

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
不同穗型超级稻品种籽粒灌浆特性
[魏颖娟, 赵杨, 邹应斌*  ]
]
]
|
|


第一作者联系方式: E-mail: yvonne_wei@qq.com
为探明不同穗粒型超级稻产量构成特点、籽粒灌浆特性及其与后期生理指标的关系。以不同生育期品种准两优527、两优培九、Y两优1号、玉香油占、黄花占为试验材料, 在大田栽培条件下分期播种以确保不同生育期品种在相似的温光条件下灌浆结实。结果表明, 不同播期平均产量及其构成因子的品种间差异显著, 以两优培九产量最高(9.47 t hm-2), 玉香油占产量最低(8.39 t hm-2)。依据每穗粒数和千粒重系统聚类分析, 可将品种分为大粒型(准两优527)、中粒型(两优培九、Y两优1号)和小粒型(玉香油占、黄花占)。籽粒灌浆强度及持续时间的品种间差异明显。其中, 大粒型品种籽粒灌浆起步快、强度大、峰值持续时间短, 上、中、下部枝梗籽粒均为一段式灌浆; 中粒型品种籽粒灌浆起步快、强度较小、峰值持续时间长, 其上部枝梗籽粒为一段式灌浆, 中、下部枝梗籽粒为两段式灌浆, 高峰值分别在抽穗后12~18 d和24~30 d; 小粒型品种籽粒灌浆起步慢、强度小、峰值持续时间长, 其上、中部枝梗籽粒为一段式灌浆, 下部枝梗籽粒为两段式灌浆。平均籽粒灌浆速率与品种的穗粒类型密切相关, 其中, 大粒型品种为0.68 mg d-1, 中粒型品种为0.48~0.51 mg d-1, 小粒型品种为0.41~0.47 mg d-1。籽粒灌浆强度及持续时间与后期剑叶、根系生理指标有关, 籽粒灌浆启动后, 叶绿素含量、叶绿素 a/ b值、类胡萝卜素含量逐渐降低, 剑叶可溶性糖含量、MDA含量逐渐上升, POD活性、CAT活性及根系活力先后出现2次升降过程。可见, 不同穗粒型超级稻品种产量构成及籽粒灌浆特点的差异明显, 籽粒灌浆强度及持续时间与品种的穗粒型及枝梗着生的部位有关, 也与后期剑叶及根系衰老生理指标的变化趋势一致。
The objective of this study was to ascertain the characteristics of the grain-filling and physiological indexes after heading and the yield components of super rice with different panicle-grain types. An experiment with different sowing date was conducted using different growth duration varieties (Zhunliangyou 527, Liangyoupeijiu, Y-liangyou 1, Yuxiangyouzhan, and Huanghuazhan) under the field conditions to ensure the grain filling was under the similar temperature and solar radiation conditions. The grain yield and its components averaged in three sowing treatments displayed significant difference among tested varieties, of which the highest yield reached 9.47 t ha-1 for Liangyoupeijiu and the lowest was 8.39 t ha-1 for Yuxiangyouzhan. According to the hierarchical cluster analysis based on the grain number per panicle and 1000-grain weight, tested varieties were divided into the big grain type (Zhunliangyou 527), the medium grain type (Liangyoupeijiu and Y-liangyou 1), and the small grain type (Yuxiangyouzhan and Huanghuazhan). There were obvious differences intensity and peak duration of grain filling among the tested varieties. The big grain variety showed quick grain growth at the beginning, high intensity and short peak duration for grain filling during the middle-late periods, being one step filling in the upper, middle and lower branch grains; the medium grain variety showed quicker grain growth at the beginning, smaller intensity and longer peak duration for grain filling during the middle- late periods, with two filling peaks from 12 d to 18 d and from 24 d to 30 d after heading, respectively; the small grain variety showed slow grain growth at the beginning, smaller intensity and shorter peak duration for grain filling during the middle-late periods, which was one step filling in the upper or middle branch grains and two step filling in the lower branch grains. The average grain filling rate closely related to the panicle-grain type of varieties, was 0.68 mg d-1 for big grain variety, 0.48-0.51 mg d-1 for medium varieties, and 0.41-0.47 mg d-1 for small grain varieties. The intensity and duration of the grain-filling also associated with the changes of physiological indexes of flag leaf and roots after heading. At the beginning of the grain-filling, chlorophyll content, chlorophyll a/ b ratio, carotenoids content gradually declined, leaf soluble sugar content, MDA content gradually increased, POD and CAT activities and root activity successively underwent two rise and fall processes. In conclusion, different panicle-grain types super rice have significantly different characteristics of yield components and grain filling. The intensity and the duration of grain-filling depend upon the panicle-grain type of varieties and the branch and grain positions inserted on the panicle, with the same tendency of physiological indexes variation in flag leaf and roots.
