




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
大豆14-3-3蛋白与转录因子蛋白GmMYB173的互作
[董萌1, 2 , 高友菲2 , 韩天富2 , 东方阳1, *  , 蒋炳军
, 蒋炳军2, * ]
, 蒋炳军]
|
|


第一作者联系方式: E-mail: dongmeng902@163.com, Tel: 13164257831
14-3-3蛋白家族在真核生物中普遍存在, 可与其他蛋白相互作用, 调控多种生理生化过程。MYB基因家族作为植物中最大的一类转录因子, 广泛参与了植物的生长发育和代谢调控。本研究通过分析1个从自贡冬豆中克隆的MYB转录因子GmMYB173的亚细胞定位情况, 发现GmMYB173在细胞核中特异表达; 序列分析发现GmMYB173与GmMYB176相似, 具有1个14-3-3蛋白的潜在结合结构域, 即pST结合结构域。通过重叠延伸PCR (SOE-PCR)删除了GmMYB173序列中pST结合结构域编码序列, 发现GmMYB173细胞核表达特异性消失。酵母双杂交互作分析表明, 大豆基因组中所有具有表达的16个14-3-3蛋白GmSGF14a~GmSGF14p均能与GmMYB173互作。β-半乳糖苷酶活性分析发现, 与GmMYB173互作最强的是GmSGF14n, 其次是GmSGF14k、GmSGF14e和GmSGF14o。这些结果说明14-3-3蛋白不仅与GmMYB173互作, 且可能调控其在细胞内的定位, 有助于研究14-3-3蛋白与GmMYB173的互作关系及其在大豆生长发育中的作用。
14-3-3 proteins, nearly existing in all eukaryotic cells, may regulate many physiological and biochemical processes through interacting with other proteins. As the largest class of transcription factors in plants, MYB gene family is widely involved in plant growth and metabolism regulation. A gene cloned from soybean cultivar Zigongdongdou, and specifically expressed in the nuclear in the subcellular location assay. Sequence analysis showed that there was a binding site of 14-4-4 proteins, namely the pST binding site, in GmMYB173 similar to that in GmMYB176. The nuclear-specific expression of GmMYB173 disappeared when the sequence of the pST binding site was deleted through the splicing by overlap extension PCR. All 14-3-3 proteins from GmSGF14a to GmSGF14p could interact with GmMYB173. Among them, GmSGF14n interacted with GmMYB173 strongest, GmSGF14e and GmSGF14k took second place, which was