



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
花后低温对水稻籽粒灌浆与内源激素含量的影响
[曾研华1, 2, ** , 张玉屏2, ** , 潘晓华1 , 朱德峰2, *  , 向镜
, 向镜2 , 陈惠哲2 , 张义凯2 ]
, 向镜|
|


第一作者联系方式: 曾研华, E-mail: zyh74049501@163.com; 张玉屏, E-mail: cnrrizyp@163.com
以籼粳杂交稻甬优538、甬优17和籼稻中浙优1号、粳稻浙粳88为材料, 设置灌浆期不同时段低温处理, 分析籽粒灌浆过程中内源激素含量的动态变化及其差异。结果表明, 花后不同时段低温对籽粒灌浆的影响表现为前期>中期>后期, 而中、后期与对照差异不大, 且低温对甬优17、中浙优1号籽粒灌浆的影响大于对甬优538和浙粳88。花后前期低温显著降低甬优17籽粒灌浆, 延长灌浆时间, 而甬优538与对照差异不明显。花后不同温度处理下供试品种内源激素含量变化存在一定差异, 其变化与品种灌浆动态保持一致。花后前、中期低温明显降低甬优17籽粒中IAA、ZR含量, 显著增加GA3、ABA含量; 而对甬优538显著下降GA3含量和略增加IAA、ZR含量, 这可能是籽粒正常灌浆的主要原因。生产中可通过适宜的激素调控措施来提高水稻籽粒低温灌浆。
Low temperature (LT) after flowering impacts greatly on grain filling, decreasing rice quality through extended filling duration, shortened filling rate, and decreased physiological activity. Indica-japonica hybrid rice has been applied widely in production practice, due to high yield potential. However, there has been little study on the effect of LT on endogenous hormones in indica-japonica hybrid cultivars at filling stage. In the study, four varieties, i.e. two indica-japonica hybrid rice (Yongyou 538, YY538; Yongyou 17, YY17), one indica rice (Zhongzheyou 1, ZZY1) and one japonica rice (Zhejing 88, ZJ88), were treated with those low temperature treatments at different grain filling phases, to compare endogenous hormone levels in treatments, aiming to reveal the influence of low temperature