水稻是世界重要的粮食作物, 提高水稻单产对于确保粮食安全具有重要意义。不同穗粒型水稻产量构成特点及籽粒灌浆增长过程不同, 而籽粒灌浆速率及其后期生理指标是影响最终产量的关键因子。水稻籽粒灌浆速率及粒重与其稻穗上着生的部位关系密切, 前人的研究证明稻穗上部开花早的颖花为强势粒, 稻穗下部开花迟的颖花为弱势粒, 在灌浆过程中, 强势粒灌浆速率较快, 弱势粒灌浆速率较慢[1, 2, 3], 但也有报道指出水稻强、弱势颖花主要决定于颖花着生的位置, 而不完全决定于开花顺序[4]。大穗型水稻的强、弱势粒的灌浆速率差异明显, 存在“ 两段灌浆” 现象, 即强势粒灌浆启动早, 速度快, 待强势粒灌浆高峰过后弱势粒才开始灌浆[5, 6, 7]。这种水稻强、弱势粒的异步灌浆现象同样存在于密穗型籼稻[8]和弯曲穗型粳稻[9, 10]。水稻弱势粒灌浆速率慢的原因, 一是灌浆初期籽粒生理活性低, 导致灌浆启动慢[11, 12, 13]; 二是蔗糖转化为淀粉的效率低, 导致灌浆速率低[1, 14, 15]。研究还发现水稻籽粒灌浆启动后, 植株内部碳、氮代谢发生了变化, 叶片逐渐衰老, 叶绿素含量降低[16, 17, 18]。由于根系与叶片衰老密切相关, 根系衰老早于叶片, 根系衰老会加速叶片的衰老, 因此维持水稻后期较高的根系活力有利于促进籽粒灌浆结实[19, 20, 21, 22, 23]。超级稻, 尤其是超级杂交稻具有明显的大穗优势, 强弱势籽粒灌浆强度、达到最大灌浆速率的时间及灌浆持续时间差异明显, 且稻穗越大强弱势籽粒异步灌浆的特征越明显[24, 25, 26], 抽穗前贮存的非结构碳水化合物与籽粒灌浆强度密切相关[27, 28]。有关不同穗粒型超级稻与常规稻籽粒灌浆特性及其与后期生理指标变化的关系尚缺乏系统的比较研究, 本研究试图在大田栽培条件下, 采用分期播种的方法调节不同生育期品种在同期抽穗开花, 即在相同的光温条件下研究不同穗粒型超级稻籽粒灌浆特性及其与后期生理指标的关系, 为水稻高产栽培及品种选育提供理论依据。
以不同穗粒型超级稻品种准两优527 (ZLY527)、两优培九(LYPJ)、Y两优1号(YLY1)、玉香油占(YXYZ)和高产品种黄华占(HHZ)为试验材料, 种子分别由湖南杂交水稻研究中心和广东省农业科学院水稻研究所提供。
2011— 2012年在湖南农业大学试验基地(长沙)大田, 采用分期播种促使各不同生育期的参试品种能够在大致相同的日期抽穗开花和灌浆结实, 以便在相同的光温气候条件下测定各品种的籽粒灌浆动态及反映叶片、根系活力的生理指标。试验前土壤含有机质27.71 g kg-1、总氮1.60 g kg-1、无机氮40.92 mg kg-1、有效磷54.53 mg kg-1、有效钾63.2 mg kg-1、有效锌3.68 mg kg-1, pH 5.81。设置5月3日、5月13日、5月23日3个播期处理, 顺序排列于田间。同一播期不同品种按随机区组排列, 重复3次, 小区面积25 m2, 共45个小区。不同播期处理间作田埂, 以便对其分别施肥、灌溉和管理。各处理的肥料用量和施肥方法相同, 即氮肥150 kg N hm-2、磷肥75 kg P2O5 hm-2、钾肥150 kg K2O hm-2, 氮肥分基肥(50%)、分蘖肥(20%)、穗肥(30%) 3次施用。磷肥和钾肥作基肥, 在耙田前2 d一次施用。经250 mg kg-1咪鲜胺溶液浸种消毒, 催芽后播种, 播种量25 g m-2。其后泥浆踏谷, 湿润育秧。秧龄期25 d, 即分别于5月28日、6月7日、6月17日手工插秧, 密度20 cm × 20 cm, 每穴常规稻和杂交稻均栽插2苗。不同播期处理均按照高产栽培方法进行灌溉和病虫草害防治等田间管理。