proved by the β-galactosidase activity analysis. These results suggest that 14-3-3 proteins not only interact with GmMYB173, but also probably regulate its subcellular location. The information provided by this study will facilitate the study of interaction relationship between 14-3-3 proteins and GmMYB173 and its function on the soybean development.
14-3-3蛋白是一类在真核生物中广泛存在的蛋白。关于植物中14-3-3蛋白的报道最先见于大麦[1]、拟南芥[2]和菠菜[3]。目前组学研究发现, 14-3-3蛋白在植物基因组中存在多个编码基因, 呈现差异表达。在拟南芥基因组中已经鉴定出15个14-3-3蛋白基因, 其中13个有转录表达[4, 5]。在水稻中鉴定出8个, 其中6个有转录表达[6]。Li等[7]在大豆基因组中发现了18个14-3-3蛋白基因, 其中16个即GmSGF14a~ GmSGF14p可通过RT-PCR检测到其表达。14-3-3蛋白在植物体内主要以同源或异源二聚体的形式存在。14-3-3蛋白与其他蛋白的结合主要通过磷酸化的多肽域来实现, 包括2种序列模式, 一种是RSXpSXP, 另一种是RXY/FXpSXP, 其中X为任意氨基酸, pS是磷酸化的丝氨酸[8]。但是, 一些14-3-3蛋白也能通过识别非磷酸化的序列来实现与互作蛋白的结合[9, 10]。由此可见, 不同14-3-3蛋白可能由于表达差异化、结合蛋白多样化, 而呈现丰富的生理生化功能。
植物14-3-3蛋白家族基因调控着植物中多种生理生化过程。14-3-3蛋白在细胞核中能够调节转录激活子的亚细胞定位和基因的表达。Carrasco等[11]研究发现14-3-3蛋白能与DNA结合蛋白磷酸酶(DBP1)结合而调控转录激活因子的亚细胞定位, 并能将DBP1转运出细胞核, 且能与DBP1结合从而调控目的基因CEVI1的表达以应对病毒入侵带来的危害。14-3-3蛋白还参与了植物的光信号通路。Folta等[12]利用酵母双杂交和免疫共沉淀证明14-3-3蛋白能够与重要的光周期蛋白Constans相互作用, 暗示14-3-3蛋白参与了植物的光信号途径。此外, 14-3-3蛋白还参与了蛋白合成、蛋白折叠、质膜质子泵的初级代谢、激素代谢和染色质重组等生理生化过程[13, 14]。Hajduch等[15]对大豆种子的蛋白质组学分析发现14-3-3蛋白对于大豆种子发育有重要的作用。
MYB超级家族是序列特异性转录因子家族之一, 具有种类多和功能多样化的特点[16], 参与细胞分化、细胞周期的调节, 激素和环境因子应答, 并对植物的次生代谢以及叶片等器官形态的建成具有重要的调节作用[17]。Liao等[18]在大豆中获得了48个具有基因全长开放阅读框的MYB类似基因, 其中43个基因对盐、干旱、低温和ABA处理产生了相应的应答反应。李晓薇[19]从大豆中克隆了两个R2R3-MYB转录因子基因GmMYB12a和GmMYB12B2, 对转基因拟南芥的研究发现, 植物类黄酮合成途径中的多个关键酶基因均受这2个基因的正向调控。Liu等[20]研究发现, GmMYB73可能通过减少GL2, 进而释放GL-2Y抑制性的PLDa1表达来积累油脂, GmMYB73的调控潜在提高豆类作物的油产量。Chen等[21]发现MYB56为CULLIN3 (CUL3)-based E3连接酶的一个新的靶位点, 遗传学研究表明MYB56是开花的负调控因子。Liu等[22]研究发现韧皮部中MYB30表达水平的提高通过直接调控FT而促进拟南芥的开花。