on grain filling and the response of endogenous hormone in grains to low temperature. The effect of LT on grain filling after different flowering stage appeared significantly at early grain filling stage, while a little difference was observed at later filling stage, compared with CK. Furthermore, the influence on grain filling was found higher in Yongyou 17 and Zhongzheyou 1 than in Yongyou 538 and Zhejing 88. Compared with the contract, LT after flowering at early stage significantly decreased grain filling and prolonged grain filling duration in Yongyou 17, but not in Yongyou 538. Based on the effect of LT on grain filling process and its relation with endogenous hormones contents, it might be deduced that the normal grain filling should be ascribed to the significantly reduced GA3 content and the slightly increased IAA, ZR contents in developing grains of Yongyou 538 at early and middle grain-filling stage. On the contrary, the significantly reduced IAA, ZR contents and the significantly increased GA3, ABA contents in Yongyou17 could restrain grain development. It should enhance grain filling of rice encounted LT by selected measures adjusting hormone in practice management.
随着全球气候变暖, 极端性的低温冷害事件也频频发生[1, 2], 严重影响水稻生产, 而水稻又是喜温性作物, 对温度非常敏感。在温度胁迫条件下, 水稻产量变化是各种器官生长发育过程和生理生化变化的最终体现。关于温度对水稻籽粒发育的影响, 前人从籽粒灌浆特性[3]、淀粉合成代谢[4, 5]、氮代谢[6]、籽粒内源激素含量变化[3]等方面进行了较为深入的报道。程方民等[5]研究认为高温处理有利于增加灌浆初期籽粒中的蔗糖含量、淀粉含量及蔗糖合成酶、焦磷酸化酶、可溶性淀粉合成酶、淀粉分支酶和淀粉去分支酶, 而灌浆中后期有关酶的活性变化与蔗糖含量、淀粉含量间的关系相对较为复杂。但多以高温研究为主[7, 8, 9, 10, 11]。高温处理显著降低水稻灌浆初期籽粒IAA、ZR、GA3的含量, 对灌浆中、后期的籽粒内源激素含量影响较小[3], 在玉米不同穗位中趋势也一致[12]。而关于低温处理的影响, 袁莉民等[13]研究指出灌浆期不同时段低温处理, 易导致籽粒外形异常, 垩白面积提高, 胚乳结构明显变化。付景等[14]研究认为超级稻品种籽粒结实对低温反应比常规高产品种更敏感。且灌浆期低温显著降低结实率和穗粒重[15]。杨杰等[16]发现籼粳杂种低温花粉不育的QTL遗传特性。但结实期水稻籽粒灌浆与其内源激素含量变化的关系尚不清楚[13]。籼粳杂交稻, 尤其是甬优系列籼粳杂交稻[17, 18, 19], 由于具有强大的杂种优势, 产量潜力巨大, 目前在生产上大面积推广, 产生了巨大的产量效益和经济效益。但其穗大粒多和生育后期过长的灌浆结实期增加了花后遭遇低温风险的机率, 此外, 结实期温度是不断变化的。因此, 在花后各时段设置不同低温, 探讨籽粒发育过程中内源激素含量的变化规律, 阐明籽粒发育的生理机理, 具有重要的意义。本文研究灌浆结实期不同时段低温对籼粳杂交稻籽粒内源激素含量的影响, 揭示低温对水稻籽粒内源激素含量的调节作用及其与籽粒灌浆的关系, 为籼粳杂交稻抗逆栽培提供理论依据。
于2013— 2014年在中国水稻研究所富阳试验基地人工气候箱(PGV-36型CONVIRON)内进行盆栽试验。供试品种为甬优17和甬优538 (籼粳杂交稻), 对照品种为中浙优1号(杂交籼稻)、浙粳88 (常规粳稻)。
参考曾研华等[20]的种植方法和温度日变化设置。人工气候箱具有自动控温控湿的功能, 气候箱透光面所用材质为超透明的超白钢化玻璃, 具有低自爆率、颜色一致、可见光透过率高、通透性好等特点, 其光照以外界自然光为主, 光谱色损失少, 基本保持箱内光照与外界自然光照一致, 同时人工气候箱底部装有鼓风装置, 能保持二氧化碳浓度与室外基本一致。箱内环境条件能达到试验处理要求。
根据长江中下游近60年来9月至10月份的气候温度变化, 并结合富阳地区近5年来9月至10月份室外同期温度的变化规律, 以及花后面临外界环境温度的动态变化, 设置花后3个不同低温度处理时段。第一时段为花后0~15 d, 温度为20℃ (Tmin/Tmax: 17℃/23℃), 记作S1, 称作灌浆前期; 第二时段为花后15~30 d, 温度为17℃ (14℃/20℃), 记作S2, 称作灌浆中期; 第三时段为花后30 d至成熟期, 温度为14℃ (11℃/17℃), 记作S3, 称作灌浆后期。气候箱内对照处理(CK)温度为25.0℃ (22℃/ 28℃)。各时段低温处理结束后, 全部盆栽植株在气候箱内恢复25.0℃, 进行正常生长, 与对照保持一致的生长环境, 直至成熟。为保持各品种花期基本一致而错期播种, 中浙优1号、甬优17和浙粳88均为6月14日播种, 7月4日移栽, 甬优538为6月21日播种, 7月11日移栽, 保持各品种9月4日左右开花。每品种各低温处理15盆, 对照26盆, 以保证足够的稻穗取样。穗顶抽出剑叶鞘5 cm左右时选择大小基本一致的单穗(包括主茎穗和部分分蘖穗)挂牌标记, 每处理标记200~300穗, 供取样分析。