1.3.1 籽粒灌浆速率测定 对于根系、叶片、籽粒灌浆测定有15个取样小区, 即抽穗期相近的播期作为取样区。选择抽穗日期大致相同的播期处理作为籽粒灌浆速率测定的取样区, 以抽穗50%的日期为抽穗期, 两年试验取样方法相同。两年抽穗日期稍有差异, 其中2011年5个品种同期抽穗的日期为8月15日至17日, 2012年为8月14日至16日。在各处理抽穗当天选择生长基本一致的稻穗挂牌标记, 并开始第1次取样, 以后每6 d取样一次, 到开花后30 d, 连续取样6次, 成熟期(开花后37~41 d)最后一次取样。在标记取样的试验小区选取10个大小基本一致的稻穗, 每个稻穗以上部3个一次枝梗籽粒作为上部枝梗籽粒, 下部3个一次枝梗籽粒作为下部枝梗籽粒, 其余为中部枝梗籽粒, 分别脱粒和计数, 在70℃恒温条件下烘干至恒重, 用1%的电子天平称籽粒干重。
1.3.2 剑叶和根系生理指标测定 取样日期与1.3.1相同。从每个试验小区, 随机选取除边3行外生长均匀且有代表性的5穴。全株(包括带泥根部)。简易清洗根部后带回实验室, 参照萧浪涛等[29]方法测定生理指标:
取一部分剑叶用液氮处理后低温贮存, 用于测定过氧化氢酶、过氧化物酶活性、丙二醛含量等生理指标。剑叶叶绿素含量为叶绿素a、b之和, 应用95%乙醇提取叶绿素, 分别在665 nm、649 nm和470 nm波长下测定吸光度计算叶绿素a、b含量; 硫代巴比妥酸比色法测定丙二醛(MDA)含量; 蒽酮比色法测定可溶性糖含量; 愈创木酚法测定过氧化物酶(POD)活性; 过氧化氢法测定过氧化氢酶(CAT)活性。
取全株根系用蒸馏水洗净, 吸水纸吸干, 采用α -萘胺法测定根系活力。
1.3.3 产量及其构成 取样区为全部45个小
区。于成熟期去除每个试验小区边3行稻株, 割125穴(5 m2)测定实收产量, 折算为13.5%含水量的产量。同时调查每小区20穴计算有效穗数, 取10穴人工脱粒, 用清水浸泡15 min, 以下沉的稻谷为实粒, 上浮的稻谷为空秕粒。晾干实粒和空秕粒后分别用1%的电子天平称重, 并从实粒中称取3份30 g样品。分别计数实粒样品和空秕粒样品, 在70℃下烘至恒重, 用1%的电子天平称重, 计算每穗总粒数、结实率和千粒重。
用Microsoft Excel 2007整理数据, DPS软件分析数据, LSD0.05法进行多重比较。
2.1.1 产量及其构成的年份间、播期间及品种间差异 从表1可以看出, 不同穗粒型超级稻的干物质重、收获指数、产量及产量构成因子的品种间差异显著或极显著。除了干物质重和单位面积颖花数的差异不显著外, 不同播期对收获指数、产量及产量构成因子的影响显著或极显著。不同年份间的产量、颖花数、千粒重差异极显著, 但干物质重、收获指数及其他产量构成因子的差异不显著。品种× 年份的互作对产量的影响显著, 对各产量构成因子的影响不显著; 年份× 播期的互作对各产量构成因子的影响显著, 对产量的影响不显著; 品种× 播期的互作对产量、有效穗数、每穗粒数的影响显著, 对其他产量构成因子的影响不显著。
2.1.2 不同播种期条件下的产量表现及生育期变化 从表2可以看出, 不同穗粒型超级稻产量及生育期存在年份间、播期间、品种间差异。不同品种平均产量随播期推迟而增加, 2011年第一、二、三播期分别为9.01、9.27、9.31 t hm-2, 差异不显著; 2012年分别为8.42、8.55、9.04 t hm-2, 处理间差异显著。不同播期平均产量品种间差异显著, 其中以两优培九最高, 2011年、2012年分别为9.92 t hm-2和9.03 t hm-2; 以玉香油占最低, 两年分别为8.44 t hm-2和8.34 t hm-2。表3表明平均产量2011年为9.20 t hm-2, 2012年为8.67 t hm-2, 年度间差异显著。值得注意的是, 同一块稻田种植同一品种, 在不同年份、不同播期的产量差异主要受生长期间, 尤其是灌浆结实期间光照、温度等气候因素影响。表2还表明, 每推迟10 d播种, 播种至抽穗日数缩短1~2 d, 全生育期缩短2~3 d。品种间全生育期差异明显, 以两优培九最长, 为136~141 d, 其余依次为Y两优1号(131~137 d)、准两优527 (127~133 d)、黄华占(122~128 d)、玉香油占(123~125 d)。全生育期年度间稍有差异, 2011年平均为129.3 d, 2012年为131.1 d。
