{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
光温要素对水稻群体茎蘖增长动态影响的分析及模拟
[王萌萌1, 2 , 杨沈斌1, 2, *  , 江晓东
, 江晓东1, 2 , 王应平3 , 陈德2 , 黄维2 , 于庚康4 , 石春林5 ]
, 江晓东|
|


为研究水稻茎蘖增长阶段光温要素对茎蘖动态的影响, 并验证现有茎蘖动态模拟模型中的光温影响方程, 以籼型两系杂交稻陵两优268和两优培九为试验品种, 进行了为期2年每年7个播期的大田试验。首先, 采用Richards方程对茎蘖观测数据进行拟合, 获取茎蘖增长动态的特征参数。然后, 分析特征参数与茎蘖增长期内平均光温要素和气候要素的关系, 并在此基础上, 以光温组合因子为自变量, 分别构建2个品种茎蘖增长速率和分蘖率的光温组合影响方程, 将获取的方程替换水稻群体茎蘖动态模拟模型中的光温影响方程。最后, 验证和比较替换前后模型的模拟结果及与实测茎蘖动态的误差。结果显示, 受光温要素的共同作用, 平均茎蘖增长速率和最大茎蘖密度均与光温要素显著正相关, 表明光温要素不仅影响茎蘖增长速率, 也影响实际最大群体茎蘖密度, 这在构建的光温组合影响方程中得到了较好的反映。较现有模型, 替换后模型在茎蘖增长动态上的模拟误差总体减小, 模拟的茎蘖增长速率和最大茎蘖密度与实际吻合较好, 但在部分验证数据上仍存在较大误差。然而, 本文提出的验证和改进光温影响方程的方法, 对了解光温影响机制和完善群体茎蘖动态模拟模型具有一定的参考价值。
, JIANG Xiao-DongIn order to investigate the effects of light and temperature on rice tillering dynamics in tillering stage and validate the light and temperature effect equation in current dynamic tillering models. We performed two-year field-seeding experiments (2012-2013) using two indica two-line hybrid rice cultivars, Lingliangyou 268 and Liangyoupeijiu. And we collected the observation data in tillering. To fit Richards equation for retrieving characteristic parameters related to the maximum tiller density, average growth rate of tillers, and duration of the growing period. Moreover, we analyzed the relationships between all these characteristic parameters and average data of light, temperature and climatic variables during the growing period for each rice cultivar. Based on the relationships we established co-effect equations for the growth rate of tillers and maximum tiller density as a function of co-effect of light and temperature. The new co-effect