
{kind=link}
{kind=link}
{kind=link}
利用水稻MAGIC群体关联定位白叶枯病抗性QTL和创制抗病新种质
[陈天晓1, 2 , 朱亚军3 , 密雪飞3 , 陈凯3 , 孟丽君3 , 左示敏1, *  , 徐建龙
, 徐建龙2, 3, 4, * ]
, 徐建龙]
|
|


第一作者联系方式: E-mail: chentx2006@163.com, Tel: 010-82105855
以8个不同亲本构建的遗传上相互关联的多亲本高代互交系(multi-parents advanced generation inter-cross, MAGIC)群体, 包括2个4亲本群体(DC1和DC2)和1个八亲本群体(DC3)为材料, 接种我国白叶枯病强致病力V型菌系(GD-V)和弱致病力II型菌系(C2), 关联分析定位MAGIC群体对白叶枯病的抗性QTL, 筛选抗病种质。结果表明, 大多数亲本对C2菌系表现抗病, 而对GD-V菌系表现感病, 3个MAGIC群体的病斑长度均出现超亲分离。共检测到7个白叶枯病抗性QTL, 大多表现数量抗性, 而且抗性QTL表达存在明显的遗传背景效应。 QBbr11-1和 QBbr11-2受遗传背景影响较小, 具有一定的育种应用价值。从3个群体筛选出8份不同抗病QTL聚合的抗病材料, 表明质量抗性基因和水平抗性数量性状位点的结合可以显著提高抗性水平。8份不同抗病QTL的聚合系可以用作抗病育种的中间抗源。研究结果表明, MAGIC群体可以将遗传研究和育种应用有机结合, 是遗传研究和开展标记辅助育种的理想群体。
, XU Jian-Long
Three genetically interconnected multi-parents advanced generation inter-cross (MAGIC) population, including two populations (DC1 and DC2) derived from four parents and one population from eight parents (DC3) were used to detect QTLs for resistance to two strains, a weak virulent C2 and a strong virulent GD-V of Xanthomonas oryzaepv. oryzae( Xoo) and to screen resistant breeding materials. Most parents were resistant to C2 and susceptible to GD-V. Transgressive segregations of lesion length for the two strains were observed in the three MAGIC populations and showed continuous distributions. A total of seven QTLs affecting lesion length of two strains were detected. Most QTLs showed quantitative resistance and obvious genetic background effect. Among the seven QTLs, QBbr11-1and QBbr11-2 had less genetic background effect, which is valuable in rice breeding