14-3-3蛋白和MYB类转录因子分别在植物的生理生化代谢中发挥重要的作用, 但是对于二者相互作用共同发挥作用的研究较少。Yi等[23]和Li等[24]发现了1个单重复结构域的MYB转录因子基因GmMYB176。在大豆中, GmMYB176能够调节CHS8的表达, 但是在发根中过表达此基因不足以增加CHS8的表达量和异黄酮的含量, 而与其互作的14-3-3蛋白编码基因GmSGF14i的过表达引起了异黄酮表达模式的改变。高友菲等[25]利用酵母单杂交技术筛选到自贡冬豆中与T1元件结合的MYB转录因子GmMYB173, 对GmMYB173功能及其调节机制的研究将对后续研究具有重要意义。
本研究利用SOE-PCR及亚细胞定位间接证明了14-3-3蛋白对于GmMYB173亚细胞定位的影响, 利用酵母双杂交证明了大豆中已知的16种在转录水平上表达的14-3-3蛋白能够与GmMYB173互作, 其中GmSGF14n编码的14-3-3蛋白与GmMYB173互作的活性最强, 为后续深入研究14-3-3蛋白及GmMYB173的功能提供了一定的分子基础。
大豆品种自贡冬豆由中国农业科学院作物科学研究所农业部北京大豆生物学重点实验室保存。将大豆种子播于混有营养基质和蛭石(体积比1∶ 1)的花盆中, 置人工气候箱培养至第1片三出复叶完全展开, 取3株不同植株叶片于液氮中速冻后放入-80℃冰箱保存备用。洋葱购自超市。用于遗传转化的大肠杆菌(Eschrichia coli)菌株DH5α 购自北京天根生化科技有限公司。用于酵母双杂交的酵母菌(Saccharomyces cerevisiae)菌株AH109及表达载体p16318-GFP、pGADT7和pGBKT7由中国农业科学院作物科学研究所农业部北京大豆生物学重点实验室保存
根据GmMYB173的基因序列, 利用Primer Blast (http://www.ncbi.nlm.nih.gov/)设计克隆引物, 以实验室保存的pLB-GmMYB173为模板, F-1和R-1为引物(表1)进行PCR扩增, 条件为94℃ 2 min; 98℃ 10 s, 55℃ 30 s, 68℃ 25 s; 30个循环; 68℃ 10 min; Sal I和BamH I双酶切PCR产物和p16318-GFP连接构建表达载体p16318-GmMYB173。
| 表1 亚细胞定位载体基因克隆引物 Table 1 Primers of cloning gene in subcellular vector construction |
以p16318-GmMYB173为模板, 以F-2、R-2、F° 和R° 为引物, 利用SOE-PCR方法, 先后进行两轮PCR扩增, 获取缺失编码14-3-3蛋白结合多肽域的GmMYB173 deletion, 其中第1轮扩增条件为94℃ 2 min; 98℃ 10 s, 55℃ 30 s, 68℃ 15 s, 30个循环; 68℃ 10 min。第2轮扩增条件为94℃ 2 min; 98℃ 10 s, 52℃ 30 s, 68℃ 25 s, 30个循环; 68℃ 10 min。Hind III和BamH I双酶切PCR产物和p16318-GFP连接构建表达载体p16318-GmMYB173 deletion。
参照曲梦楠[26]《大豆GmFT2b基因的功能分析》中“ 洋葱表皮亚细胞定位观察方法” 。
根据NCBI数据库提供的基因信息设计克隆引物如表2所示, 并以自贡冬豆cDNA为模板, 对GmSGF14家族基因进行PCR克隆; 然后用相应限制性内切酶将GmSGF14家族基因连接到表达载体pGADT7。