1.3.1 籽粒灌浆速率测定 于开花后第0、第3、第6、第9、第12、第15、第18、第21、第27、第33、第39、第45、第51、第57天分别取稻穗8~10个, 摘下所有籽粒并剔除病粒空粒, 置70℃烘箱中烘干至恒重, 然后称重。用Richards方程按照朱庆森等[21]和顾世梁等[22]方法对籽粒灌浆过程进行拟合计算灌浆速率。
1.3.2 籽粒内源激素的提取与含量测定 S1 (0~15 d)时段于低温处理第7、第14天取样, 即花后第7、第14天; S2 (15~30 d)时段于低温处理第6、第13天取样, 即花后第21、第28天; S3 (30 d至成熟)时段于低温处理第5天取样, 即花后第35天。
取各时段处理挂牌稻穗中上部籽粒30~40粒, 用80%预冷甲醇置弱光下冰浴研磨至匀浆, 于4℃过夜, 10 000× g冷冻离心15 min取上清液过C18柱; 真空干燥后, 以样品稀释液(含0.1% Tween-20和0.1%白明胶的磷酸盐缓冲液, pH 7.5)溶解即得样品激素提取液。采用酶联免疫法测定IAA、GA3、ABA和ZR含量, 参照Yang等[23]的方法。酶联免疫试剂盒购自中国农业大学, 每样品重复测定3次, 取平均值。IAA、ZR、ABA和GA3的回收率分别为95.1%、86.1%、92.2%和89.1%。
用Microsoft Excel 2003处理数据和SPSS16.0软件作统计分析, 采用Duncan’ s法检验差异显著性。因2013年为预备试验, 本文主要报告2014年数据。
由图1可见, 与对照相比, 花后各时段低温对籽粒灌浆的影响以灌浆前期高于中、后期, 而灌浆中、后期与对照差异不大, 且存在品种间差异。灌浆前期低温下甬优17籽粒结实初期灌浆偏慢, 在0~9 d趋于停滞, 之后其籽粒重量一直低于对照处理, 而花后9~27 d内差异明显; 灌浆中期低温下籽粒重量在花后15~30 d比对照略有下降, 而灌浆后期低温下籽粒灌浆与对照差异不大。对照籼稻中浙优1号籽粒增重与之较为相似。此外, 甬优538和对照粳稻浙粳88的各时段低温处理籽粒重量趋势较为一致。总体来看, 花后灌浆前期低温明显影响甬优17和中浙优1号的籽粒灌浆, 延长灌浆持续时间, 而对甬优538和浙粳88的影响不明显。
表1中, 籽粒最终重量(A)与其灌浆进程相对应, A值总体表现为S1< S2< S3, 以中浙优1号和甬优17灌浆期低温处理下降显著, 而甬优538与浙粳88差异不明显。同时, 各拟合方程的决定系数均在0.99以上, 拟合度高, 说明用Richards方程描述不同温度处理下水稻籽粒灌浆过程是可行的。
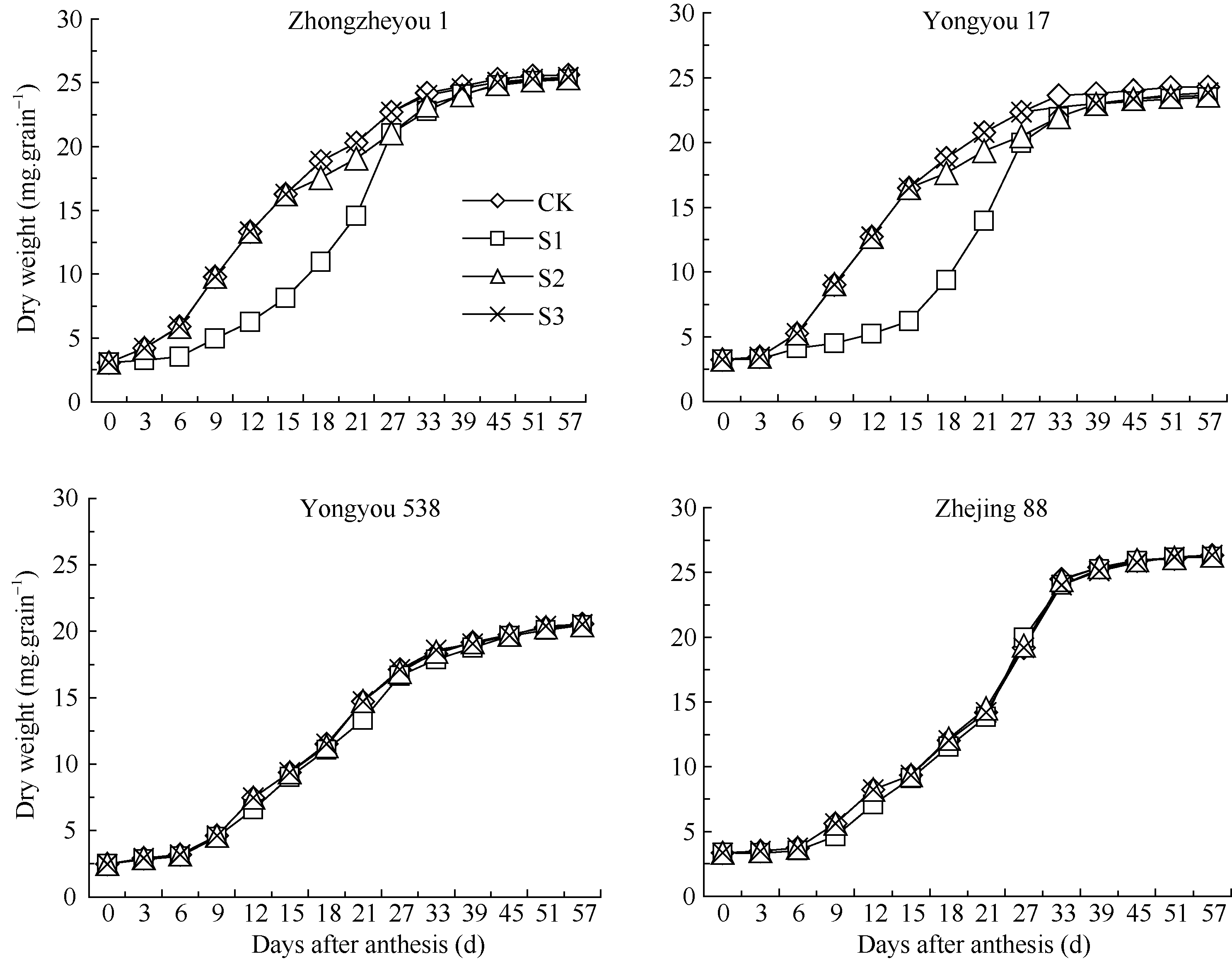 | 图1 灌浆期温度处理下不同品种籽粒重量动态变化S1、S2、S3为花后灌浆前、中、后期时段的低温处理。Fig. 1 Dynamics of grains weight of varieties at grain-filling stageS1, S2, S3 indicate low temperature from 0 to 15 days, from 15 to 30 days, and from 30 days after flowering to maturity, respectively. |