functions for each cultivar were substituted for the effect functions of light and temperature in a widely used dynamic tillering model to simulate the dynamic tillering in tillering stage. Finally, validation and comparison were carried out for the models applied observation data in tillers period. The result showed that the light and temperature affected crop growth in tillering period. The growth rate of tillers and the actual maximum tiller density positively correlated with the two meteorological factors significantly. Compared with current dynamic tillering model, the model with the established co-effects of light and temperature functions decreased the error significantly in simulating the growth dynamics of tillers. The simulated rice tillering was well consistent with the observed in both growth rate and maximum tiller density for both rice cultivars. However, the discrepancy could also be found in some seeding periods and validation samples, which may be caused by rice adaptability to different light and temperature environments. In conclusion, the validation and improvement of the light and temperature co-effect functions put forward in this study can be used further for understanding the effects of light and temperature factors on rice tillering and improving dynamic tillering models in the future.
水稻产量由单位面积穗数、每穗粒数、结实率和千粒重组成[1, 2, 3, 4]。其中, 单位面积穗数是由移栽密度、单株分蘖数和分蘖成穗率三者构成。为提高水稻产量, 增加水稻穗数成为重要的途径之一[5, 6, 7, 8, 9]。决定穗数的关键时期在水稻分蘖期[10], 在该时期, 合理的栽培措施是提高水稻分蘖能力和单位面积穗数的重要保障。除品种因素外, 水稻分蘖能力还受到同期光温资源配比的影响[11, 12, 13]。因此, 有必要进一步研究水稻茎蘖动态与光温要素的关系, 为在气候增暖趋势下稳定和提高水稻群体分蘖能力提供农业气象服务依据。