| 表1 不同穗粒型超级稻产量表现、产量构成的年份、播期、品种间差异及其交互作用(F值) Table 1 Variance analysis of yield and its components of supper rice with different panicle-grain types (F-value) |
| 表2 不同播种期条件下不同穗粒型超级稻品种的产量表现 Table 2 Effect of sowing date on the yield performance of supper rice with different panicle-grain types |
2.1.3 不同播种期条件下的产量构成特点 从表3可以看出, 播种期对不同穗粒型超级稻产量构成因子有显著影响, 同一播期产量构成因子的品种间差异显著。按3个播期平均值计: 每平方米有效穗数以黄华占最多, 2011、2012年分别为312.7穗和319.5穗, 其余4个品种差异不显著; 每穗粒数以准两优527最少(分别为114.4粒和139.6粒), 其次为黄华占(分别为134.5粒和161.0粒), 其余3个品种每穗粒数差异不显著; 结实率以黄华占和准两优527较高, 分别为84.8%~88.4%和83.3%~84.6%, 其余品种差异不显著; 千粒重以准两优527最高(分别为29.3 g和29.6 g), 其次为Y两优1号和两优培九, 分别为23.4~23.7 g和23.8~24.3 g, 以黄华占和玉香油占最低(分别为19.9~20.4 g和19.4~19.7 g), 品种间差异显著。依据品种的每穗粒数和千粒重系统聚类分析, 将参试的5个品种划分为大粒型品种(准两优527), 中粒型品种(两优培九和Y两优1号), 小粒型品种(玉香油占、黄华占) 3种类型。
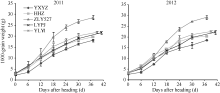
2.2.1 全穗籽粒增长动态 从图1可以看出, 不同穗粒型超级稻籽粒增长动态, 从抽穗开花当天到开花后6 d为渐增期, 从开花后6~30 d为快增期, 开花后30 d到成熟期为缓增期。渐增期大穗小粒品种(玉香油占)籽粒增长的起步慢, 其余4个品种起步快, 但增长速度的差异不明显; 快增期籽粒增长的品种间差异显著, 其中小穗大粒型品种(准两优527)增长最快, 大穗小粒型品种(玉香油占)增长最慢, 而大穗中粒型品种(两优培九)、中穗中粒型品种(Y两优1号)、中穗小粒型品种(黄华占)籽粒增长的差异不大; 进入缓增期籽粒增长减慢, 最终籽粒由大到小依次为准两优527、Y两优1号、两优培九、黄华占和玉香油占。
| 表3 播种期对不同穗粒型超级稻产量构成因子的影响 Table 3 Effect of sowing date on yield components of supper rice with different panicle and grain types |
 | 图1 不同穗粒型超级稻全穗的籽粒增长动态缩写同表2。Abbreviations are the same as those given in Table 2.Fig. 1 Grain growth dynamics of the whole panicle of supper rice with different panicle-grain types |
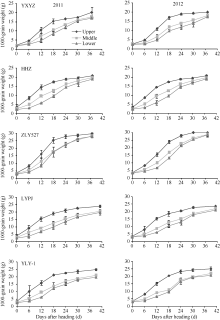
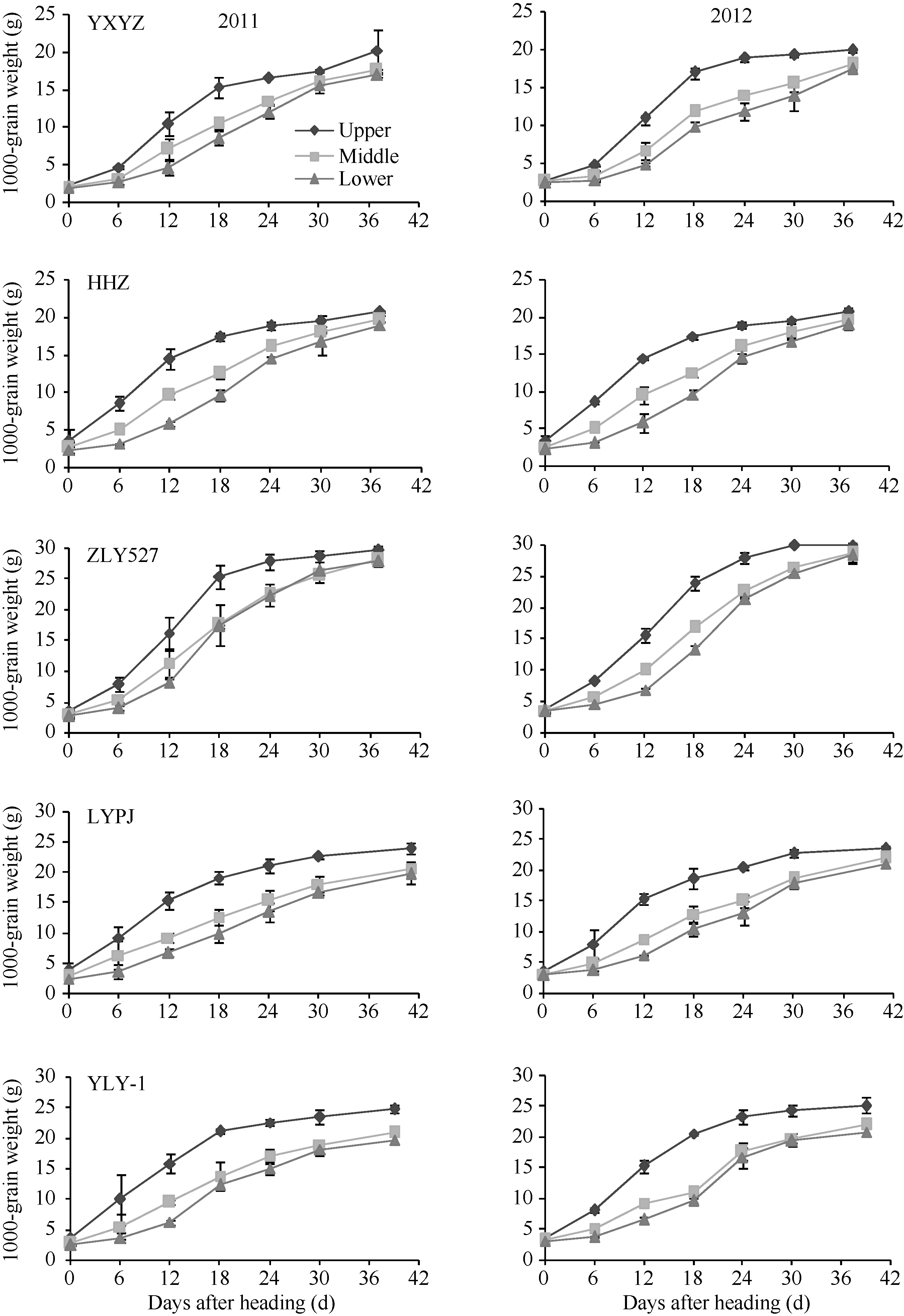