for disease resistance. Eight resistant lines pyramiding different QTLs were screened from the three MAGIC populations, indicating the combination of qualitative resistance gene and quantitative resistance gene can significantly improve resistance level. The eight resistant breeding lines could be used as resistant donors in rice breeding for resistance. The results indicated that the MAGIC populations are ideal material for genetic study and marker-assisted breeding, showing a tight integration of genetic research and breeding application in rice.
由黄单胞杆菌水稻致病变种(Xanthomonas oryzae pv.oryzae, Xoo)引起的水稻白叶枯病是一种造成稻米产量损失的细菌病害。最早在日本福冈地区, 之后陆续在世界主要水稻产区发现该病, 当前已经成为我国水稻生产中的三大病害之一[1, 2]。实践证明, 挖掘并利用抗性基因培育抗病品种是防治水稻白叶枯病最经济有效的手段[3]。
白叶枯病菌与水稻抗病基因之间存在典型的特异性互作, 水稻的R基因与病原菌无毒基因(Avr)产物之间直接或间接相互作用产生的“ 基因对基因” 抗性, 是水稻对白叶枯病抗性表现的重要形式[4], 同时被微效多基因控制的水平抗性也在水稻白叶枯病抗性中发挥重要作用[5]。迄今, 经国际注册确认和公开报道的水稻白叶枯病抗性基因已达30多个[6]。多数白叶枯病抗性基因由于抗谱狭窄或为隐性基因未被直接应用于抗病育种实践, 仅Xa3、Xa4、xa5、Xa7、xa13、Xa21、Xa23等少数抗白叶枯病基因得到较广泛的应用[7, 8], 在一定程度上降低了白叶枯病带来的产量损失。然而, 根据“ 基因对基因” 假说[9, 10], 在抗病基因的选择压下, 致病菌将不断进化, 使得某些抗性主效基因逐渐失去抗性。因此, 发掘新的抗性基因培育抗病品种将是抗病育种研究永恒不变的研究方向。在白叶枯病抗病遗传育种领域, 除直接利用抗性基因的策略外, Li等[11]也提出将垂直抗性基因和多个微效基因聚合以期获得水稻对白叶枯病的持久抗性的策略。
起源于不同地域的种质资源中往往隐藏着丰富的有利等位基因[12]。先前, Meng等[13]以8个来自国际水稻研究所、中国和其他国家的优良种质为亲本, 构建了3个相互关联的MAGIC群体, 成功开展了重要农艺性状的QTL定位研究。在本研究中, 我们对这3个MAGIC群体接种2个致病力不同的水稻白叶枯病菌, 开展了水稻抗白叶枯病QTL的定位研究, 并筛选出部分抗病中间材料。研究结果为发掘白叶枯病抗性新基因和培育抗白叶枯病水稻新品种提供了相关信息和材料基础。