在大田水肥管理一致的情形下, 水稻群体茎蘖动态能够反映出群体分蘖能力随光温环境的变化。因此, 大量研究通过分析和模拟水稻群体茎蘖动态的变化, 研究水稻分蘖能力随环境变化的响应机制。水稻群体茎蘖动态通常被划分为增长和消亡两个阶段。对于移栽稻, 增长阶段通常默认为从移栽开始直至最高分蘖期。该阶段被认为受光温条件的影响最大。紧接着水稻群体茎蘖密度减少直至有效分蘖终止期, 该阶段水稻群体茎蘖的变化主要受群体光合产物及干物质分配的影响[11, 12, 13, 14]。
已有研究显示, 在水稻茎蘖增长阶段, 水稻分蘖能力与光温条件的组合有密切联系[11]。例如, Kumagai等[14]在紫外辐射对水稻生产及产量影响的研究中发现, 温度的降低和光照的减少会导致水稻分蘖数的减少。而进一步的研究表明不同的光温组合条件对水稻分蘖能力的影响不同, 在较低的光照条件下, 光合产物较少, 导致分蘖芽无法正常生长, 因此在这种低光照条件下较低的温度反而能产生较多的分蘖; 但在光照充足的条件下, 温度越高, 能产生的分蘖越多。另外, 不同品种水稻的分蘖能力对光照和温度变化的响应特性不同[15]。对水稻茎蘖生长光温影响指标的研究表明, 分蘖发生的临界日辐射量为8.4 MJ m-2 d-1, 日辐射量低于该光强, 分蘖停止发生。分蘖生长最适温度为30~32℃, 日平均气温低于15℃或高于38℃时分蘖停止生长[16, 17]。
现有的水稻茎蘖动态模拟模型形式多样, 有以时间t为自变量的单因素统计模型, 也有综合气候及栽培因子等的多因素统计模型, 也有机制性较强的过程模型[11, 18, 19, 20, 21, 22, 23, 24, 25]。例如, FACE水稻茎蘖动态经验模型[18, 19]能够模拟CO2浓度倍增情形下的茎蘖动态。TIL水稻茎蘖动态模型[24, 25]以分蘖发生和死亡速率及干物重等为自变量建立了分蘖速率方程, 能够有效地模拟水稻茎蘖动态。黄耀等[11]则建立了机理性较强的水稻群体茎蘖动态模拟模型, 该模型综合考虑了作物品种特性、栽培技术以及环境因素对茎蘖生长的影响, 具有较高的模拟精度。上述模型中, FACE和TIL模型没有考虑光温要素对茎蘖生长的影响, 而黄耀等[11]建立的模型则考虑了光温要素对最高苗、茎蘖增长速率的作用, 但所用的影响方程未见确立的依据。因此, 本文拟结合2012年和2013年大田分期播种试验中获取的两种水稻品种的茎蘖数据, 分析茎蘖增长阶段光温要素与茎蘖动态的关系, 确立光温影响方程, 并替换黄耀等[11]建立的模型中的有关方程, 验证和比较替换前后模型在模拟不同光温条件下水稻群体茎蘖动态的效用, 为评估气候变化对水稻茎蘖动态的影响提供方法依据。
在南京信息工程大学农业气象试验站(32° 12′ 24″N, 118° 42′ 17″E)开展了为期2年的水稻大田试验(2012年和2013年)。供试品种为籼型两系杂交稻陵两优268和两优培九。在长江中下游单双季稻区, 陵两优268作为双季早稻种植, 而两优培九主要作为一季晚稻种植。
2012年和2013年均设置7个播期处理。2012年的播种日期分别为4月15日、4月30日、5月10日、5月20日、5月31日、6月10日和6月25日。由于2013年4月中旬气温偏低, 故将陵两优268的第1播期调整到4月23日, 剩余播期同2012年对应播期。同年, 两优培九则从4月30日开始第1播期, 剩余播期与2012年对应播期一致, 因此, 该年两优培九分6个播期。7个播期处理中, 第1、第2播期的秧龄为30 d, 第3~5播期的秧龄为25 d, 第6、第7播期的秧龄为20 d。移栽时叶龄平均4.5叶, 行距26 cm, 株距17 cm, 单株栽插。试验小区面积为4 m× 4 m, 区组设计完全随机, 对每个播期每个品种设置3个重复。水肥等田间管理按照常规高产田的要求进行。