参照Clontech的Yeast Protocols Handbook中提供的方法。挑取营养缺陷平板上单菌落到YPDA液体培养基中培养10~12 h, 提取质粒后进行菌落PCR验证。
| 表2 14-3-3蛋白编码基因GmSGF14家族基因信息 Table 2 Information of GmSGF14genes conforming 14-3-3 proteins |
目的基因GmMYB173的开放阅读框为852 bp, 编码284个氨基酸。利用BaCelLo (http://gpcr. biocomp.unibo.it/bacello/)对GmMYB173进行亚细胞预测, 结果显示其定位于细胞核中(图1)。其蛋白序列与NCBI中公布的部分MYB类蛋白序列比对发现, GmMYB173具有一个14-3-3蛋白的潜在结合位点(图2)。
 | 图1 GmMYB173的亚细胞定预测结果Fig. 1 Results of prediction of subcellular location of GmMYB173 |
 | 图2 GmMYB173与其他MYB类转录因子氨基酸序列比对图中黑色方框表示pST结合位点。Fig. 2 Sequence multi-alignment of GmMYB173 with other MYB transcription factorsThe pST binding sites were underlined by the black box. |
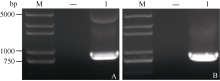
以保存的pLB-GmMYB173载体为模板, 加有相应酶切位点的克隆引物进行PCR扩增, 得到与目标大小相符的特异性条带(图3-A)。对测序正确的单克隆扩大培养后提取质粒进行双酶切验证, 酶切得到与目标基因大小相符的条带(图4), 证明GmMYB173已经正确连接入表达载体中。
在表达载体p16318-GmMYB173 deletion构建中, 亚克隆得到与GmMYB173 deletion大小相符的条带(图3-B)。测序后得到克隆无误的GmMYB173 deletion, 对提取的质粒进行了双酶切验证, 得到了大小略小于GmMYB173条带的840 bp左右的目标条带GmMYB173 deletion (图4), 表明GmMYB173 deletion已经正确连接入表达载体中。
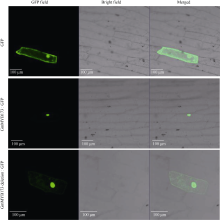
利用激光共聚焦显微镜对瞬时转化GFP、GmMYB173-GFP和GmMYB173 deletion-GFP载体的洋葱表皮细胞观察表明, 对照GFP可在洋葱表皮细胞中大量表达, 绿色荧光信号在细胞核, 细胞质均有分布; 与对照相比, GmMYB173-GFP绿色荧光信号主要分布在细胞核中, 在细胞质中未检测到明显的荧光信号; 而GmMYB173 deletion-GFP绿色荧光信号在细胞核和细胞质中均有分布, 失去了GmMYB173核表达的特异性(图5)。
 | 图3 GmMYB173 PCR和GmMYB173 deletion扩增产物A: M: DL2000 plus marker; 1: GmMYB173的PCR产物。B: M: DL2000 plus marker; 1: GmMYB173 deletion的PCR产物。Fig. 3 PCR products of GmMYB173 and GmMYB173 deletionA: M: DL2000 plus marker; 1: PCR products of GmMYB173; B: M: DL2000 plus marker; 1: PCR products of GmMYB173 deletion. |