3个MAGIC群体是利用来自不同育种项目的8个多样性丰富的优良种质为亲本构建的, 包括2个四亲本群体和1个八亲本群体。8个亲本分别是SAGC-08 (A)、HHZ5-SAL9-Y3-Y1 (B)、BP1976B-2- 3-7-TB-1-1 (C)、PR33282-B-8-1-1-1-1-1 (D)、FFZ1 (E)、CT 16658-5-2-2SR-2-3-6MP (F)、IR 68 (G)和IR 02A127 (H), 其来源及特点见表1。将8个亲本分成ABCD和EFGH 2组, 2组四亲本间两两成对杂交(A× B、C× D 和E× F、G× H), 产生4对双亲本杂交种, 将4对双亲本杂交种F1再成对杂交产生2种四亲本杂交种(A/B//C/D 和E/F//G/H), 2种四亲本杂交种通过单粒传法加代, 自交6代, 建成2个四亲本MAGIC群体, 获得221个稳定株系的A/B//C/D群体(记作DC1)和241个稳定株系的E/F//G/H群体(记作DC2)。分别从四亲本杂交种A/B//C/D和E/F//G/H中选25个F1单株, 通过互交产生八亲本杂交种(A/ B//C/D///E/F//G/H), 经过单粒传法加代, 建成包含455个稳定株系的八亲本MAGIC群体(记作DC3)。
接种鉴定试验在中国农业科学院作物科学研究所北京昌平实验基地完成。2015年5月1日播种, 5月27日单本移栽, 株行距为20 cm × 17 cm。田间按2重复随机区组设计, 每系每重复种植2行, 每行6株。全生育期田间采用常规管理, 仅治虫, 不防病。
用于接种的白叶枯病菌为2个遗传稳定的菌系GD-V (强致病病菌)和C2 (弱致病菌)。首先将保存于-80℃的甘油中的菌种在PSA培养基上复壮, 挑取单菌落经毒力测试后保存于4℃。然后将筛选过的有毒菌株再培养于PSA培养基上于28℃培养48 h, 进而配制成1× 108~1× 109cfu mL-1细菌悬浮液, 并于分蘖末期采用Kauffman剪叶法接种[14]。规定每个株系第1行的6株统一接种强致病菌GD-V, 第2行的6株接种弱致病菌C2; 从每株选取3~4张完全展开的叶接种。接种后保持田间有适度水, 以促进病斑扩展。待充分发病且病情稳定时(20 d)调查病斑长度。调查每重复6株, 测量每株3张感病最长的叶片, 以这3个表型值的平均值代表所调查单株的病斑长度, 每个株系的表型值为单个重复中所有单株病斑长度的均值。病斑长度< 1 cm为高抗、1~5 cm为抗病、5~10 cm为中抗、10~15 cm为中感、> 15 cm为感病。
| 表1 用于构建MAGIC群体的品系来源及其主要特征 Table 1 Origin and agronomic characteristics of eight founder lines used in developing the MAGIC population |
DC1、DC2和DC3群体的DNA提取和基于Illumina Infinium技术的Rice 6K SNP芯片基因型分析在国际水稻研究所(International Rice Research Institute, IRRI)的基因型分析实验室完成。Rice 6K SNP芯片由美国康奈尔大学Susan McCouch博士实验室完成。按以下3个步骤过滤SNP基因型数据, 一是所有杂合基因型作为缺失数据, 删除缺失10%以上标记数据的株系; 二是剔除所有稀有等位基因频率低于3%的标记; 三是删除标记间相关性很高(在0.95以上)的关联标记。最终在DC1、DC2和DC3群体中分别得到907、892和1329个高质量SNP位点用于群体结构分析和QTL定位研究。
使用TASSEL V5.2.3软件[15]评估群体结构和检测位点间的连锁不平衡(linkage disequilibrium, LD)水平。群体结构由主成分分析(principal components analysis, PCA)中的PC1和PC2来揭示。使用r2来衡量LD程度, 计算出所有可能位点组合的r2, 选用极显著(P < 0.001)的位点对, 绘制LD衰减散点图, 并绘制拟合曲线。以LD衰减至起始值一半时所对应的物理距离作为LD衰减距离[13]。