| 表1 灌浆期温度处理下籽粒灌浆过程的Richards方程参数 Table 1 Parameters of Richards equation for grain filling |
由图2可见, 花后各时段低温下4个品种籽粒灌浆速率变化规律与籽粒增重相似。灌浆前期低温下甬优17籽粒结实初期灌浆速率明显低于对照, 达到最大灌浆速率的天数(27 d)明显迟于对照(12 d), 平均灌浆速率比对照低15.47%; 而灌浆中期低温籽粒灌浆峰值较对照略有下降。对照籼稻中浙优1号籽粒灌浆速率与甬优17较为相似, 前期低温籽粒平均灌浆速率比对照低21.06%。总体来看, 花后灌浆前期低温显著降低甬优17和中浙优1号籽粒灌浆速率, 减缓灌浆进程, 而对甬优538和浙粳88则不明显。
2.3.1 生长素(IAA)含量 由图3可知, 各品种籽粒IAA含量变化均呈先升后降趋势, 但品种间存在一定差异。甬优17和中浙优1号籽粒IAA含量各处理均低于对照, 且灌浆前期低温处理差异更明显, 说明籽粒胚乳细胞的生长受到严重影响; 甬优538低温处理籽粒的IAA含量在花后21 d显著高于对照, 其余时期与对照差异不显著, 而对照粳稻低温处理总体略高于对照。
2.3.2 玉米素(ZR)含量 由图4可知, 供试品种籽粒灌浆过程中ZR含量动态总体均呈下降趋势, 灌浆后期低温与对照持平。甬优17籽粒的ZR含量前、中期低温均显著低于对照, 说明不利于胚乳细胞增殖与分化; 但甬优538的ZR含量在花后7 d显著高于对照, 而后与对照相差不大。对照籼稻籽粒ZR含量变化与甬优17相似, 粳稻则与甬优538相似。
2.3.3 赤霉素(GA3)含量 与ZR变化相似, 籽粒中GA3在灌浆初期较高, 而后随着灌浆进程, 总体呈下降趋势(图5)。甬优17籽粒GA3含量各阶段低温处理均高于对照, 且花后14~28 d差异显著, 对照籼稻变化趋势与之相似; 相比甬优17, 甬优538和粳稻浙粳88的GA3含量前、中期低温处理低于对照, 且花后7~14 d差异达显著水平, 为保证籽粒正常灌浆奠定了基础。表明各品种籽粒灌浆进程的差异与花后前、中期低温处理下GA3含量变化有关。
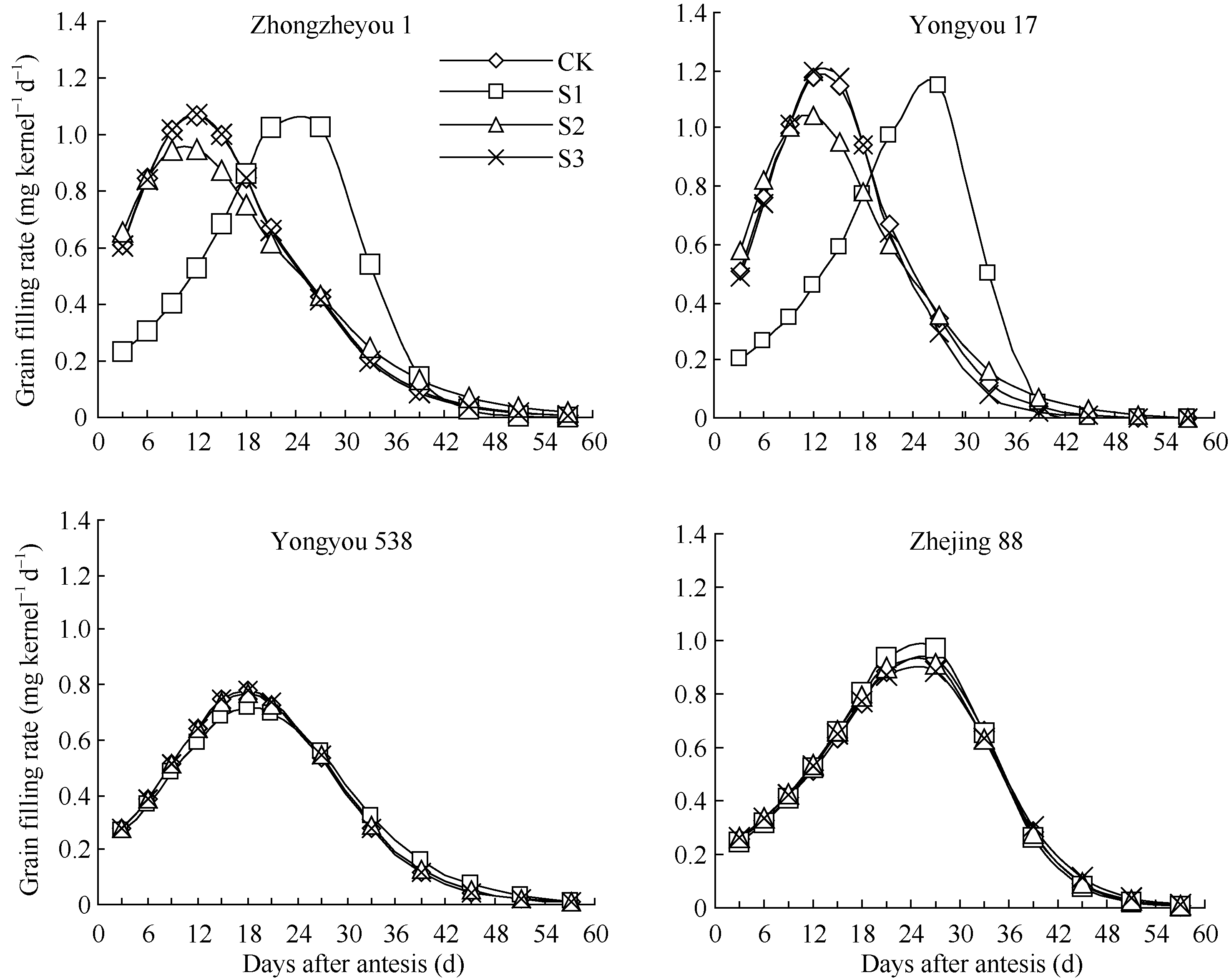 | 图2 灌浆期温度处理下不同品种籽粒灌浆速率变化S1、S2、S3为花后灌浆前、中、后期时段的低温处理。Fig. 2 Dynamics of grain filling rate of varieties in low temperature treatmentsS1, S2, S3 indicate low temperature from 0 to 15 days, from 15 to 30 days, and from 30 days after flowering to maturity, respectively. |
