没有直接作用, 但可通过影响脱落关键酶影响花荚脱落率, 进而影响产量。
 | 图4 R1期叶面喷施DTA-6对大豆产量的影响 图柱顶端不同字母表示差异达0.05显著水平。SN28: 绥农28; KF16: 垦丰16; HF50: 合丰50。Fig. 4 Effects of DTA-6 sprayed at R1 stage on soybean yield Superscripted by different letters are significantly different between treatments at the 0.05 probability level. SN28: Suinong 28; KF16: Kenfeng 16; HF50: Hefeng 50. |
从表2可知, DTA-6处理比CK增加了单株结荚数, 垦丰16中差异显著(P< 0.05), 其他2个品种中差异不显著。DTA-6处理比CK增加了单株粒数, 绥农28和合丰50中差异显著(P< 0.05), 垦丰16中差异不显著。DTA-6处理比CK增加了百粒重(干), 但在3个品种中差异均不显著。说明R1期叶面喷施DTA-6能够影响产量构成因子, 结荚数和单株粒数在个别品种上显著(P< 0.05)增加, 百粒重差异不显著。
| 表2 R1期叶面喷施DTA-6对大豆产量构成因子的影响 Table 2 Effects of DTA-6 sprayed at R1 stage on yield components of soybean |
在3个大豆品种中, R1期叶面喷施DTA-6均降低了大豆花荚离区GmAC基因相对表达量, 其中绥农28 (51%)降低最多, 与预计的结果相似花荚脱落的减少与大豆花荚离区GmAC基因相对表达量降低的结果是一致, 只是不同的品种上降低的幅度不同。
纤维素酶是细胞壁水解酶之一, 其作用越来越受到重视。利用基因工程克隆植物纤维素酶基因, 通过转基因、反义抑制调控植物的生理效应已有报道[26, 27]。作为调控脱落的水解酶基因, GmAC基因, 在大豆花荚脱落中发挥着重要作用。用14C标记氨基酸的示踪实验结果表明, 脱落发生时, 离层细胞氨基酸和蛋白质的含量增加[28]; 与此同时, 细胞内mRNA和rRNA的含量也增加, 而在ABA诱导的脱落中, RNA和蛋白质能在几小时内迅速增加[29]。这些蛋白质大部分是与脱落有关的酶[30]。说明脱落酶基因表达量与脱落酶活性有着重要的对应关系。有研究报道, 脱落纤维素酶在大豆的叶片、花和荚的脱落层中也存在[31]。柴国华[21]认为, GmAC基因表达存在组织特异性, 离层> 嫩叶> 根> 老叶> 茎, 大豆植株在离层脱落纤维素酶的活性最高, 最容易脱落, 反之, 茎中活性最低, 最不容易脱落, 该结论和Kemmerer等[31]的研究结果类似。Tucker等[32]在研究菜豆时发现, AC基因的组织表达模式, 在离层区及邻近维管束中超量表达, 在上胚轴和茎中表达量很低。因而在本试验中, 我们也选用表达量最高的花荚离区作为材料, 试验结果显示DTA-6处理后在离区检测到了GmAC基因表达, 但表现出的结果是抑制了GmAC基因相对表达量。
柴国华[21]的研究还表明, 最适浓度3种激素(乙烯、ABA、IAA)处理大豆植株, 能显著影响花荚脱落率, 且存在品种差异性, 其变化趋势与各激素对GmAC基因表达的影响相对应, 说明GmAC基因在激素对花荚脱落率的影响中起着重要作用。DTA-6作为植物生长调节剂对大豆花荚的离区脱落纤维素酶基因有调控作用, 结论与前人研究激素作用结果基本相符。首先, DTA-6对大豆脱落纤维素酶基因表达量有抑制作用, 都能降低表达量; 其次, 在不同品种上表现的降低趋势不一致。关于外源植物生长调节剂DTA-6调控GmAC基因表达过程中上下游基因的变化, 内源激素信号传导, 各种物质间的相互作用, 还有待进一步研究。