使用SAS V9.2 (SAS Institute Inc., Cary NC, USA) PROCGLM分析不同基因型、不同菌系及菌系与基因型互作在不同群体中的方差组成, PROCCORR对不同菌系病斑长进行相关分析。利用TASSEL V5.2.3软件中的MLM (Mixed Linear Model)程序对SNP标记与表型值进行全基因组关联分析, 以P < 0.001为阈值确定与白叶枯抗性显著关联的标记位点。
从8个亲本对2个菌系的抗性差异比较(表2)可以看出, 无论是接种强致病菌还是弱致病菌, 8个亲本间的抗性差异都达到极显著水平。接种弱致病力菌系C2, 除亲本B和E外, 其余亲本都表现抗病。接种强致病力菌系GD-V, 8个亲本对白叶枯病的敏病程度均显著增加, 但亲本D和G仍然表现中抗水平。总体而言, 来自中国的3个品种(A、B、E)对这2个致病菌系的抗性相对较差, 而来自东南亚和哥伦比亚的品种对2种菌株的抗性都相对较好。
对于3个MAGIC群体, 表型分析结果表明接种2个菌系各群体的病斑长度都表现出超亲分离, 呈连续变异(表3和图1)。因用于构建DC2群体及DC3群体的亲本E中感C2菌系, 接种C2菌系后DC2和DC3群体的平均病斑长度显著大于DC1群体; 接种菌系GD-V, 不同群体的整体表现较为一致。从3个群体病斑长度的频率分布中可以看出接种C2菌系引起的病斑长度分布存在较高的峰, 表明各群体对C2菌系的抗性受主基因控制, 但感病株系呈较大变异, 说明存在微效多基因的分离; 接种GD-V型菌系, 3个群体病斑长度呈连续变异, 出现抗、感双向均存在大幅度超亲分离, 表现为典型的数量性状遗传。
| 表2 亲本对不同菌系的抗性差异比较 Table 2 Comparison of resistance to the Xoo strains among eight parents |
| 表3 MAGIC群体接种白叶枯病菌后的病斑长度 Table 3 Lesion length (cm) of the MAGIC populations after inoculating two Xoo races |
方差分析结果表明(表4), 重复间差异不显著, 不同菌系、不同基因型及菌系与基因型的互作均达到极显著水平, 它们在DC1群体中分别解释表型变异的66.04%、17.56%和11.95%, DC2群体中分别为45.78%、38.02%和10.98%, DC3群体中分别为43.11%、39.24%和14.47%。相关分析表明, 接种强弱2个致病菌后, 相同群体的病斑长度存在极显著正相关, DC1群体接种C2和GD-V的相关系数为0.69, DC2群体为0.77, DC3群体为0.79, 表明多数株系对这2个菌系的抗性表现比较一致。
主成分分析结果表明, DC1群体的PC1和PC2分别是6.4%和5.0%, DC2群体的PC1和PC2分别是13.1%和3.0%, DC3群体的PC1和PC2分别是8.6%和3.2%。所有群体没有表现出明显的群体结构。
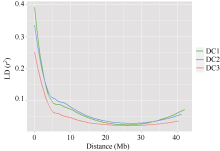
连锁不平衡(LD)分析显示3个群体的r2临界值分别为0.14、0.20和0.08, 均表现出随物理距离(Mb)增加而下降的趋势, 且DC3群体LD衰减幅度比DC1和DC2群体快(图2)。以LD降低至原始值的50% (r2> 0.2)作为LD的阈值, 3个群体LD衰减至一半的物理距离分别为2.50、2.50和1.25 Mb。从r2临界值和LD衰减幅度上来看, 8个亲本群体DC3的连锁不平衡程度最小, 衰减距离最短, 衰减速度最快, 而2个四亲本群体DC1和DC2的连锁不平衡表现趋于一致(图2)。