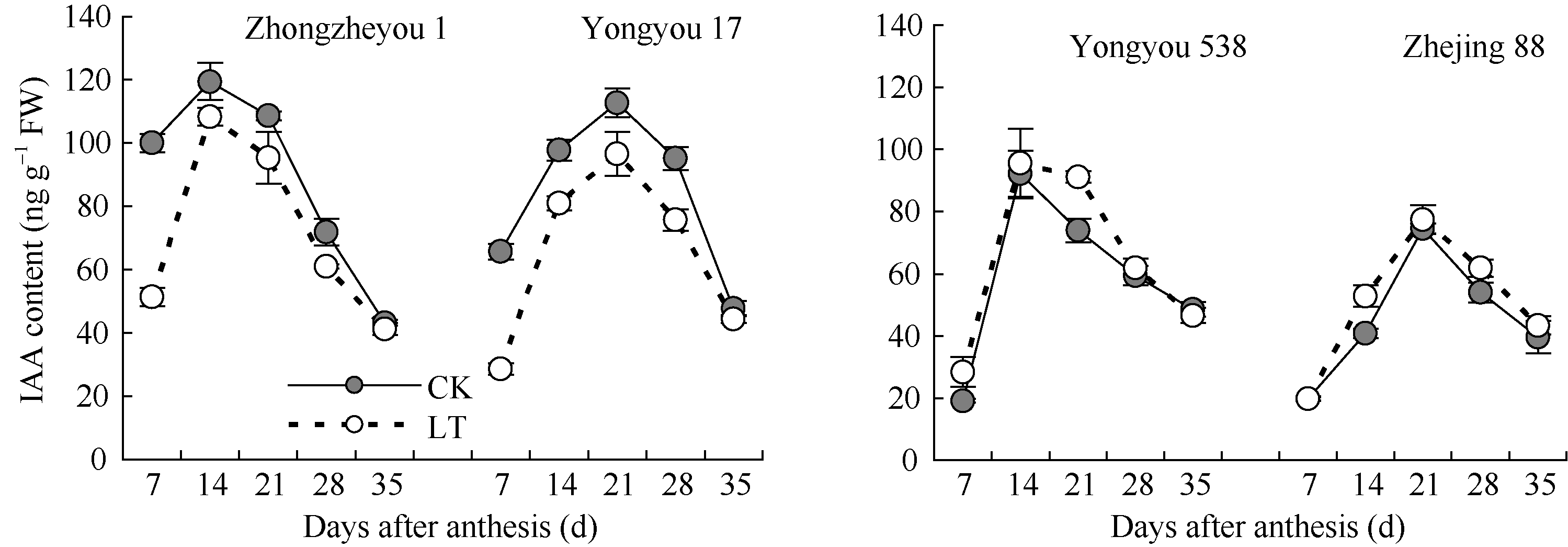 | 图3 灌浆期低温对籽粒IAA含量的影响LT: 低温处理, 包括花后灌浆前期第7、第14天, 灌浆中期第21、第28天, 灌浆后期第35天。Fig. 3 Effect of low temperature during grain-filling on IAA content in rice grains“ LT” indicates the seventh and fourteenth days, the twenty-first and twenty-eighth days, and the thirty-fifth day after flowering, respectively in low temperature treatment. |
2.3.4 脱落酸(ABA)含量 由图6可知, 供试品种籽粒ABA含量随籽粒灌浆进程呈下降的变化趋势。甬优17和中浙优1号籽粒中ABA含量低温处理均低于对照, 且甬优17前、中期差异显著; 而甬优538和浙粳88籽粒ABA含量与对照差异较小。说明在籽粒灌浆过程中, 前、中期低温对甬优17和中浙优1号籽粒ABA的形成和积累有显著的促进作用, 从而阻碍了灌浆进程。
相关性分析(表2)表明, 籽粒灌浆与ZR、GA3和ABA含量均呈负相关, 达到极显著水平, 与IAA呈正相关但不显著, 说明花后低温处理下, ZR、GA3和ABA含量变化协同影响籽粒灌浆进程。各激素间, ABA与ZR、GA3含量呈显著或极显著正相关, ZR与GA3含量呈极显著正相关, 而IAA与ZR、GA3、ABA含量相关但不显著, 表明各激素存在密切而复杂的关系。
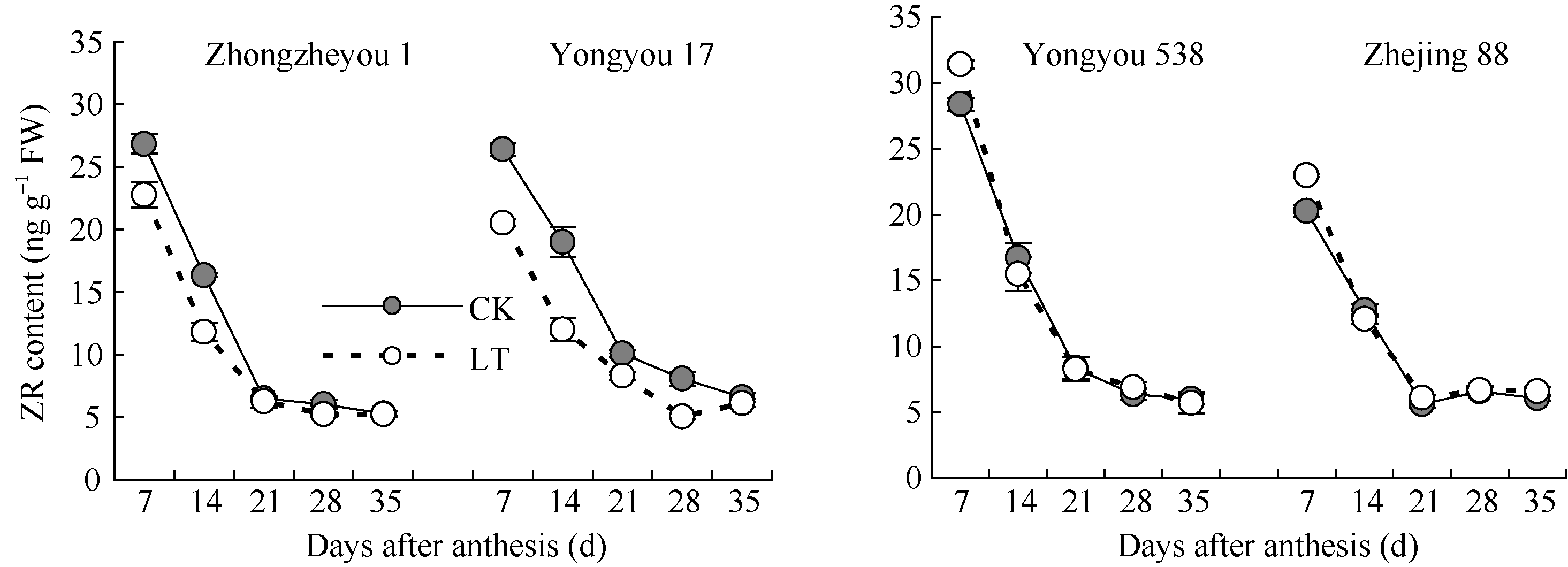 | 图4 灌浆期低温对籽粒ZR含量的影响LT: 低温处理, 包括花后灌浆前期第7、14天, 灌浆中期第21、28天, 灌浆后期第35天。Fig. 4 Effect of low temperature during grain-filling on ZR content in rice grains“ LT” indicates the seventh and fourteenth days, the twenty-first and twenty-eighth days, and the thirty-fifth day after flowering, respectively in low temperature treatment. |
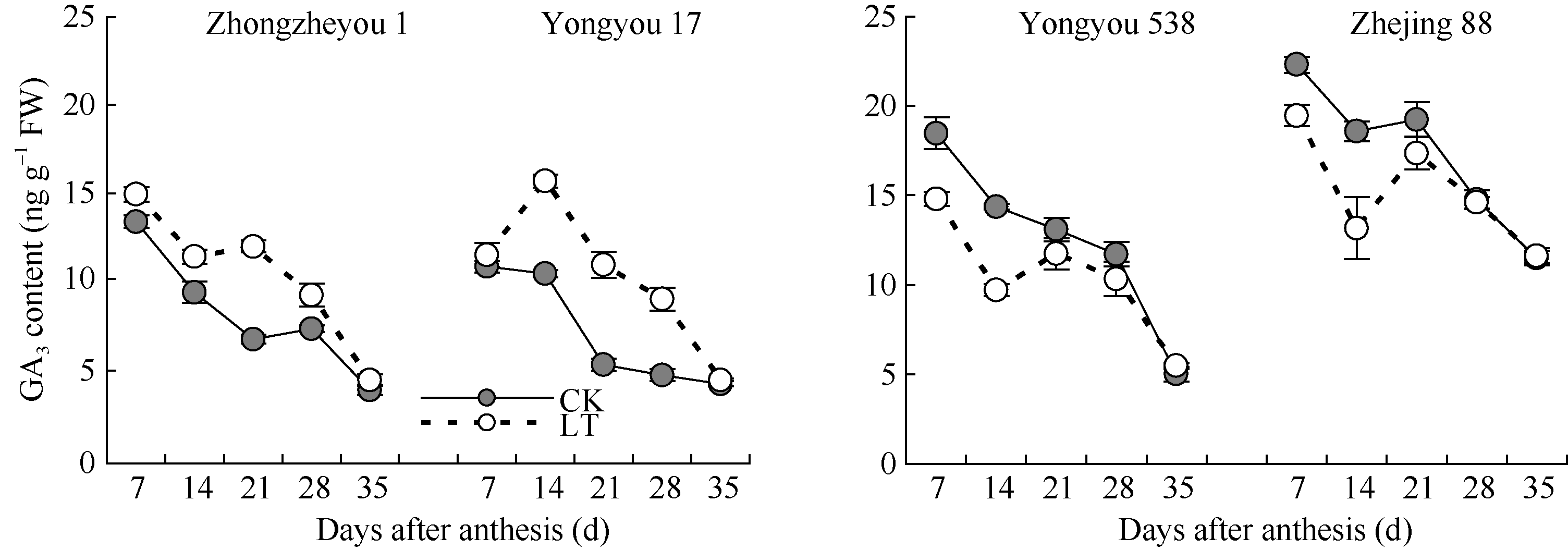 | 图5 灌浆期低温对籽粒GA3含量的影响LT: 低温处理, 包括花后灌浆前期第7、第14天, 灌浆中期第21、第28天, 灌浆后期第35天。Fig. 5 Effect of low temperature during grain-filling on GA3 content in rice grains“ LT” indicates the seventh and fourteenth days, the twenty-first and twenty-eighth days, and the thirty-fifth day after flowering, respectively in low temperature treatment. |