2.2.2 各部位枝梗籽粒增长动态 从图2可以看出, 不同穗粒型超级稻上、中、下部枝梗籽粒增长动态差异明显, 但增长动态的变化趋势品种间基本一致。不同穗粒型超级稻上部枝梗籽粒增长起步快, 从开花0 d到开花后18 d籽粒快速增长; 从开花后18 d到成熟期籽粒增长缓慢; 中部枝梗籽粒增长开始较慢, 开花后6~30 d快速增长; 下部枝梗籽粒增长较慢, 在开花后12 d才进入快速增长, 直到成熟期仍然增长较快。无论是上部、中部, 还是下部枝梗, 以小穗大粒型品种(准两优527)籽粒增长速度快, 大穗小粒型品种(玉香油占)籽粒增长速度慢, 其余3种类型品种(两优培九、Y两优1号和黄华占)籽粒增长速度差异不明显, 两年观测结果基本一致。
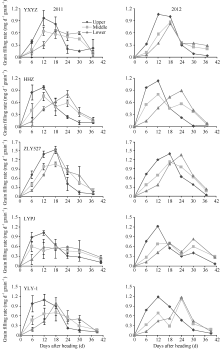
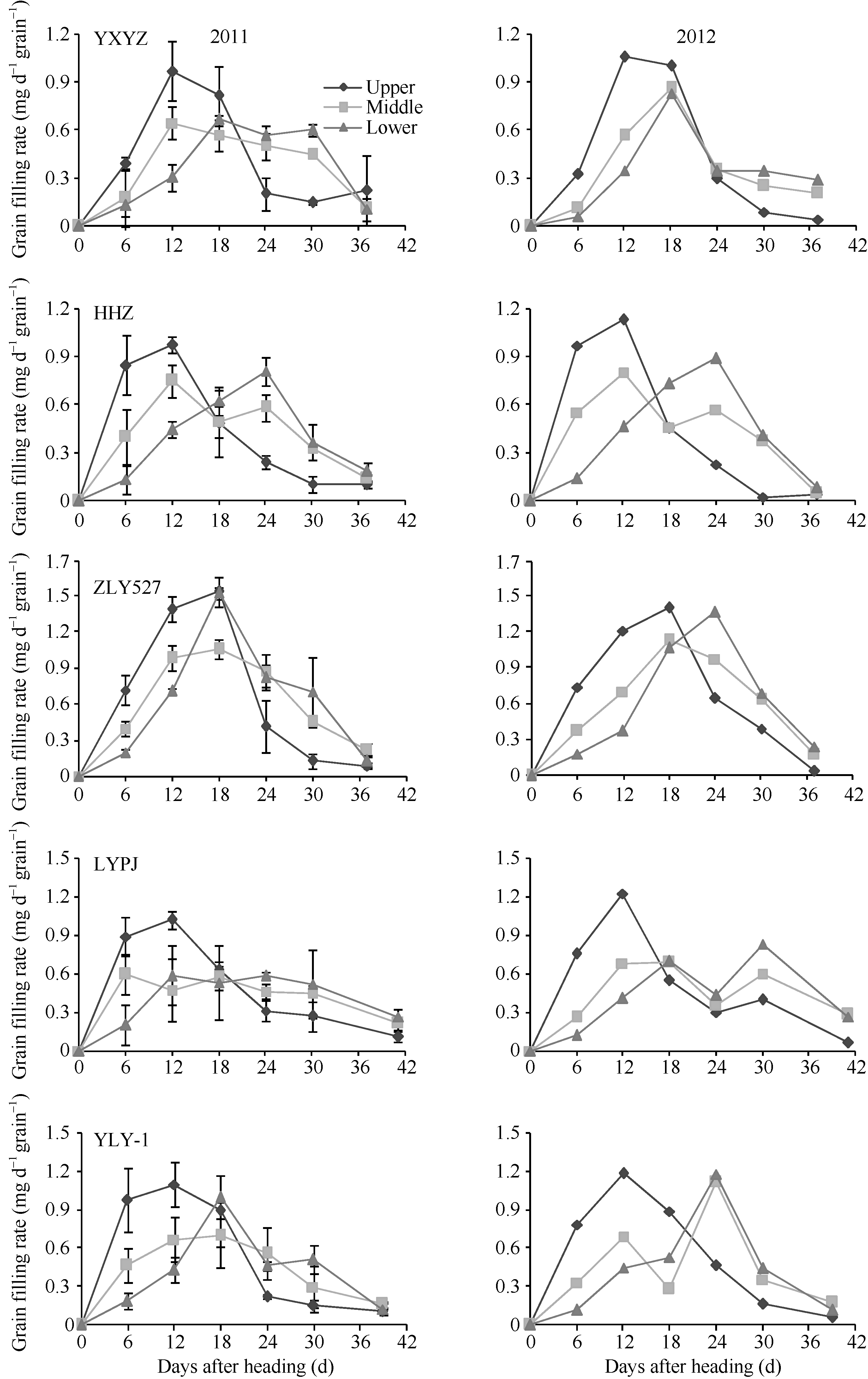
2.3.1 整穗籽粒灌浆速率变化 从图3和表4可以看出, 不同穗粒型超级稻灌浆速率差异显著。小穗大粒型品种准两优527灌浆速率快, 灌浆强度大, 在开花后12~18 d灌浆速率明显大于其他4个品种, 其中在开花后18 d达到灌浆高峰(1.26~1.19 mg d-1)。大穗小粒品种玉香油占灌浆起步慢, 开花后6 d灌浆速率开始快速增加, 到12~18 d达到灌浆高峰(0.67~0.90 mg d-1), 但高峰持续时间短, 高峰值后快速下降。中穗小粒型品种黄华占灌浆起步快, 开花后12 d达到灌浆高峰(0.74~0.83 mg d-1), 高峰持续的时间短, 高峰值后快速下降。大穗中粒型品种两优培九和中穗中粒型品种Y两优1号灌浆速率快, 开花后12~18 d出现第1个灌浆高峰, 但品种间差异较大, 分别为0.62~0.72 mg d-1和0.80~1.04 mg d-1, 且2011年的高峰值的持续时间长, 2012年在开花后24 d出现第2个灌浆高峰, 表现出两段式灌浆现象。从表4还可以看出, 超级稻籽粒平均灌浆速率与品种的穗粒型关系密切, 不同部位枝梗籽粒表现一致。整穗平均灌浆速率大粒型品种准两优527最高, 为0.68 mg d-1; 中粒型品种两优培九、Y两优1号中等, 分别为0.48~0.49 mg d-1和0.50~0.51 mg d-1; 小粒型品种玉香油占、黄华占最低, 分别为0.42~0.41 mg d-1和0.45~0.47 mg d-1。
2.3.2 不同部位枝梗籽粒灌浆速率变化 从图4可以看出, 超级稻不同部位枝梗籽粒灌浆速率差异显著, 并且不同穗粒型品种间表现一致。稻穗上部一次枝梗籽粒表现出一个灌浆高峰, 为一段式灌浆, 高峰出现在抽穗后6~18 d, 其灌浆强度大; 稻穗中、下部枝梗籽粒灌浆速率因品种穗粒型不同而异。其中, 小穗大粒型品种准两优527籽粒灌浆起步快, 灌浆强度大, 持续时间短, 上、中、下部枝梗籽粒均为一段式灌浆; 大穗中粒型或中穗中粒型水稻籽粒灌浆起步快, 灌浆强度小, 持续时间长, 其上部枝梗籽粒为一段式灌浆, 中、下部枝梗籽粒为两段式灌浆, 高峰值出现在开花后12~18 d和24~30 d; 大穗小粒型或中穗小粒型水稻籽粒灌浆起步慢, 灌浆强度小, 持续时间长, 其上、中部枝梗籽粒为一段式灌浆, 下部枝梗籽粒为两段式灌浆。
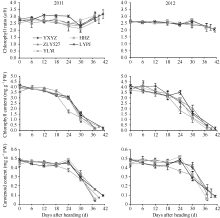
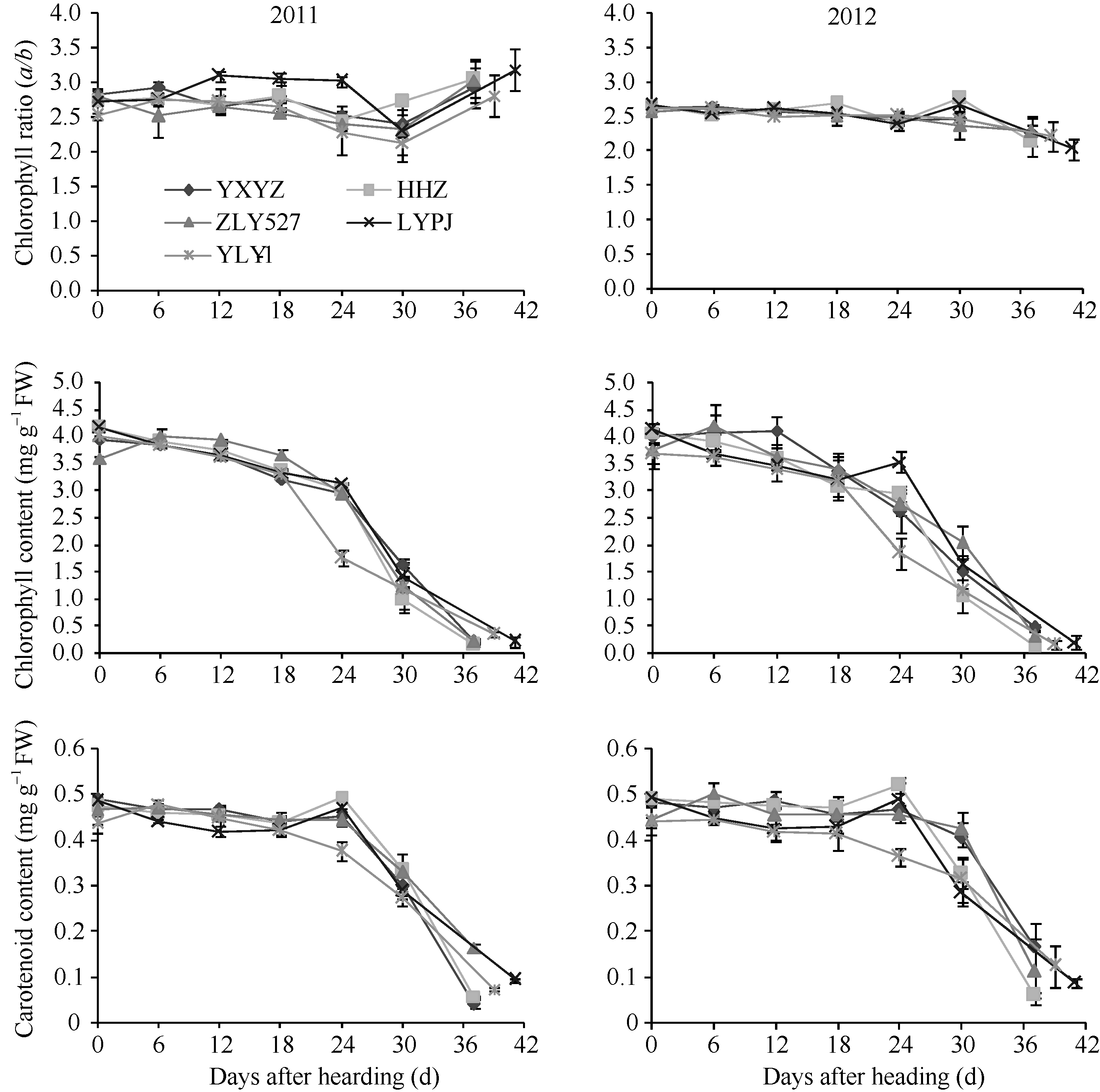
2.4.1 剑叶生理指标变化 水稻叶片叶绿素含量、叶绿素a/b值、类胡萝卜素含量是反映叶片衰老程度的重要生理指标。从图5可以看出, 不同穗粒型超级稻品种抽穗后剑叶叶绿素含量变化有差异, 年度间基本一致。其中, 小穗大粒型品种准两优527抽穗后叶绿素含量缓慢上升, 6 d出现小高峰后缓慢下降, 24 d后开始快速下降; 中穗中粒型品种Y两优1号抽穗后叶绿素含量开始缓慢下降, 18 d后快速下降, 下降幅度大; 其他3个品种抽穗后叶绿素含量开始缓慢下降, 24 d后快速下降。除2011年大穗中粒型品种两优培九外, 其他品种抽穗后叶绿素a/b值呈下降趋势, 24~30 d后变化趋势年度间不同, 2011年止降反升, 2012年延续下降或出现小高峰后下降。不同穗粒型超级稻品种抽穗后类胡萝卜素含量呈下降趋势, 到24 d大粒品种准两优527、两优培九止降反升出现小高峰; 其他品种则延续下降, 下降幅度以Y两优1号最大。