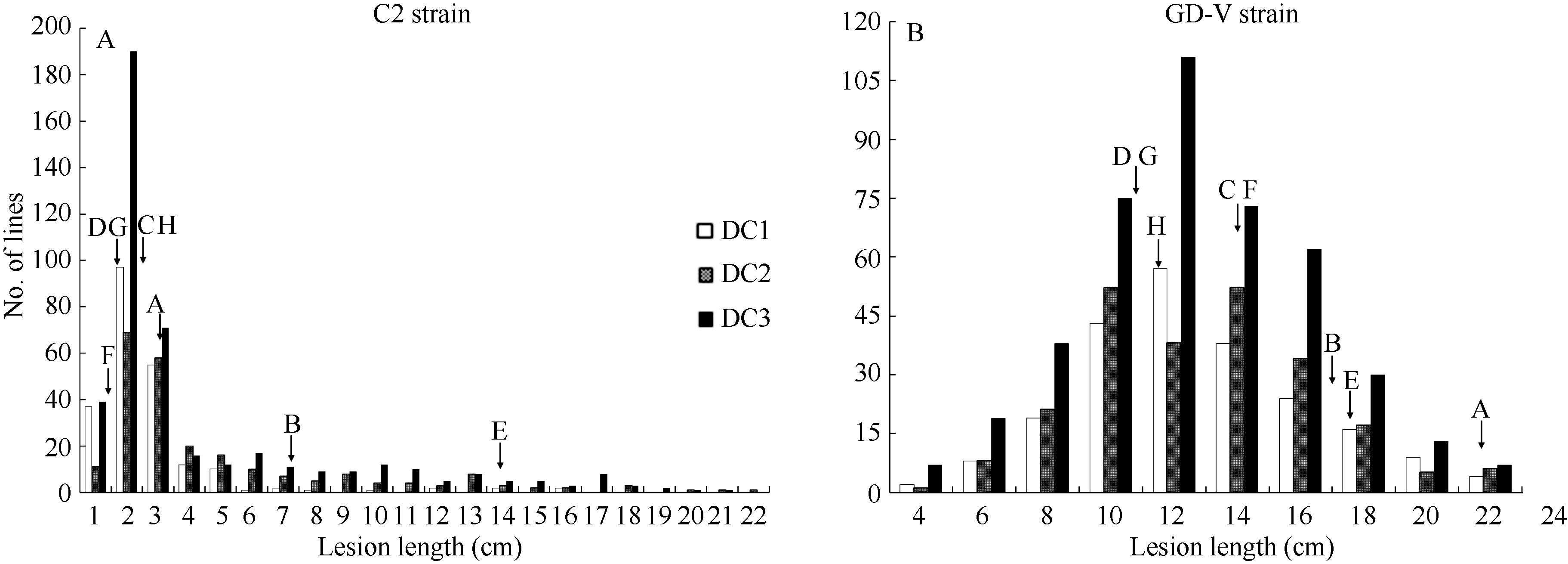 | 图1 MAGIC群体接种C2 (A)和GD-V (B)菌株的病斑长度频率分布亲本A~H的代号见表1。Codes of A-H represent varieties given in Table 1.Fig. 1 Frequency distributions of lesion length caused by Xoo strain, C2 (A) and GD-V (B) in the MAGIC populations |
| 表4 MAGIC群体接种不同菌系病斑长度的方差组成 Table 4 Variance component estimates for multiple factors of lesion length caused by Xoo in the MAGIC populations |
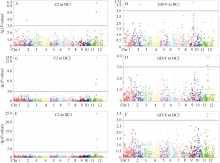
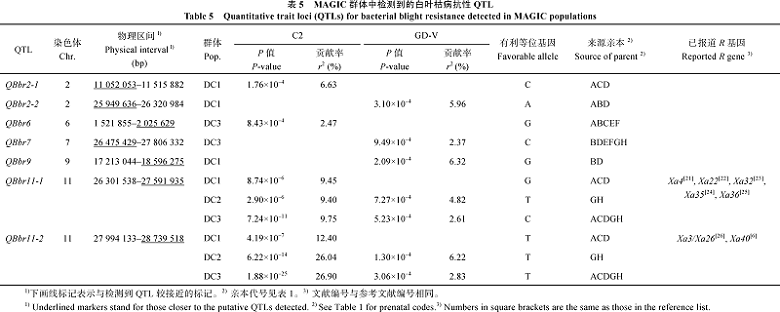
3个群体中共检测出7个抗白叶枯病QTL, 分布在第2、第6、第7、第9和第11染色体上(图3和表5)。接种C2菌系, DC1群体共检测出3个QTL, 即QBbr2-1、QBbr11-1和QBbr11-2, 分别解释表型变异的6.63%、9.45%和12.40%; DC2群体检测出2个QTL, 即QBbr11-1和QBbr11-2, 分别解释表型变异的9.40%和26.04%; DC3群体共检测出3个QTL, 即QBbr6、QBbr11-1和QBbr11-2, 分别解释表型变异的2.47%、9.75%和26.90%。接种GD-V菌系, DC1群体检测出2个QTL, 即QBbr2-2和QBbr9, 分别解释表型变异的5.96%和6.32%; DC2群体检测出2个QTL, 即QBbr11-1和QBbr11-2, 分别解释表型变异的4.82%和6.22%; DC3群体检测出3个QTL, 即QBbr7、QBbr11-1和QBbr11-2, 分别解释表型变异的2.37%、2.61%和2.83%。
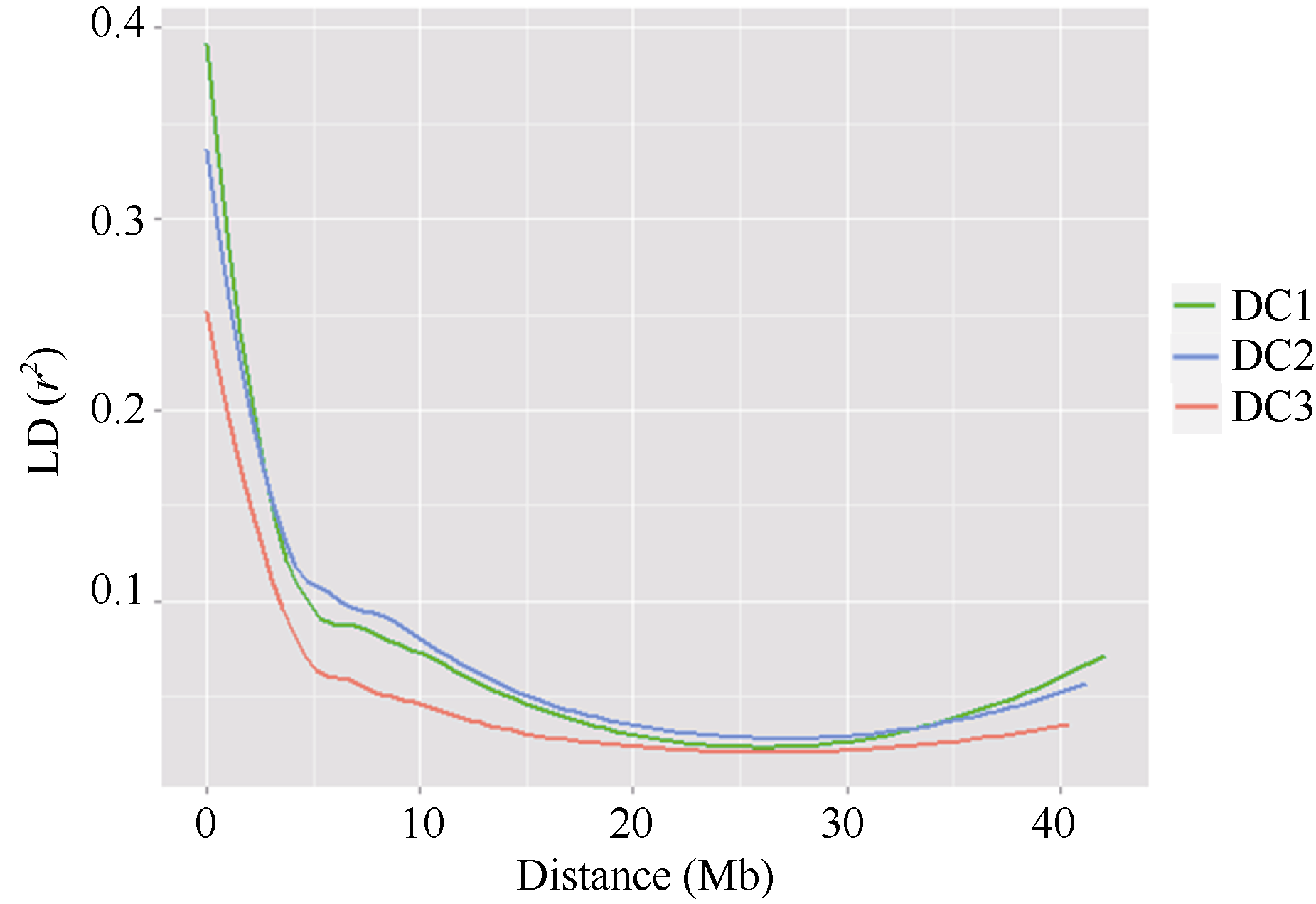 | 图2 3个MAGIC群体的LD衰减Fig. 2 LD (r2) decay of marker-pairs over all chromosomes as a function of physical distance (Mb) for three MAGIC populations |
所有被检测的QTL中, 只有QBbr11-1和QBbr11-2在不同群体中被检测到对不同菌系都表现抗性, 但抗性效应存在显著差异。显然QBbr11-2对弱毒性C2菌系表现出主基因抗性, 但对强毒性GD-V表现出数量抗性, 表明抗性QTL存在小种专化性, 这种专化性具体表现在同一抗病QTL对不同菌系的抗性反应不同及对不同菌系抗性效应存在差异。QBbr11-2在不同群体中对不同菌系均能被检测到且效应都最大, 该位点的有利等位基因来自亲本A、C、D、G和H。