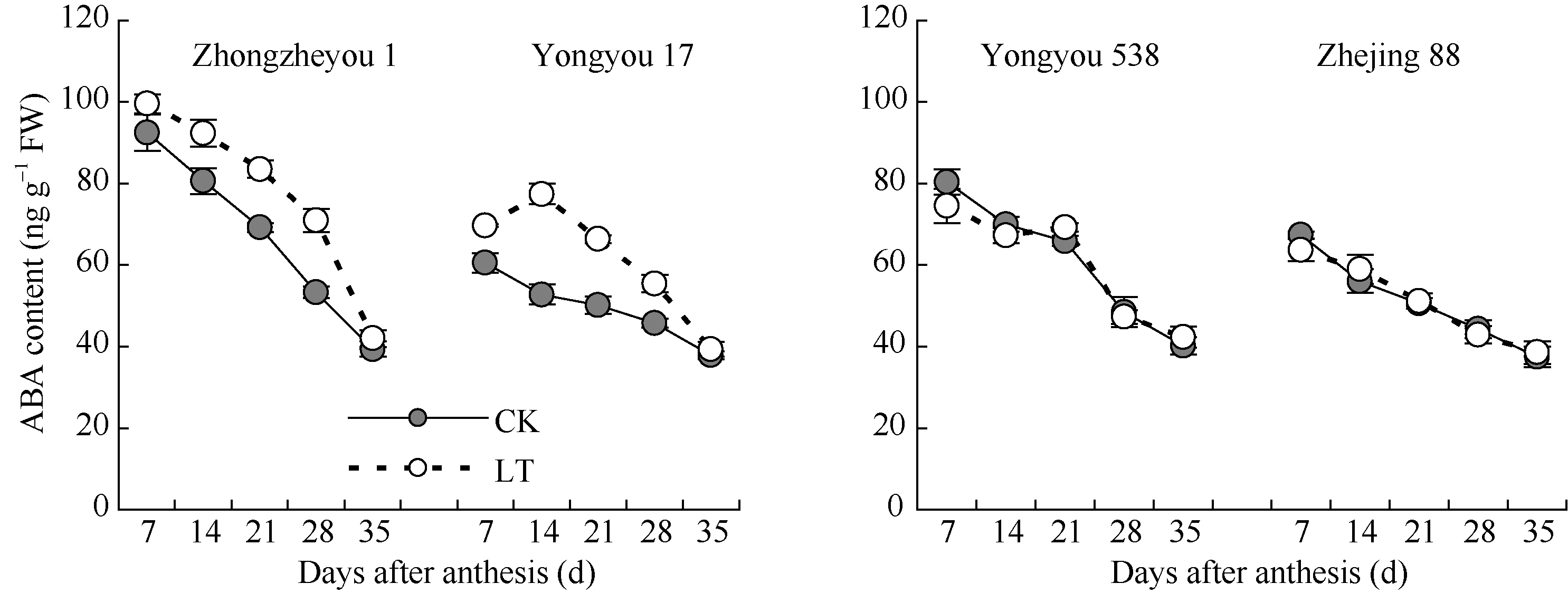 | 图6 灌浆期低温对籽粒ABA含量的影响LT: 低温处理, 包括花后灌浆前期第7、14天, 灌浆中期第21、28天, 灌浆后期第35天。Fig. 6 Effect of low temperature during grain-filling on ABA content in rice grains“ LT” indicates the seventh and fourteenth days, the twenty-first and twenty-eighth days, and the thirty-fifth day after flowering, respectively in low temperature treatment. |
灌浆结实期是籽粒发育和粒重增长的重要时期。研究认为低温通过降低最大灌浆速率而延迟整个灌浆进程[24, 25]。齐穗后20 d是粒重增长及温度敏感的主要时段, 此时段正是水稻受精后胚乳生长的关键时期[26]。从本试验结果来看, 花后低温影响籽粒灌浆, 低温的效应以灌浆前期大于中、后期, 这与前人研究结论相似[14, 25]; 供试品种籼粳杂交稻和籼、粳稻的籽粒灌浆对低温的响应存在差异, 以低温对甬优17、中浙优1号的影响高于甬优538和浙粳88, 验证了前期工作中花期耐冷性鉴定的结论[20]。甬优17比甬优538籽粒灌浆启动早, 灌浆速率快, 达到最大灌浆速率时间早, 但同时受低温影响大, 可能也是对低温敏感的另一重要因素。此外, 花后前期低温处理对甬优17籽粒灌浆影响最大, 显著降低灌浆速率, 延长灌浆时间, 中期影响相对较小, 而后期几乎没有影响; 甬优538籽粒灌浆各时段变化不显著。
| 表2 花后不同温度处理下水稻籽粒灌浆与籽粒中内源激素含量变化的相关分析 Table 2 Correlation coefficients between grain filling and changes of endogenous hormone contents in different temperature treatments |
植物内源激素对水稻籽粒发育过程起重要的调节作用, 胁迫条件影响作物籽粒中植物激素的含量[27]。一般而言植物内源激素IAA、ZR为生长促进型激素, 而ABA、GA3为生长抑制型激素。前人从高温角度探讨了籽粒内源激素含量的变化, 研究认为花后前期高温降低籽粒IAA、ZR等激素含量[3, 12]。本研究结果显示, 花后不同低温处理下, 供试品种籽粒中内源激素含量变化存在一定差异, 其变化与品种灌浆动态保持一致。植物在生长发育阶段中引起生理变化的某一个反应往往是激素间协同作用的结果, 而不是某一种激素单独作用的结果[28]。研究表明ZR能促进籽粒中胚乳细胞分裂[12]。同时, IAA含量与籽粒淀粉含量具有较好的一致性, 可促进籽粒淀粉含量的积累[29]。本试验甬优17和对照品种中浙优1号籽粒灌浆前、中期低温处理下籽粒中IAA、ZR含量显著降低。由此说明灌浆前、中期低温通过降低籽粒ZR含量而抑制胚乳细胞分裂和分化来降低籽粒灌浆; 同时通过降低IAA含量, 减少胚乳细胞数量的增加和体积的增大, 从而降低谷粒同化产物的积累, 并影响与谷粒积累同化产物有关的酶(蛋白质), 最终抑制籽粒淀粉含量的积累, 导致籽粒灌浆不良。