在移栽返青后, 选择各小区内长势均匀的连续10穴水稻进行标记。随后, 每隔7 d对标记水稻进行生育期和茎蘖密度的观测。其中, 对水稻茎蘖密度的观测持续至抽穗期, 此时, 记录的水稻茎蘖密度为有效茎蘖密度。取每次每个小区观测的10穴茎蘖数据的平均值, 乘以移栽密度得到群体茎蘖数据。
在观测同期, 还获取了来自试验站内WatchDog小型自动气象站记录的10 min一次太阳辐射、气温和降水量等气象要素。对这些要素进行时间尺度上的整合和平均, 输出为逐日太阳总辐射、日最高温度、日最低温度和日总降水量。
以茎蘖增长阶段的群体茎蘖为研究对象, 并考虑到记录的茎蘖数据难免存在一定的误差, 因此选用Richards方程对群体茎蘖增长阶段数据的时间序列进行拟合, 得到平滑的群体茎蘖增长曲线。根据获得的曲线提取表征水稻群体茎蘖动态的特征参数, 即最大群体茎蘖数(A, 条 m-2)、平均群体茎蘖增长速率(Gm, 条 m-2 d-1)和大田茎蘖增长期(D, d)。以移栽后天数为自变量t(d), 以茎蘖数W (条 m-2)为因变量。
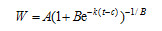 |
式中, A为最大群体茎蘖密度(条 m-2), k为生长速率参数, B和c为方程曲线的定型参数。
大田茎蘖增长期定义为移栽当日t1至到达最大群体茎蘖数t2之间的日数, 即D=t2-t1。在此期间群体茎蘖平均增长速率Gm(条 m-2 d-1)表达如下;
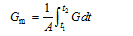 |
式中, G为逐日群体茎蘖增长率(条m-2 d-1), 即式(1)的一阶导数。
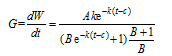 |
以黄耀等[11]建立的水稻群体茎蘖动态模型为基础, 模拟水稻茎蘖增长阶段的群体茎蘖动态。该模型主要采用乘性模型将光、温、水、肥以及自身的竞争作用影响相结合, 模拟内外部环境对水稻茎蘖增长的影响。其中, 光温的影响均采用非线性的分段函数来表示, 函数中定义了临界光强(20 MJ m-2 d-1)、下限温度(15℃)、最适温度区间(30~33℃)和上限温度(40℃)。当光强低于临界光强或温度处在非最适温度区间时, 光温组合将对群体茎蘖生长产生影响; 水的限制表达为烤田天数的函数, 而氮肥的影响则依据土壤肥力和施氮量; 自身的竞争作用由品种特性(分蘖率)、氮肥水平和移栽基本苗共同确定。上述因子直接作用于模型中的茎蘖增长系数, 计算每天实际的茎蘖增长率, 再累计求和模拟茎蘖增长。
总体上看, 该模型在模拟茎蘖增长阶段时表现出较强的机理性, 参数和变量的物理意义明确, 也易于编程实现, 但模型在实际应用时可能低估了光温要素的作用。例如, 在模拟最高苗时, 除品种特性外, 仅考虑了氮素的影响。根据2012年和2013年观测的各播期最大群体茎蘖密度发现, 实际的最高苗与光温组合条件关系密切。每日的光温条件不但影响茎蘖的增长速率, 还影响到潜在的最大茎蘖密度。为此, 根据试验数据分别建立茎蘖增长速率和分蘖率的光温影响方程, 将方程替换模型中原有的方程和相关变量。
本文采用统计回归方法建立上述光温影响方程。但由于光温要素之间存在较高的相关性, 从大田资料中较难建立各自独立的影响方程, 因此, 选择光温组合因子K=RT/(T+R)为自变量, 分别以平均茎蘖增长速率Gm和分蘖率Le为因变量, 建立光温组合影响方程。其中, Le是A、单株理论分蘖数S0和移栽基本苗P0的函数[5], R为光照强度(MJ m-2 d-1), T为日平均气温(℃)。对获取的方程进行归一化处理, 使得光温影响系数在0~1之间。以F(K)代表光温组合影响方程通式, 则归一化影响系数C的计算公式如下:
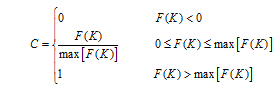 |