 | 图3 全基因组关联分析定位影响MAGIC群体对2个白叶枯病菌的抗性QTLA、C、E表示混合线性模型下DC1、DC2和DC3群体中与C2菌系抗性显著关联标记的曼哈顿散点图; B、D、F表示混合线性模型下DC1、DC2和DC3群体中与GD-V菌系抗性显著关联标记的曼哈顿散点图。蓝色实线表示全基因组关联分析中的显著性阈值。Fig. 3 Genome-wide association analysis of QTLs underlying resistance to two bacterial blight strains in MAGIC populationsA, C, E: Manhattan plots (MLM) showing genome-wide association study (GWAS) with bacterial blight resistance to strain C2 in DC1, DC2, and DC3 MAGIC populations, respectively. B, D, F: Manhattan plots (MLM) showing GWAS with bacterial blight resistance to strain GD-V in DC1, DC2, and DC3 MAGIC populations, respectively. Blue solid line indicates the genome-wide significance threshold. |
2个四亲本群体DC1和DC2, 尽管它们的亲本数目相同、群体大小和标记密度相近, 但亲本的遗传差异较大导致群体间的遗传差异也较大。DC1群体共检测出5个QTL, DC2群体共检测出2个QTL, 其中QBbr11-1和QBbr11-2在2个群体中都被检测到。接种C2菌系, QBbr11-2在DC2群体中解释表型变异的26.04%, 为DC1群体(12.40%)的2倍多; 接种GD-V菌系, QBbr11-1和QBbr11-2只在DC2群体中被检测到, 表型贡献率相似, 而在DC1群体中均未被检测到。相似地, 八亲本群体相对四亲本群体亲本数目加倍, 群体大小也接近2倍。比较四亲本群体和八亲本群体接种C2菌系的定位结果发现, QBbr11-1和QBbr11-2在DC1、DC2和DC3群体中均被检测到, QBbr11-1在3个群体中的表型贡献率相似, 但QBbr11-2在DC1群体中的表型贡献率几乎是DC2和DC3群体中的一半。接种GD-V菌系时只在DC2和DC3群体中检测到QBbr11-2, 对表型贡献率均较小, 而在DC1群体中未检测到该QTL。表明抗病QTL的表达具有明显的遗传背景效应, 具体体现在同一抗病QTL在不同群体中被检测的效果不同, 而且表型效应也存在明显差异。
根据3个群体对强致病菌GD-V的抗性, 从3个群体中共筛选出8份抗病材料, 其中从DC1和DC2群体各筛选出1份, DC3群体筛选出6份(表6)。这些材料接种弱致病力菌系C2的病斑长度除L1602和L1959外均小于1 cm, 达到高抗水平; 接种强致病力菌系GD-V的病斑长度均小于4 cm, 达到抗病水平。通过对比这些株系携带的抗病位点可以看出, 同一遗传背景下如DC3群体, 不同株系的抗性水平与携带的抗病QTL数目之间具有较高的一致性, 而且带有第11染色体QBbr11-2主基因的株系抗性较强, QBbr11-1与QBbr11-2互作表现出较强的抗性效应。表明对弱毒菌系通过主效抗病QTL与微效QTL的累加, 或对强毒菌系通过不同微效QTL的累加, 都可以显著提高寄主对白叶枯病菌的抗性水平。
| 表5 MAGIC 群体中检测到的白叶枯病抗性QTL Table 5 Quantitative tralt locl(QTLs) for bacterial blight resistance detected in MAGIC populations |