这与高温处理下研究的结果较为相符[3, 12]。此外, 胚乳细胞活跃增殖和籽粒活跃灌浆期与籽粒中的GA3含量呈显著负相关[30]。灌浆前、中期低温处理通过增加甬优17和中浙优1号籽粒的GA3含量, 这表明GA3含量的增加进一步影响了籽粒胚乳细胞的增殖与灌浆进程, 降低籽粒灌浆速率, 从而延长了籽粒灌浆持续期。有研究表明, 水稻灌浆初期, ABA主要通过提高籽粒淀粉合成酶、酸性转化酶及ATP酶的活性从而提高籽粒对蔗糖的卸载和转化能力[31, 32], 通过提高籽粒ADPG焦磷酸化酶和淀粉分支酶(SBE)活性促进淀粉合成[31]。但甬优17和中浙优1号籽粒有较高含量的ABA, 并未增加胚乳细胞的物质吸收, 反而降低籽粒充实, 说明低温处理下ABA在籽粒积累同化产物方面可能起着抑制作用, 导致籽粒中ABA与GA3、IAA等激素的比值失衡, 影响胚乳细胞内淀粉的进一步合成, 从而阻碍灌浆进程。这与前人研究报道的结果不一致[3, 12, 33], 可能与试验处理或研究对象不同有关。同时, 也暗示低温处理下植物各内源激素间可能存在着拮抗作用, 尽管某些激素与籽粒灌浆存在一致的负相关性。对于甬优538和对照粳稻浙粳88而言, 灌浆前、中期低温处理下籽粒明显较低的GA3和稍高的IAA、ZR含量, 保证了籽粒胚乳细胞的分裂与增殖速率、淀粉的合成与积累、以及籽粒的灌浆进程, 可能也是其籽粒低温下能完成正常灌浆的主要原因。此外, 本研究基于内源激素含量的变化规律, 阐明了甬优538比甬优17号耐低温灌浆结实的机制, 可为生产中通过适宜的激素调控措施来提高籼粳杂交稻抗逆栽培提供理论依据。
本试验籼粳杂交稻灌浆期各时段的低温处理并没有选择同一个温度, 而是基于不同时段分别设置了不同的温度梯度, 比以往单纯设置单一温度的研究方法[13, 34], 具有一定的合理性和真实性。花后灌浆结实期所处的外界环境温度其实是一个动态变化的过程, 籽粒灌浆进程的不同, 对温度的响应也会不一致; 此外, 籼粳杂交稻花后具有较长的灌浆结实期, 长达60 d左右, 此时所处9月至10月份的外界环境温度多变, 产生的温度差异也很大, 导致籽粒灌浆遭遇温度的影响趋势不一。因此, 有必要根据水稻花后各时段籽粒的灌浆进程, 并结合其所处自然环境温度的变化, 来合理地降低设置的处理温度, 以更能反应外界自然环境温度对水稻籽粒灌浆的真实影响, 保证具有一定的可行性和准确性。
花后低温降低水稻籽粒灌浆, 以灌浆前期低温处理影响最大, 灌浆中、后期其次。甬优538籽粒灌浆结实比甬优17更耐低温, 花后前期低温显著降低甬优17籽粒灌浆, 延长灌浆时间, 而对甬优538的效应不显著。花后低温处理下, 各品种内源激素含量变化存在一定差异, 甬优17灌浆前、中期籽粒中IAA、ZR含量显著降低, 但GA3、ABA含量明显提高; 而甬优538籽粒中GA3含量显著降低, IAA、ZR含量较高, 这是保证籽粒能正常灌浆的主要原因。低温处理后籽粒中内源激素含量变化影响籽粒灌浆, 其变化与品种灌浆动态保持一致。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|