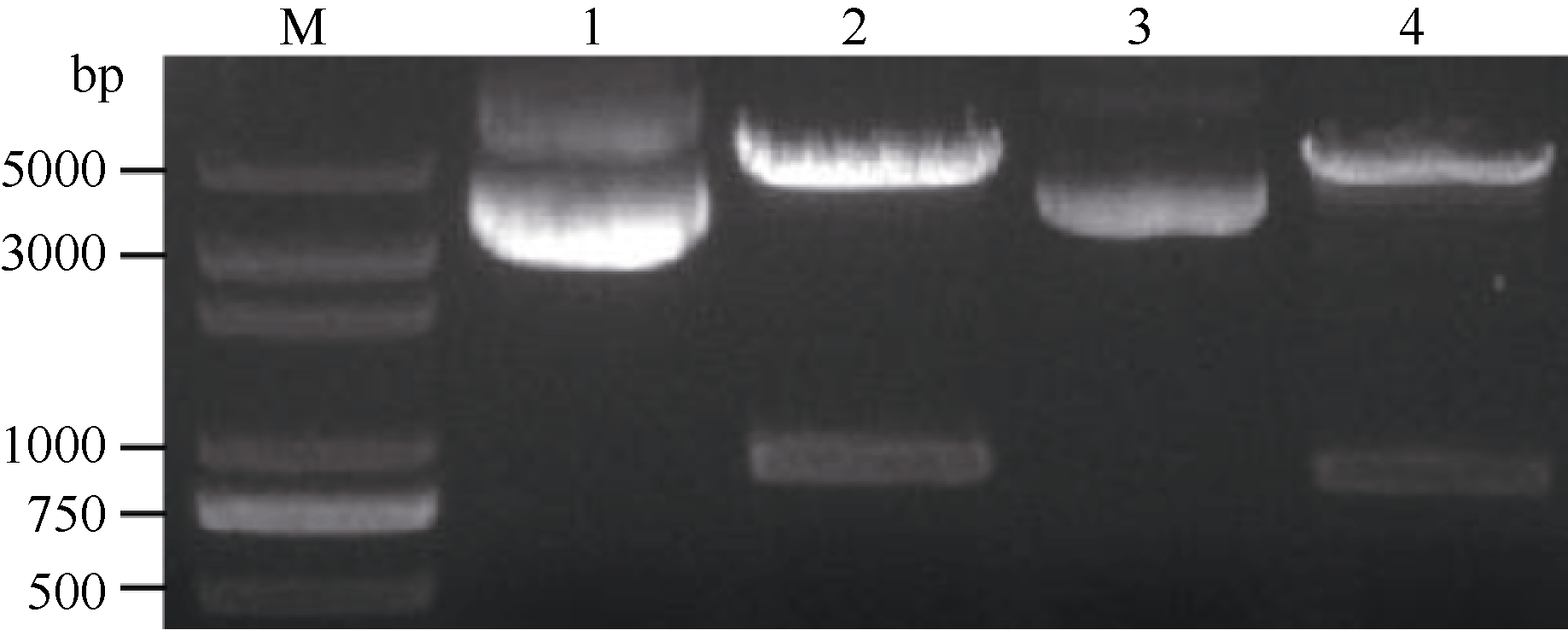 | 图4 p16318-GmMYB173和p16318-GmMYB173 deletion双酶切验证M: DL2000 plus marker; 1: p16318-GmMYB173质粒; 2: p16318-GmMYB173双酶切; 3: p16318-GmMYB173 deletion 质粒; 4: p16318-GmMYB173 deletion 双酶切。Fig. 4 Identification of p16318-GmMYB173 and p16318-GmMYB173 deletion double enzyme digestionM: DL2000 plus marker; 1: p16318-GmMYB173 vector; 2: p16318-GmMYB173 double enzyme digestion; 3: p16318-GmMYB173 deletion vector; p16318-GmMYB173 deletion double enzyme digestion. |
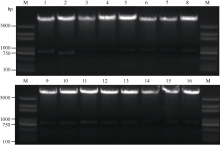
以自贡冬豆叶片cDNA为模板, 表2中克隆引物对GmSGF14家族基因进行克隆, 1%琼脂糖凝胶电泳分离, 得到与目标基因大小相符的特异性条带(图6), 胶经回收并连接pLB载体后测序。将序列与NCBI中的参考序列比对发现, GmSGF14a、GmSGF14b、GmSGF14c、GmSGF14d、GmSGF14e、GmSGF14g和GmSGF14m与参考序列之间存在多处的SNP, 其余GmSGF14家族基因与参考序列相同。将连接入表达载体的阳性克隆提取质粒后进行酶切验证, 所有载体均得到与目标基因大小相符的酶切片段, 表明目标基因已正确连入载体中(图7)。
 | 图5 GmMYB173-GFP和GmMYB173 deletion-GFP融合蛋白的亚细胞定位Fig. 5 Subcellular location of GmMYB173-GFP and GmMYB173 deletion-GFP fusion proteins |
 | 图6 14-3-3蛋白编码基因GmSGF14a-p PCR扩增产物M: DL2000 marker; 1~16:GmSGF14a-p克隆产物。Fig. 6 PCR products of 14-3-3 proteins coding genes GmSGF14a-pM: DL2000 marker; 1-16: products of genes GmSGF14a-p. |
 | 图7 pGADT7-GmSGF14a-p和pGBKT7-GmMYB173载体双酶切验证M: DL 2000 plus maker; 1~16: pGADT7-GmSGF14a-p载体双酶切; 17: pGBKT7-GmMYB173载体双酶切。Fig. 7 Identification of pGADT7-GmSGF14a-p and pGBKT7-GmMYB173 double enzyme digestionM: DL 2000 plus maker; 1-16: double enzyme digestion of pGADT7-GmSGF14a-p vectors; 17: double enzyme digestion of pGBKT7-GmMYB173 vector. |
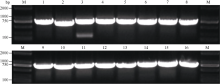
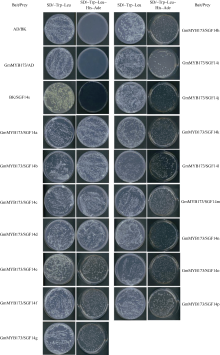