| 表6 MAGIC群体中筛选出抗病株系的抗性及抗病QTL的分布 Table 6 Lines exhibiting resistance to bacterial blight screened from MAGIC populations and distributions of resistant QTLs |
与传统的双亲本群体如RIL相比, MAGIC群体中更多的亲本数量增加了等位基因多样性, 大量累积的重组事件会提高被检测QTL准确性, 并提高作图分辨率[16, 17]。与种质资源等自然群体相比, MAGIC群体降低了假阳性的出现并提高稀有等位基因检测的效率。利用MAGIC群体定位QTL还具有三项优势: (1)相比传统双亲本定位群体, MAGIC群体中包含来源于不同亲本的多个等位基因, 可以同时探讨多个等位基因对某个性状的影响。(2)以前通过双亲群体定位的很多QTL在离开了特定的遗传背景后, 对相应的农艺性状无效。MAGIC群体由不同亲本来源的多个群体构成, 比较不同遗传背景群体的定位结果, 就能发现受亲本背景影响较小的QTL, 使QTL的效应能够更加确定和具有通用性。如本研究中定位出的QBbr11-1和QBbr11-2在不同群体中均被检测到, 说明这2个QTL受遗传背景影响小, 可靠性较高, 可以用于进一步研究。(3)构建MAGIC群体可选用育种项目中性状优异的材料作为亲本, 多次重组创造大量的遗传变异, 群体中出现的优良株系可用做育种中间材料或直接选育品种, 定位到的QTL可以直接指导育种, 真正做到育种群体和定位群体的有机整合[18, 19, 20]。本研究筛选了8份抗病株系, 聚合多个抗病QTL, 不但可以用于进一步的遗传研究, 还具有较高抗病育种价值。我们已
经将部分抗病材料与感病材料配组, 构建主效抗病QTL的精细定位群体, 以期挖掘新的抗病基因或验证本研究定位到的主效抗病QTL与已往报道的抗病基因或QTL的关系。
不可否认, 由于本研究所用芯片通量不高, 造成在每个群体中呈现多态的标记数量有限, 因而无法有效实现抗病位点的有利等位基因的剖析, 使得多亲本群体在基因定位上的优越性受到一定程度的制约。有鉴于此, 我们正有利用3K重测序种质资源开发的56K芯片, 重新鉴定这3套多亲本群体SNP基因型, 以实现基因或QTL的精细定位。
根据与定位到的QTL显著关联SNP标记的物理位置, 我们将本研究定位到的抗水稻白叶枯病QTL与前人的定位结果比较, 发现第11染色体QBbr11-1所在区域26 301 538~27 591 935存在已被定位的水稻白叶枯抗性基因Xa4[21]、Xa22[22]、Xa32[23]、Xa35[24]和Xa36[25], 在QBbr11-2所在区域27 994 133~28 739 518存在Xa3/Xa26[26]和Xa40[6]基因。本研究结果表明QBbr11-1对弱毒力菌系C2和强毒力菌系GD-V均表现为一个微效基因, 而QBbr11-2对C2表现为质量抗性基因, 但对GD-V仍有显著的残效, 表现为一个加性效应的微效QTL。上述定位到的2个抗病QTL与所在区域已报道的白叶枯病抗性基因的关系, 还需加密标记精细定位后方能知晓。此外, 在第2、第6、第7和第9染色体上定位到的5个抗病QTL (QBbr2-1、QBbr2-2、QBbr6、QBbr7和QBbr9)所在区域, 未发现有抗白叶枯病基因或QTL的报道, 可能属于新发现的抗白叶枯病位点。