式中, max[F(K)]为上下临界温度和光照范围内的最大F(K), 如T在15~40℃和R在0~20 MJ m-2 d-1范围内的最大F(K)。但由于缺乏两品种分蘖的适宜光温指标, 针对Gm, max[F(K)]设置为P0与肥水适宜条件下的最大分蘖率Le0的乘积。就此, 分别形成Gm和Le的归一化光温组合影响方程FG(K)和FL(K)。这里FG(K)代表茎蘖增长速率的光温影响方程, FL(K)代表分蘖率的光温影响方程。将FG(K)计算的影响系数替换模型中茎蘖增长速率的光温影响系数PRF× PTF。类似地, 将模型中计算肥水适宜条件下实际分蘖率Le的方法修改如下;
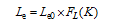 |
式中, Le0与品种有关。在实际应用中, 需要从试验资料获取。本文通过调试后, 对陵两优268和两优培九分别取0.72和0.80。
计算两个品种群体茎蘖大田增长期内的光照和温度累积量与平均值, 以及同期内的高温日数(T≥ 35℃且T≥ 30℃)、热积温、低温日数(T≤ 24℃)和冷积温(T≤ 24℃)。分析这些要素与水稻群体茎蘖增长参数的相关关系, 同时采用单因素方差分析和最小显著性差异(LSD)方法多重比较检验各播期观测的水稻群体茎蘖数据和群体茎蘖生长参数。统计分析均利用SPSS19.0软件完成。
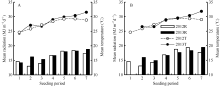
在两年试验中, 各播期水稻群体茎蘖增长经历了不同的光温环境。茎蘖增长期内光温水平随播期推后呈升高趋势(图1)。由图1-A可见, 2012年陵两优268群体茎蘖增长期内平均光照随播期呈先减小后增大的趋势, 平均气温呈先增大后减小的趋势, 其中平均最大光照出现在第6期, 约18.36 MJ m-2 d-1, 平均最大气温出现在第5期, 约29.29℃。2013年陵两优268群体茎蘖增长期内平均气温和平均光照随播期整体呈现逐渐增大的趋势, 平均光照和平均气温最大值均出现在第7期, 最大光照约为18.8 MJ m-2 d-1, 最高气温约为31.55℃。两优培九两年群体茎蘖增长期内平均光温随播期的变化趋势与陵两优268相似, 2012年平均光照最大值出现在第6期, 约19.36 MJ m-2d-1, 2013年出现在第7期, 约19.43 MJ m-2d-1; 平均气温最大值2012年出现在第5期, 约29.4℃, 2013年出现在第7期, 约31.87℃。
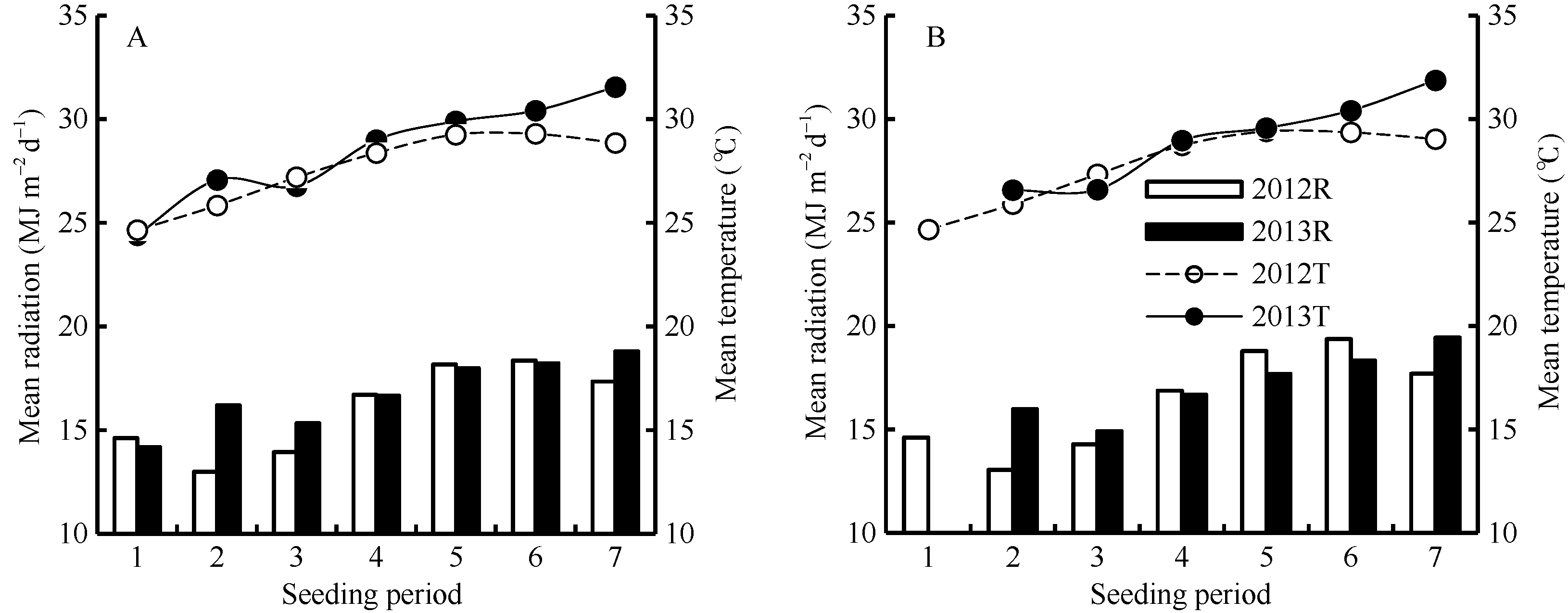 | 图1 2012-2013年陵两优268(A)和两优培九(B)各播期群体茎蘖增长期内平均光照与气温Fig. 1 Average daily total radiation and mean temperature during increasing period of tillering for Lingliangyou 268 (A) and Liangyoupeijiu (B) with different seeding periods in 2012 and 2013, respectively |
两年不同茎蘖增长期的光温条件差异是各播期水稻群体茎蘖动态特征差异的主要原因。从两品种两年不同播期群体茎蘖动态特征参数和茎蘖增长期内平均光温的方差分析看(表1和表2), 2012年陵两优268各茎蘖特征参数和增长期内平均光温均表现出显著的播期差异, 但2013年上述变量的播期差异不显著; 与此类似, 两优培九2012年各变量播期差异显著, 但2013年最大茎蘖数和茎蘖增长期的播期差异不显著。
两品种的群体茎蘖动态特征参数与光温要素之间的相关性分别如表3和表4所示。两品种茎蘖增长速率以及最大群体茎蘖数与茎蘖增长期内的平均光温条件具有显著的正相关性, 表明光照增强和温度升高有利于促进分蘖, 使茎蘖生长速率以及最大茎蘖数增大。茎蘖增长速率、最大茎蘖数与积温、累积光照的负相关性不显著, 但茎蘖增长期与积温、累积光照表现出极显著的正相关性, 这主要与平均茎蘖增长速率的增大导致茎蘖增长期缩短有关。