 | 图2 不同穗粒型超级稻上部、中部、下部枝梗籽粒增长动态缩写同表2。Abbreviations are the same as those given in Table 2.Fig. 2 Grain growth dynamics of upper, middle and lower branches grains of supper rice with different panicle-grain types |
 | 图3 不同穗粒型超级稻整穗籽粒灌浆速率变化缩写同表2。Abbreviations are the same as those given in Table 2.Fig. 3 Grain filling rate of the whole panicle of supper rice varieties with different panicle-grain types |
| 表4 不同穗粒型超级稻品种的平均灌浆速率 Table 4 Average grain filling rate of supper rice varieties with different panicle-grain types |
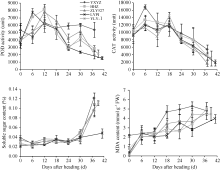
水稻抽穗后叶片过氧化氢酶(CAT)活性、过氧化物酶(POD)活性、丙二醛(MDA)含量与叶片的衰老程度有关。图6表明不同穗粒型超级稻抽穗后剑叶POD活性变化趋势品种间有差异。其中, 大穗小粒品种玉香油占抽穗后POD活性小幅下降后快速上升, 12 d出现第1个高峰, 大穗中粒品种两优培九、中穗小粒品种黄华占抽穗后POD活性快速上升, 分别在6 d、12 d出现第1个高峰, 小穗大粒品种准两优527、中穗中粒品种Y两优1号抽穗后POD活性缓慢上升, 12 d 出现第1个高峰。第1个高峰后, POD活性呈快速下降趋势, 24 d开始止降反升, 30 d出现第2个小高峰后开始下降。值得注意的是, 黄华占在第1个高峰后, POD活性仅有小幅下降, 直至成熟期仍有较高活性。图6还表明水稻剑叶中膜脂过氧化的中间产物MDA含量、可溶性糖含量与POD、CAT等抗氧化保护性酶的变化趋势相反。抽穗后剑叶MDA含量表现上升趋势, 品种间变化趋势一致, 但其含量差异明显, 以玉香油占含量较高, 两优培九含量较低。另外, 抽穗后30 d以内, 剑叶可溶性糖含量相对稳定或略有增加, 品种间差别不大; 抽穗30 d以后, 除两优培九外, 剑叶可溶性糖含量快速增加。可见, 抽穗后6 d剑叶开始衰老, 反映剑叶衰老的生理指标存在年度间、品种间差异, 且其变化趋势基本一致。
2.4.2 根系生理指标变化 从图7可知, 不同穗粒型超级稻根系活力(根系α -萘胺氧化力)抽穗后表现为下降趋势, 两年变化趋势一致, 但品种间差异明显。其中, 大穗中粒型品种两优培九、中穗小粒型品种黄华占先降后升; 中穗中粒型品种Y两优1号、小穗大粒型品种准两优527根系活力抽穗后开始上升, 抽穗12~18 d出现第1个峰值后缓慢下降, 18~24 d出现第2个峰值后快速下降; 大穗小粒品种玉香油占在抽穗后根系活力表现为先快后慢的下降趋势, 18~24 d止降反升, 出现一个小高峰值后持续下降。可见, 超级稻抽穗后根系开始衰老的时间早于剑叶。
 | 图4 不同穗粒型超级稻上部、中部、下部枝梗籽粒灌浆速率缩写同表2。Abbreviations are the same as those given in Table 2.Fig. 4 Grain filling rate of upper, middle and lower branches of supper rice with different panicle-grain types |