QTL表达的遗传背景效应在水稻抗旱[27, 28]、耐盐[29, 30]等性状上已有报道。在植物抗病基因表达上也存在遗传背景效应, Banerjee等[31]发现拟南芥抗细菌性斑点病菌(P. syringae)基因RPS2在Po-1遗传背景下不起作用, 但该基因在哥伦比亚(Col-0)品种背景下对细菌性斑点病菌的avrRpt2致病因子具有完全抗性。在水稻抗白叶枯病基因表达的遗传背景方面, Xa21[32]和Xa26[33]的表达受遗传背景影响, 认为这两个抗性基因在粳稻背景下比在籼稻背景下抗性表达更完全。本研究中, 同样2个四亲本群体DC1和DC2, 群体大小相似, 对C2和GD-V菌系DC1群体共检测出5个QTL, 而DC2群体只检测出2个QTL, 其中QBbr2-1、QBbr2-2和QBbr9只在DC1中被检测到, 未能在DC2群体中检测到。接种C2菌系, 尽管QBbr11-1和QBbr11-2在2个群体中都被检测到, QBbr11-2在DC2群体中的表型变异(26.04%)比在DC1群体(12.40%)中高出2倍多。同样, 接种GD-V菌系, QBbr11-1和QBbr11-2只在DC2群体中检测到, 在DC1群体中未检测到, 表明抗白叶枯病QTL存在明显的遗传背景效应。抗性基因表达存在遗传背景效应, 意味着抗性基因从一个背景转育到另一个完全不同的背景下, 其抗性水平可能会发生变化, 这种遗传背景效应在抗病育种中应引起足够的重视。
近几十年来, 利用来自双亲构建的重组自交系、DH系、双向导入系等群体定位了大量水稻抗白叶枯病QTL[34, 35, 36, 37], 但多数QTL效应不大, 有些与环境间有很强的互作, QTL表达的遗传背景效应明显, 育种应用价值较小。一直以来, 对水稻白叶枯病的遗传研究和抗性育种主要集中在主基因抗性[37]。迄今虽然定位到了30多个质量抗性基因, 但抗病育种大多集中利用Xa4、Xa21、Xa23等少数广谱抗性基因[7, 38]。这种抗性基因的集中利用会导致白叶枯病菌的消长, 产生新的致病菌系, 造成对带有Xa4基因的品种在中国和带有Xa21基因的品种在菲律宾、韩国、印度和中国丧失抗性[39, 40, 41]。因此, 发掘新的抗性基因或将不同抗性基因累加是提高品种的抗性水平或延长抗病品种使用年限的重要举措。本研究表明, 水稻对白叶枯病存在主基因抗性和多个微效基因抗性, 它们共同影响抗性水平, 这与3个多亲本群体中观察到抗性的极端超亲分离相符合。鉴于水平抗性数量化及抗病QTL小种专化性弱的特点, 利用标记辅助选择方法借助紧密连锁的分子标记进行质量抗性基因和水平抗性QTL的组合, 可能是提高品种抗性水平的有效措施[42]。本研究定位了来自不同亲本的7个抗性QTL, 大多表现为数量抗性, 具有明显的抗性聚合效应。将筛选出的8个不同抗病QTL的聚合株系作抗性供体, 通过分子标记辅助选择将这些抗性基因导入到优良品种背景, 可以培育出符合育种目标的抗病品种。
在遗传上相互关联的2个四亲本群体和1个八亲本群体中观察到对白叶枯病弱毒菌系C2和强毒菌系GD-V的抗性超亲分离。共检测到影响水稻白叶枯病抗性的7个QTL, 大多QTL均表现数量抗性, 抗性表达存在明显的遗传背景效应。QBbr11-2对C2表现质量抗性, 对GD-V表现一定的抗性残余效应。QBbr11-1和QBbr11-2受遗传背景影响较小, 具有一定的育种应用价值。从3个群体筛选出8份不同抗病QTL聚合的抗病材料, 可用作抗病育种的中间抗源。本研究证实了水稻多亲本群体既是遗传研究群体也是理想的育种群体, 可以实现基因定位与育种应用的有机结合。
致谢: 感谢中国农业科学院作物科学研究所水稻分子遗传与分子育种实验室周永力老师和王明明同学帮助培养菌株, 张强、张建、王小倩等同学帮助田间接种和病斑调查。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|