DTA-6处理的AC和CK比较差异显著(P< 0.05), 随着时间的推移, 变化呈先升高后降低再升高的趋势, 合丰50接近成熟期时生理衰老引起其酶活性快速下降。喷施DTA-6后, AC活性在不同品种间存在差异, 且随着喷药后天数的变化而呈现各自的变化特点。DTA-6处理与CK相比, 在3个品种的脱落纤维素酶变化趋势是在喷药后42 d (R5)较低, 喷药后28 d (R4)和喷药后56 d (R6)较高, 这与宋莉萍 “ 在合丰50上R1期喷DTA-6处理AC活性, 呈先升高后降低再升高的趋势” 部分一致[33]。合丰50在喷药后49 d前与宋莉萍的结论(喷药后50 d)基本一致, 本实验在喷药后56 d, AC大幅降低, 宋莉萍试验中没有体现, 分析其原因可能是合丰50接近成熟期时, 生理衰老引起的酶活性快速下降。
多聚半乳糖醛缩酶(polymerziatoin)和纤维素酶(cellulase)在植物整个生长发育阶段有重要作用, 其变化主要受两类相应基因表达调控。诸多研究表明, 纤维素酶、多聚半乳糖醛酸酶等细胞壁降解酶活性与离区脱落进程有着密切的关系[34, 35]。纤维素酶己被认为是影响菜豆、棉花等叶柄外植体脱落的主要酶, 并受植物激素的调节[36]。Oberholste等[37]观察到用乙烯利处理整体植株和外植体时, 发现大豆花的脱落有细胞学上和超微结构上的不同。如果多种水解酶同工酶在脱落中是需要的, 那么引起脱落的生理条件是相对重要的[38]。更应该注意的是, 基因在自然条件下与诱导脱落中的表达是不一样的。说明脱落是一个由多种酶参加的复杂过程, 在脱落过程中哪种酶起主导作用, 要看脱落发生时具体的生理条件[39]。试验中DTA-6处理和CK的变化不同与“ 基因在自然条件下与诱导脱落中的表达是不一样的” 结果基本相符, 在生理条件方面的研究, 我们还有待进一步的完善。
DTA-6处理的绥农28、垦丰16、合丰50都表现为降低花荚脱落率, 与CK差异显著(P< 0.05)。R1叶面喷施DTA-6能够有效地降低大豆花荚脱落率, 这一结果与柴国华 “ 最适浓度三种激素(乙烯、ABA、IAA)处理大豆植株, 能显著影响花荚脱落率, 且存在品种特异性” 的结论基本一致[21]。说明植物生长调节剂DTA-6与激素乙烯、ABA、IAA处理效果有着相似的规律。品种的变化趋势与DTA-6对GmAC基因表达的影响相对应说明GmAC基因在DTA-6对花荚脱落率的影响中起着重要作用。DTA-6处理的3个品种产量均高于CK, 差异显著(P< 0.05)。表明DTA-6对亚有限大豆品种产量都有显著增产效果。2年的产量结果, 均表明DTA-6对不同基因型亚有限大豆品种产量都有显著的增产效果, 与郑殿峰等研究结果“ 喷施调节剂后, 在不同大豆品种上都有增产的效应, 但是不同大豆品种增产幅度不同” 基本一致[40]。大豆花荚离区GmAC基因相对表达量变化规律与花荚脱落率基本一致, 与产量相反。DTA-6处理在喷药后5 d, 花荚离区GmAC基因相对表达量在不同品种上表现出不同的程度抑制, 喷药28 d后豆荚的AC活性降低, 最终反映在大豆花荚脱落率有效降低、产量显著增加上。
DTA-6通过抑制大豆花荚离区GmAC基因相对表达量, 阶段性地降低大豆荚的AC酶活性, 显著降低花荚脱落率(P< 0.05), 进而显著增加产量(P< 0.05), 并且存在品种差异。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|