2.4.3 叶片、根系生理指标变化与籽粒增重的关系
从表5可以看出, 除了玉香油占、准两优527的POD活性及黄华占的叶绿素a/b值外, 剑叶CAT、POD等保护酶活性, 叶绿素、类胡萝卜素含量、叶绿素a/b与籽粒增重之间存在显著或极显著的负相关性, 表明水稻抽穗后叶片衰老不利于籽粒灌浆结
实, 即抽穗后剑叶保护酶活性、光合色素含量下降说明叶片开始衰老。可溶性糖、MDA含量与籽粒增重之间存在显著或极显著的正相关性, 但这仅仅是一种统计假象, 因为可溶性糖含量增加可能是碳水化合物的转运遇到阻碍, MDA含量增加恰恰是剑叶衰老的象征。
 | 图5 不同穗粒型超级稻抽穗后剑叶叶绿素含量、叶绿素a/b值及类胡萝卜素含量变化缩写同表2。Abbreviations are the same as those given in Table 2.Fig. 5 Variation of carotenoid, chlorophyll content and chlorophyll a/b ratio of the flag leaf after heading in super rice with different panicle-grain types |
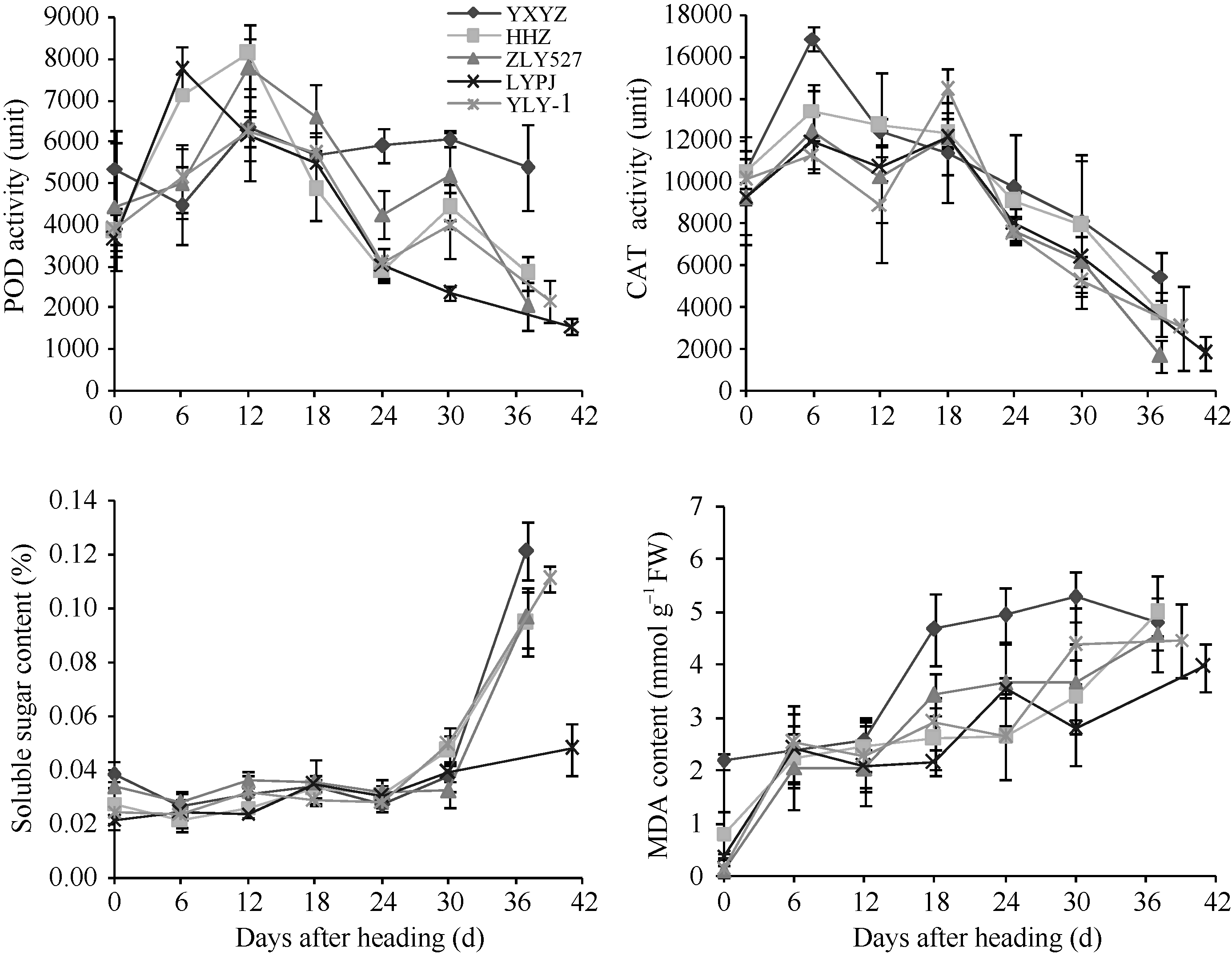 | 图6 不同穗粒型超级稻抽穗后剑叶POD、CAT酶活性和可溶性糖含量及MDA含量变化(2012年)缩写同表2。Abbreviations are the same as those given in Table 2.Fig. 6 Variation of POD, CAT activities, soluble sugar content and net photosynthetic rate of the flag leaf after heading in supper rice with different panicle-grain types in 2012 |
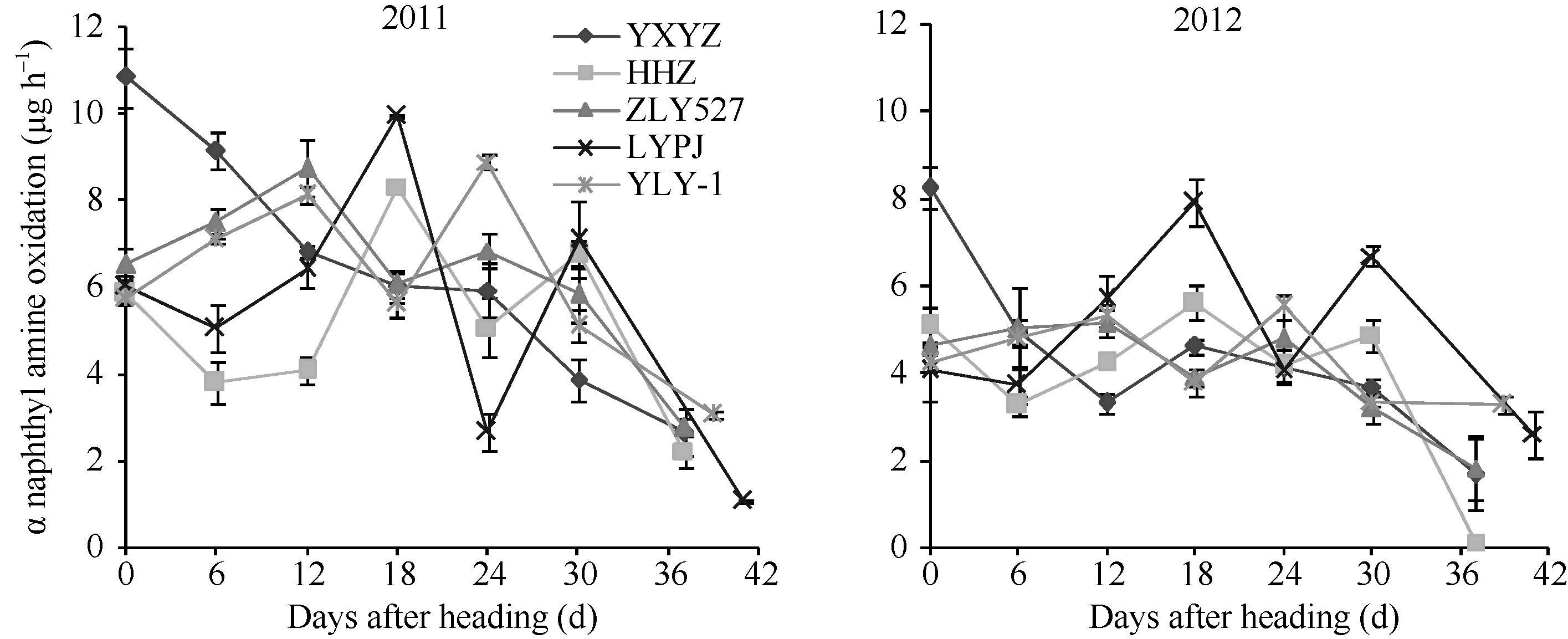 | 图7 不同穗粒型超级稻抽穗后根系活力变化缩写同表2。Abbreviations are the same as those given in Table 2.Fig. 7 Variation of root activity after heading in supper rice with different panicle-grain types |
| 表5 不同穗型超级稻抽穗后剑叶生理指标及根系活力与籽粒增重间的相关系数 Table 5 Correlation coefficient between the grain weight increasing and the flag leaf physiology index and root activity after heading in supper rice with different panicle and grain types |
水稻高产栽培的重要举措是在增加单位面积颖花数的基础上提高结实率和千粒重, 而结实率和粒重与籽粒增长或灌浆速率关系密切。水稻籽粒的增长动态表现为非线性过程, 可分为渐增期、快增期和缓增期[1, 2, 3, 5, 30, 31], 研究发现不同类型水稻籽粒灌浆强度在这3个增长期均存在差异[32, 33], 而增强籽粒的灌浆强度, 尤其是弱势粒的灌浆强度, 对于提高千粒重有重要作用[9, 24, 34]。本研究证明不同穗粒型超级稻籽粒增长的品种间差异明显, 无论在渐增期, 还是快增期, 以大穗小粒品种(玉香油占)籽粒增长最慢, 小穗大粒型品种(准两优527)增长最快, 但进入缓增期后, 籽粒增长的品种间差异不大。研究还发现稻穗上、中、下不同部位枝梗籽粒增长动态差异明显, 即上部枝梗籽粒≥ 中部枝梗籽粒≥ 下部枝梗籽粒, 这一变化趋势在品种间基本一致。