采用上述实验方法制备酵母感受态并转化, 转化后将感受态细胞分别涂布于营养缺陷平板SD/-Trp- Leu和SD/-Trp-Leu-Ade-His上。在SD/-Trp-Leu平板上均有酵母单菌落生长, 表明AD和BK载体已成功转入酵母中。在SD/-Trp-Leu-Ade-His平板上, 只转入AD和BK空载体的酵母不能在平板上生长。同时, 转入pGBKT7-GmMYB173和pGADT7载体的酵母也不能在平板上生长, 表明pGBKT7-GmMYB173不具有自激活活性。转入pGBKT7和pGADT7-GmSGF14 family载体的酵母也不能生长(在此以pGADT7- GmSGF14d的结果省略表示), 表明pGADT7- GmSGF14 family也不具有自激活活性。而同时转入pGBKT7-GmMYB173和pGADT7-GmSGF14 family的酵母菌能够生长出正常的菌斑(图8), 表明GmMYB173与GmSGF14 family存在互作。挑取SD/- Trp-Leu-Ade-His单菌落摇菌后提取质粒, 菌落PCR验证表明, pGBKT7-GmMYB173和pGADT7-GmSGF14 family均已转入酵母中(图9)。
 | 图8 GmMYB173与14-3-3蛋白的酵母双杂交结果Fig. 8 Results of yeast two-hybrid assay of GmMYB173 and 14-3-3 proteins |
 | 图9 酵母双杂实验中酵母单菌落的菌落PCR验证M: DL2000 marker; 1~16:GmMYB173基因的菌落PCR验证; a~p: 依次为GmSGF14a至GmSGF14p基因的菌落PCR验证。Fig. 9 PCR Identification of yeast individual colonyM: DL2000 marker; 1-16: individual colony-PCR ofGmMYB173; a-p: individual colony-PCR of GmSGF14a to GmSGF14p. |
参照上述实验方法测定β -半乳糖苷酶活性, 确定GmMYB173与14-3-3蛋白互作强度表明, 不同的14-3-3蛋白与GmMYB173互作之后激活的β -半乳糖苷酶活性存在一定的差异。其中, 对照AD/BK、GmMYB173/AD、SGF14d/BK在本底水平上存在一定的β -半乳糖苷酶活性, 与对照相比, GmSGF14n与GmMYB173互作后激活的β -半乳糖苷酶活性最强, GmSGF14k、GmSGF14e和GmSGF14o稍次之(图10), 暗示GmSGF14n与GmMYB173在植物体中相互作用来发挥一定的生物学功能。
蛋白质的亚细胞定位是功能基因组学研究的重要内容, 对其研究有助于更加系统全面地理解植物的生长发育, 形态建成等。融合报告基因定位法是研究蛋白亚细胞定位最广泛使用的方法之一[27]。洋葱鳞茎内表皮细胞因其具有不含叶绿体、结果容易观察等诸多优点而被广泛用作亚细胞定位的受体细胞[28]。本研究通过在线的BaCelLo对GmMYB173的亚细胞定位预测, 显示其定位于细胞核中(图1)。进一步利用基因枪介导洋葱表皮细胞瞬时转化实验验证预测结果, 实验结果与预测结果一致(图5)。通过氨基酸序列比对分析, 发现GmMYB173与GmMYB176相似, 都存在一个潜在的14-3-3蛋白结合位点, 即pST结合位点, 氨基酸序列为SSLFDITT[23]。进一步利用SOE-PCR技术删除相关编码序列发现GmMYB173失去了细胞核表达特异性(图5), 这一结果与GmMYB176的亚细胞定位结果[23]类似。转录因子需要在细胞核中实现对靶基因的精准调控[29], 由此该结合位点不仅影响了GmMYB173的亚细胞表达特性, 而且可能影响到GmMYB173功能的发挥, 暗示14-3-3蛋白可能通过pST结合位点影响GmMYB173亚细胞定位进而调控GmMYB173的功能。