此外, 表3和表4还显示了茎蘖动态特征参数与气候要素的关系。陵两优268的群体平均茎蘖生长速率、最大茎蘖数与低温天数、冷积温表现为显著的负相关, 表明在低温胁迫下茎蘖生长受到抑制。与此相反, 上述特征参数与高温天数、热积温均表现为极显著的正相关, 暗示高温环境可能对分蘖生长有促进作用。然而, 这样的作用离不开高温期间的稻田灌溉。通过灌溉降低了稻田土壤的温度, 不但对根系和茎蘖起到了保护作用, 还促进了分蘖, 使得最大茎蘖数有显著的增加。另外, 高温显著提高了茎蘖增长速率, 使得茎蘖增长期明显缩短。同样, 两优培九各参数与极端温度条件也表现出与陵两优268类似的关系。
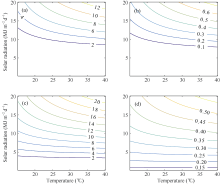
从两年所有播期茎蘖动态观测数据中随机选取一半用于光温组合影响方程的构建和模型参数标定, 另一半则用于验证。由统计回归方法获取的光温组合影响方程如表5所示。表中, Gm和Le与K建立的回归方程都通过了0.01水平的显著性检验, 两者与K的回归关系均为一般线性方程。图2显示了两品种FG和FL的光温变化曲线。两者对光照和温度的变化均呈现为非线性关系。光照一定时, FG和FL随温度的降低而减小。温度一定时, FG和FL随光照的减少而减小。在低光照条件下(< 6 MJ m-2 d-1), 两优培九的FG和FL对温度变化不敏感, 而陵两优268的分蘖受到明显抑制, 表明两品种群体茎蘖动态对于光温的响应存在差异。
| 表1 陵两优268各播期群体茎蘖生长参数和增长期内光温要素的方差分析 Table 1 Variance analysis of tillering characteristic parameters and radiation and temperature variables between seeding periods for Lingliangyou 268 |
| 表2 两优培九各播期群体茎蘖生长系数和茎蘖增长期光温要素的方差分析 Table 2 Variance analysis of tillering characteristic parameters and radiation and temperature variables between seeding periods for Liangyoupeijiu |
| 表3 陵两优268群体茎蘖生长参数与光温要素的相关系数 Table 3 Correlation coefficients between tillering characteristic parameters and radiation and temperature factors for Lingliangyou 268 |
| 表4 两优培九群体茎蘖生长参数与光温要素的相关系数 Table 4 Correlation coefficients between tillering characteristic parameters and radiation and temperature factors for Liangyoupeijiu |
| 表5 光温组合影响方程 Table 5 Equations of combined effect from radiation and temperature for average growth rate of tillers Gm and final tillering rateLe |