水稻籽粒充实度与其穗上着生的部位有关。一般稻穗中上部早开花的强势粒, 灌浆快、充实好、粒重高; 稻穗下部迟开花的弱势粒, 灌浆慢、充实差、粒重低[1, 3, 8, 12, 35], 其强、弱势籽粒灌浆速率的差异对于大穗型品种[9, 35, 36]或密穗型品种[8]尤为突出。本研究证明超级稻籽粒灌浆强度与枝梗着生的部位和品种的穗粒类型有关, 即上部枝梗籽粒≥ 中部枝梗籽粒≥ 下部枝梗籽粒, 大粒型品种≥ 中粒型品种≥ 小粒型品种。小穗大粒型品种准两优527灌浆起步快、强度大, 在开花后18 d达到灌浆高峰且持续时间长; 大穗小粒品种玉香油占灌浆起步慢, 花后12~18 d达到灌浆高峰, 峰值持续时间短; 中穗小粒型品种黄华占灌浆起步快, 花后12 d达到灌浆高峰, 峰值持续时间短; 大穗中粒型品种两优培九和中穗中粒型品种Y两优1号灌浆速率快, 花后12~18 d出现第1个灌浆高峰, 峰值持续时间长或出现第2个灌浆高峰, 表现出“ 两段式灌浆现象” [6]。本研究还发现, 稻穗上部一次枝梗籽粒表现出一个灌浆高峰, 为一段式灌浆, 高峰出现在抽穗后6~18 d, 其灌浆强度大; 稻穗中、下部枝梗籽粒灌浆速率变化因品种的穗粒型不同而异, 即大粒型品种为一段式灌浆, 中、小粒型品种为两段式灌浆。
有报道指出单位面积产量与籽粒灌浆持续时间呈正相关, 与籽粒灌浆速率呈负相关或相关不显著[37], 因此延长籽粒灌浆持续时间, 有利于产量形成。也有报道指出水稻单位面积产量受灌浆能力的约束, 籽粒灌浆能力可通过激发源活性和促进非结构性碳水化合物转移得到改善[38]。本研究证明中粒型品种因籽粒灌浆强度较大, 持续时间较长, 有利于高产; 大粒型品种虽然灌浆强度大, 但籽粒灌浆高峰持续时间短, 不利于高产; 小粒品种虽然灌浆高峰持续时间长, 但灌浆强度小, 同样不利于高产。可见, 水稻灌浆强度及其持续时间可作为水稻新品种选育和高产栽培的技术指标。
本研究证明籽粒灌浆启动后, 剑叶逐渐衰老, 叶绿素含量、叶绿素a/b值、类胡萝卜素含量降低, 剑叶可溶性糖含量、MDA含量逐渐上升, 这与前人研究结果基本一致[16, 17, 18]。由于根系衰老早于叶片, 维持水稻后期较高的根系活力, 将有利于缓和叶片衰老和促进籽粒灌浆结实[19, 20, 21, 22, 23]。本研究发现超级杂交稻剑叶抽穗后POD活性、CAT活性及根系活力先后出现两次升降过程, 剑叶叶绿素含量、叶绿素a/b值、类胡萝卜素含量、CAT活性、POD活性、根系α -萘胺氧化力与光合速率呈极显著正相关, 其相关系数分别为0.638、0776、0.883、0.610、0.832、0.495; 可溶性糖含量、MDA含量与光合速率呈极显著负相关, 其相关系数分别为-0.716、-0560。本研究还发现大穗小粒品种玉香油占抽穗后剑叶产生的活性氧和自由基增加, 诱导产生的POD和CAT活性高, 但不能完全消除活性氧和自由基的氧化作用, 其MDA含量依然较高, 可能就是其产量低的原因之一。相反, 大穗中粒型品种两优培九由于抽穗后产生的活性氧和自由基较少, 诱导产生的POD和CAT活性较低, 其MDA含量较低, 可能是其产量高的原因之一。
不同穗粒型超级稻籽粒增长符合非线性模式, 即经过渐进增长、快速增长、缓慢增长3个过程, 但籽粒灌浆强度及灌浆持续时间的品种间差异显著, 表现出一段式灌浆, 或两段式灌浆过程。籽粒灌浆强度既与其枝梗着生的部位有关, 即上部枝梗籽粒≥ 中部枝梗籽粒≥ 下部枝梗籽粒; 又与品种的穗粒类型有关, 即大粒型品种≥ 中粒型品种≥ 小粒型品种。籽粒灌浆启动后, 叶绿素含量、叶绿素a/b值、类胡萝卜素含量逐渐降低, 剑叶可溶性糖含量、MDA含量逐渐上升, POD活性、CAT活性及根系活力先后出现2次升降过程。研究结果为超级稻高产栽培与新品种选育提供了参考。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|