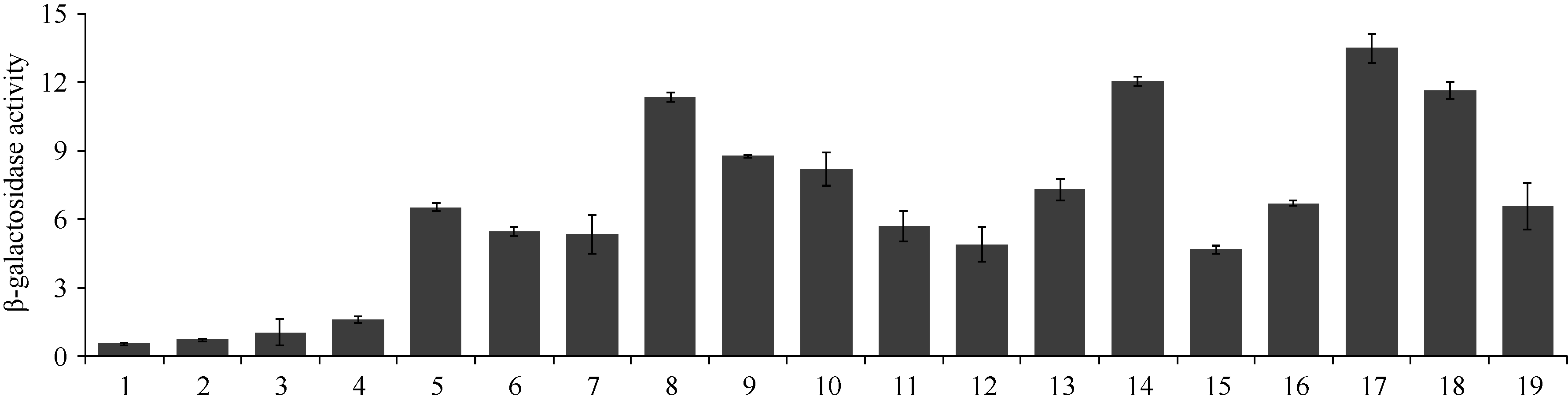 | 图10 β -半乳糖苷酶活性测定Fig. 10 Identification of β -galactosidase activity1: AD/BK; 2: GmMYB173/AD; 3: SGF14d/BK; 4: GmMYB173/SGF14a; 5: GmMYB173/SGF14b; 6: GmMYB173/SGF14c; 7: GmMYB173/SGF14d; 8: GmMYB173/SGF14e; 9: GmMYB173/SGF14f; 10: GmMYB173/SGF14g; 11: GmMYB173/SGF14h; 12: GmMYB173/SGF14i; 13: GmMYB173/SGF14j; 14: GmMYB173/SGF14k; 15: GmMYB173/SGF14l; 16: GmMYB173/SGF14m; 17: GmMYB173/SGF14n; 18: GmMYB173/SGF14o; 19: GmMYB173/SGF14p. |
为进一步明确14-3-3蛋白是否能够与GmMYB173互作, 本研究采用酵母双杂交方法对此分析, 该方法是研究蛋白之间相互作用的经典方法。利用此方法发现16个14-3-3蛋白与GmMYB173均能互作, 通过测定蛋白互作后激活的β -半乳糖苷酶活性发现它们与GmMYB173互作能力各有不同, 其中GmSGF14n与GmMYB173的结合活性最强, GmSGF14k、GmSGF14e和GmSGF14o稍次之, 这一方面表明在植物体中可能是GmSGF14n与GmMYB173结合进而调控靶基因的功能, 另一方面也说明植物有可能通过不同14-3-3蛋白实现对GmMYB173的精准调控。在对GmMYB176的研究中也发现16个14-3-3蛋白与GmMYB176均能产生互作, 其中GmSGF14i与GmMYB176互作激活的β -半乳糖苷酶活性最高[24], 进一步表明14-3-3蛋白有可能广泛参与MYB转录因子功能活性的调控, 但对于不同的MYB转录因子又呈现一定的结合特异性。
转录因子GmMYB173是通过酵母单杂交技术, 以自贡冬豆中开花基因GmFT2a的启动子的T1元件为诱饵筛选出来的[25]。GmFT2a在大豆叶片中表达进而促进茎尖端花分生组织决定基因的表达, 最终促进大豆开花[30]。Li等[7]研究发现大豆中14-3-3蛋白表现不同的亚细胞分布和组织表达特异性, 其中GmSGF14n被定位于细胞核中, 并且在大豆叶片中的表达量最高。本研究也发现GmMYB173定位于细胞核中, 并且GmSGF14n与GmMYB173互作强度最高。由此推断二者的互作可能会对GmFT2a的表达产生一定的影响, 但是仍需进一步实验验证。
由此可见, 14-3-3蛋白不仅与GmMYB173互作, 而且可能对其在亚细胞中的定位产生影响, 也就会对其功能的发挥产生一定的影响, 这一研究结果有助于今后更好地研究14-3-3蛋白与GmMYB173的互作关系及其在大豆生长发育中的作用, 促进大豆育种应用研究。
GmMYB173在细胞核中特异性表达, 这种特异性与14-3-3蛋白结合位点即pST结合位点有关。GmMYB173可与大豆中所有检测到表达的16个14-3-3蛋白GmSGF14a-GmSGF14p互作, 其中与GmSGF14n互作最强。由此可见, 14-3-3蛋白不仅与GmMYB173互作, 而且可能通过pST结合位点对其在亚细胞中的定位产生影响。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|