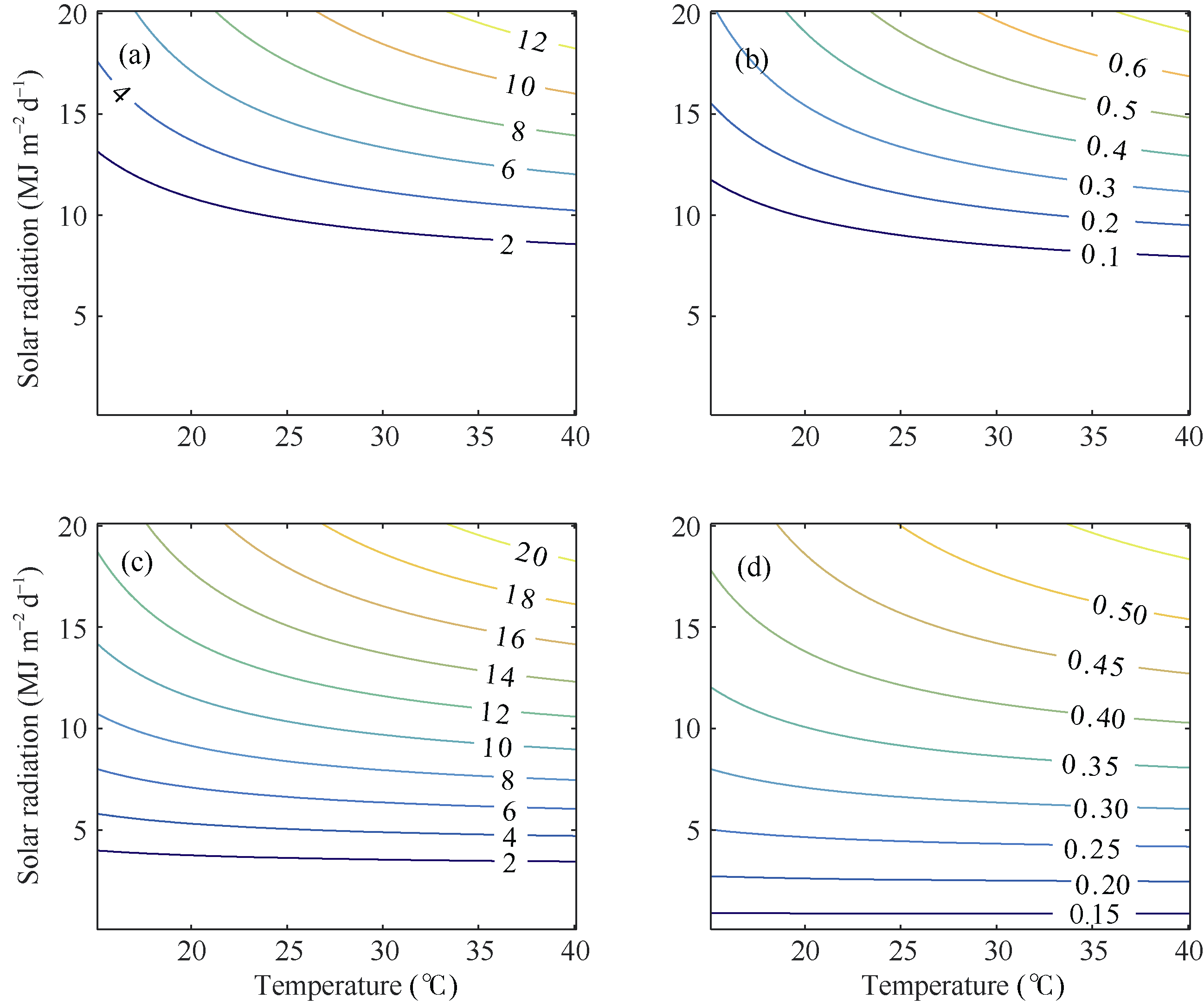 | 图2 两品种FG和FL的光温响应关系 (a)和(b)分别对应陵两优268的FG和FL; (c)和(d)分别对应两优培九的FG和FL。Fig. 2 Changes of FG and FL in response to radiation and temperature for both rice varieties (a) and (b) for FG and FL of Lingliangyou 268, respectively; (c) and (d) for FG and FL of Liangyoupeijiu, respectively. |
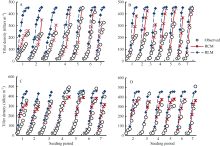
结合验证数据, 对替换前后模型的模拟结果与实测数据进行了比较。为了便于分析和阐述, 用RLM表示原模型, 用RCM表示替换了光温组合影响方程后的模型。图3显示了两模型的模拟结果。相比RLM, RCM模拟的两品种茎蘖增长趋势与实际情况更接近, 模拟的最大茎蘖密度与实测值误差较小, 且年份和播期之间的变化得到较好的模拟。值得注意的是, 茎蘖增长期经历7、8月份的播期, 其茎蘖增长趋势和最大茎蘖密度模拟结果普遍较好。如2012年陵两优268的第5和第6期及2013年的第6和第7期, 两优培九2012年的第5和第6期及2013年的第6期。对应茎蘖增长期经历5、6月份的播期, 模拟结果较好的主要集中在两优培九的第1~3期。这进一步说明了低温环境对陵两优268茎蘖增长的影响要大于两优培九。
虽然RCM的模拟误差相对于RLM有所减小, 但模拟的两品种茎蘖增长速率和最大茎蘖密度仍存在一定的误差。例如, 两优培九2012年最大茎蘖密度模拟值整体偏小, 且2013年该品种所有播期最大密度茎蘖模拟值均接近于370条 m-2, 未反映出实际播期间的差异。与两优培九相反, 陵两优268的最大茎蘖密度模拟值整体偏高, 2012年的前4个播期模拟值大于实测值, 而2013年的模拟值普遍大于实测值。此外, 模拟的两品种2012年第1期茎蘖增长趋势明显偏大, 表明光温组合影响方程低估了前期低温作用。
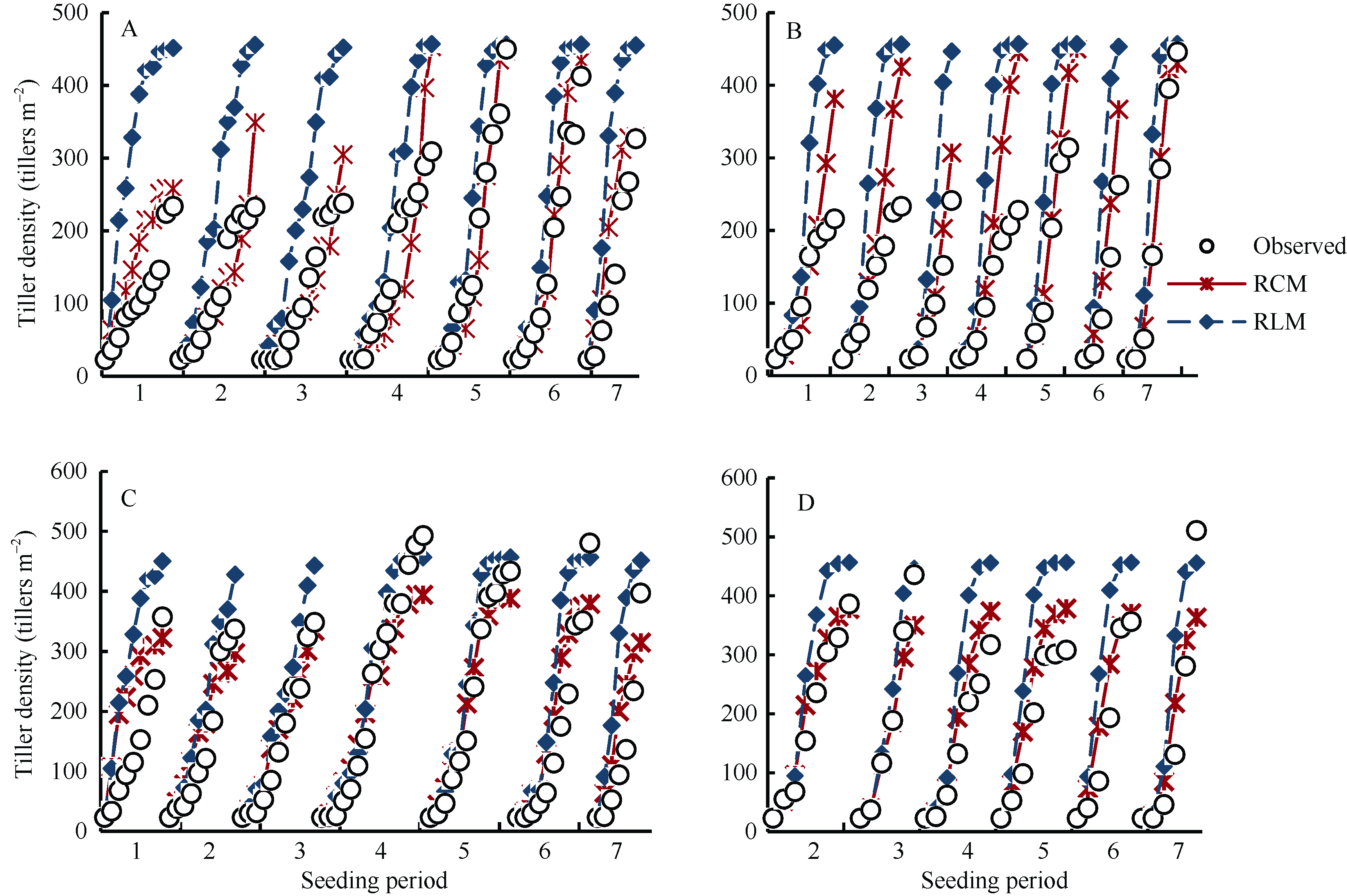 | 图3 陵两优268 2012年(A)和2013年(B)及两优培九2012年(C)和2013年(D)群体茎蘖动态模拟值与实测值的比较Fig. 3 Comparison between simulated tiller density and the observed for Lingliangyou 268 in 2012 (A) and 2013 (B), and for Liangyoupeijiu in 2012 (C) and 2013 (D), respectively |
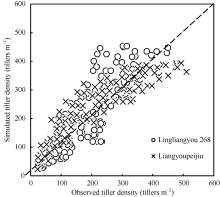
图4显示了两品种验证数据与模拟结果的1∶ 1关系。图中显示, 两优培九的模拟误差主要出现在茎蘖增长阶段的中后期, 随着茎蘖不断增长, 模型模拟的误差也在逐渐增大。对于陵两优268, 模拟误差主要集中在茎蘖增长的后期, 即高估了茎蘖增长趋势。总体上看, 两优培九茎蘖增长动态的模拟结果优于陵两优268。
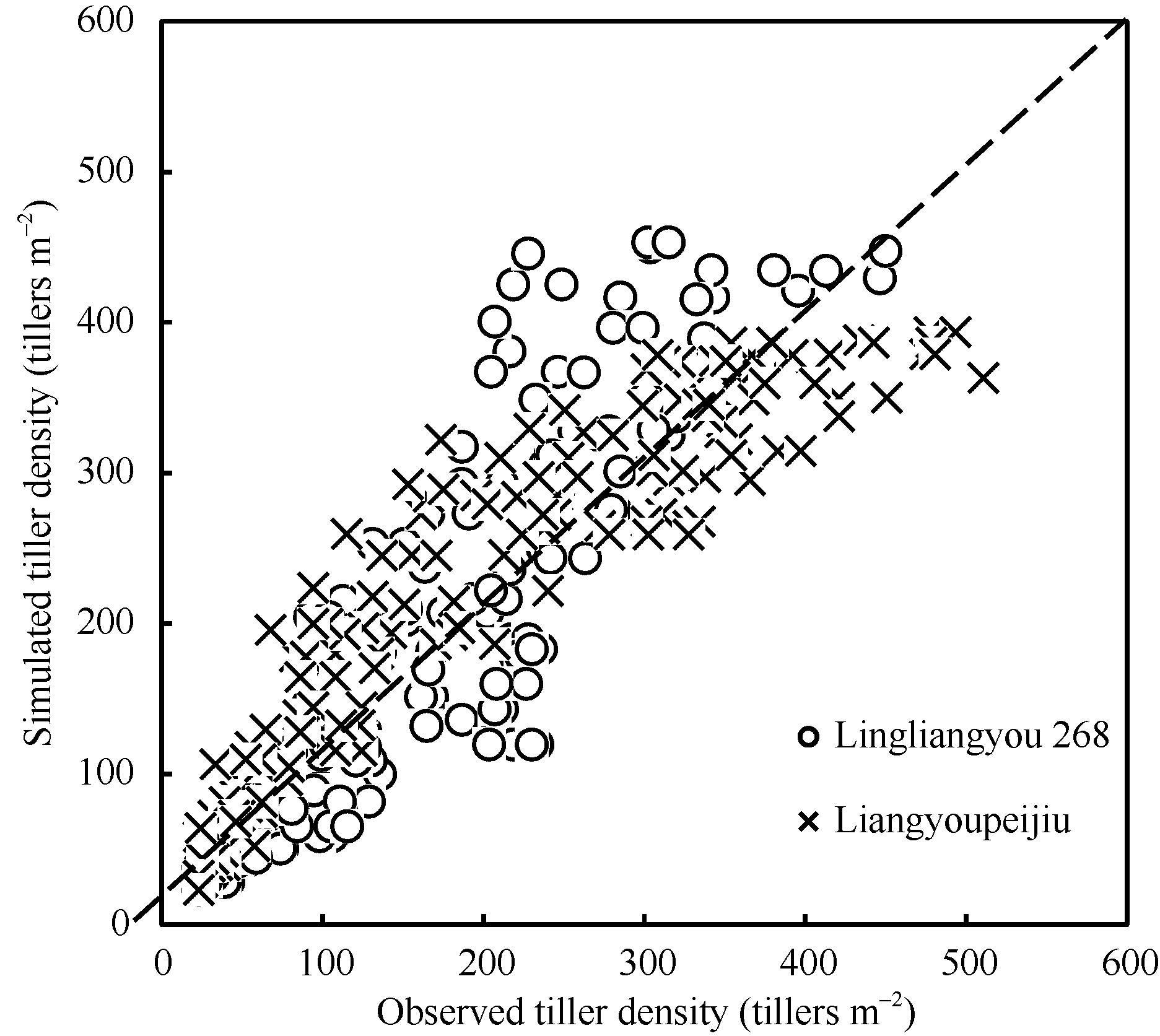 | 图4 陵两优268和两优培九水稻群体茎蘖动态实测值与模拟值比较Fig. 4 Comparison of simulated and observed tiller density for both Liangyoupeijiu and Lingliangyou 268, respectively |
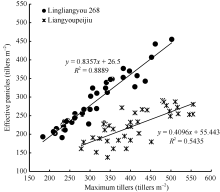
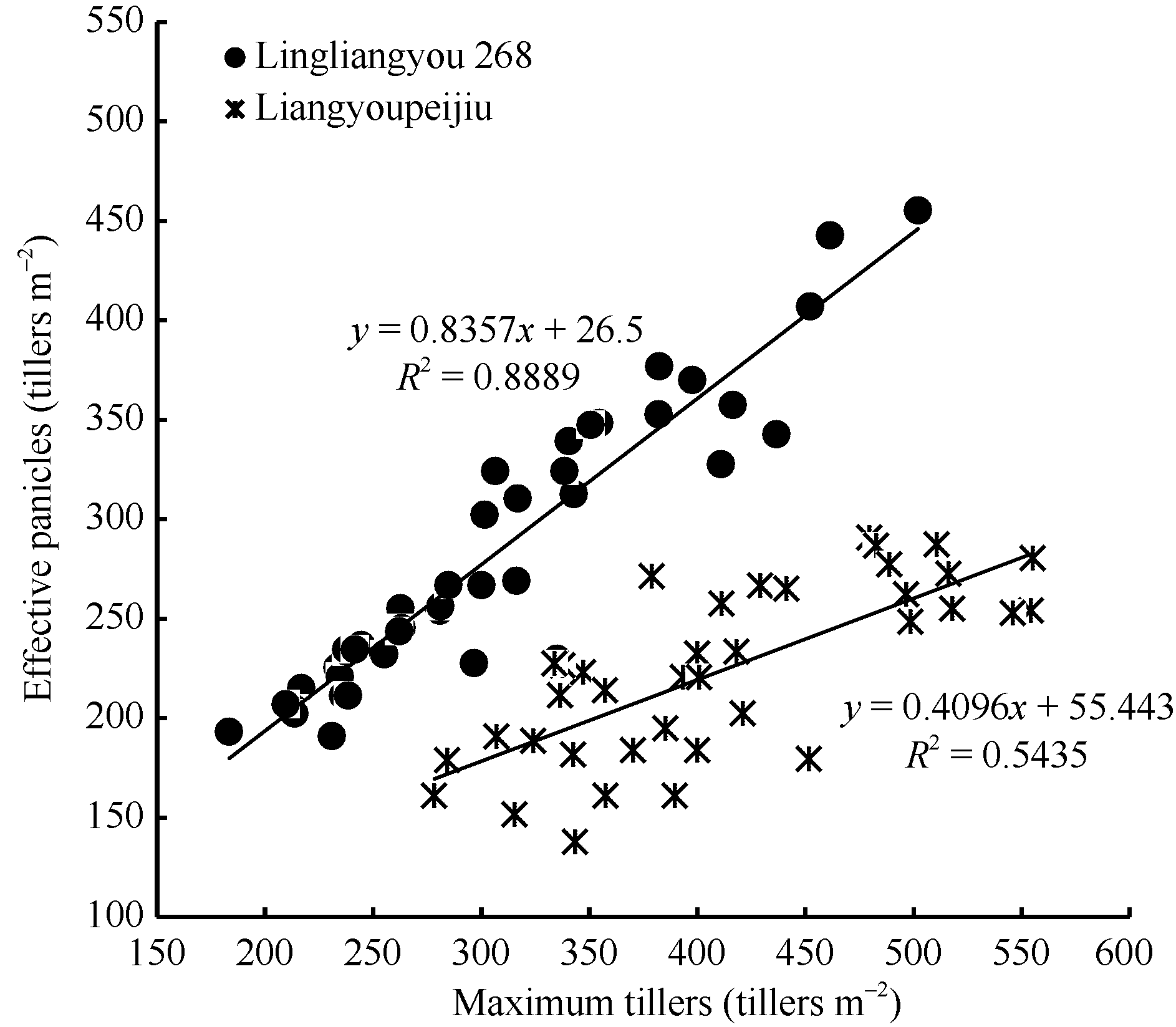 | 图5 陵两优268和两优培九群体最大茎蘖数与群体最大有效茎蘖数的关系Fig. 5 Comparison between observed number of effective panicles and maximum tiller density for Lingliangyou 268 and Liangyoupeijiu, respectively |
水稻产量要素中, 有效穗是重要的一个。有效穗的多少不但取决于品种分蘖能力、抽穗前生长量、移栽密度和水肥管理等, 还与整个阶段气象条件有关[26]。本文选择水稻移栽至最高分蘖期的群体茎蘖动态作为研究对象, 一方面是该时期内光温要素影响显著, 另一方面是达到的最大茎蘖密度与有效穗数有着密切的联系[12, 27, 28, 29](图5)。因此, 通过研究增长阶段的茎蘖动态可以为有效穗的估算提供重要依据。
结合2012年和2013年水稻分期播种试验, 分析了水稻茎蘖增长阶段光温要素对平均增长速率、最大茎蘖密度和增长期的影响。从结果看, 群体茎蘖增长受增长期光温要素的共同作用, 平均茎蘖增长速率以及最大茎蘖密度与光温要素均显著正相关, 但不同品种之间存在一定的差异, 与已有研究结果吻合[30, 31, 32, 33, 34]。通过分析增长期内极端温度对茎蘖增长的影响, 高温热害天数与茎蘖特征参数之间具有较高的正相关, 似乎表明高温有利于分蘖的发生, 与已有的研究结论[11, 16]相悖。这可能与不同研究采用的品种存在遗传特性上的差异有关。但2013年夏季出现了较长时间的高温热害, 从试验资料看, 高温热害并未明显抑制分蘖, 反而表现出促进作用, 这与试验期间有效的灌溉降温措施有关。相比高温, 低温环境则表现出对茎蘖增长明显的抑制作用。
受到试验的限制, 从大田观测中无法分离光温对茎蘖增长的影响。在增长期内, 光温要素表现出较强的相关性, 因此, 选择光温组合因子K用于分析和构建针对水稻茎蘖增长的光温组合影响方程。从模拟结果看, 该因子较好地表达了光温要素对茎蘖增长的共同作用。构建的光温组合影响方程, 不但确定了逐日光温对茎蘖日增量的贡献, 还决定了光温条件对最大茎蘖密度的影响。总体上看, 光温组合影响系数随K值的增大而增大, 即随着K的增大, 茎蘖增长速率与分蘖率都相应增大, 该趋势与实际观测相符。
相比RLM, RCM在模拟精度上有一定的提高, 但仍无法避免出现一些较大的误差。这与试验、建模过程和其他未知因素有关。就试验而言, 试验环境和田间管理无法达到绝对的一致, 也无法避免人为观测误差。在建模过程中, Richards方程在拟合观测数据并提取特征参数时会引入新的误差, 这个误差与观测误差一起影响光温组合影响方程的精度。另外, 选取光温要素和特征参数的平均量参与建模, 难免会出现高估或低估不利温度环境的影响, 从而增加了模拟误差。此外, 水稻都具有一定的气候适应能力, 在两年气候条件存在明显差异的情况下, 使用相同的潜在最大分蘖率(Le0)等参数可能与实际情况不符。最后, RCM中的光温影响函数具有一定的普适性, 但在实际应用中, 函数中的参数需要针对研究品种的生理特性进行标定。
光温要素不仅影响群体茎蘖增长速率, 也会影响群体最大分蘖能力。建立的光温组合影响方程具有较强的可靠性。在替换原模型中的有关方程后, 模拟的茎蘖增长速率和最大茎蘖密度均具有较高的精度, 但仍需要结合更多的观测数据研究光温影响机制并完善光温影响方程。获取不同品种水稻群体茎蘖生长的光温特性, 为应对气候变化制定高产栽培计划、优化品种种植布局具有重要的指导作用。光温影响方程的建立对进一步量化光温对群体茎蘖增长的影响具有重要的意义, 也为今后茎蘖消亡阶段的光温影响研究奠定基础。
致谢: 感谢江苏省农业科学院金之庆研究员对论文撰写提出的宝贵意见。感谢南京信息工程大学张弥博士对论文撰写的指导和帮助。感谢湖北省气象局农业气象室冯明研究员对论文研究给予的指导和帮助。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|