



{kind=link}
{kind=link}
{kind=link}
{kind=link}
小麦DELLA获得性突变体矮变1号增强了幼苗的抗盐能力
[王润青1, 2, *  , 樊晓聪
, 樊晓聪2, 3, * , 宋梅芳2, 4 , 肖阳5 , 郭林2 , 孟凡华2 , 杨青华3 , 吴大付1, * , 杨建平1, 2, * ]
, 樊晓聪, 宋梅芳, 杨建平]
|
|


第一作者联系方式: Tel: 010-82105851; 王润青, E-mail: wrq1990520@163.com; 樊晓聪, E-mail: xiaocong_fan@163.com
DELLA蛋白是GA信号途径中重要的负向调控因子, 能够响应各种环境信号, 在植物抵抗生物和非生物胁迫中起着重要的作用。功能获得性DELLA突变体矮变1号在矮化育种上已得到广泛应用, 但是其在耐盐方面的研究鲜有报道。本文采用含200 mmol L-1 NaCl的Hoagland水培溶液处理中国春、京411、矮秆早和矮变1号幼苗7 d, 测定盐胁迫下的总叶绿素含量、相对含水量、超氧化物歧化酶(SOD)活性和丙二醛(MDA)含量, 并借助免疫印迹检测了4个小麦品种中DELLA蛋白的积累。盐胁迫处理后, 矮变1号的叶片没有出现明显的萎蔫和失绿, 而其他3个品种均表现出不同程度的萎蔫。盐胁迫使4个小麦品种的总叶绿素含量和相对含水量均有所下降, 但矮变1号的下降幅度最小; 与此同时, 矮变1号体内SOD活性的相对上升幅度最大, 并且体内的MDA含量相对上升幅度最小。4个品种中矮变1号的DELLA蛋白积累量最高, 矮变1号具有较高的耐盐性与其体内的高DELLA蛋白含量密切相关。
, FAN Xiao-Cong, SONG Mei-Fang, YANG Jian-Ping
DELLA proteins are negative regulators in GA signal pathway. DELLA protein is able to response to various environmental signals and plays an important role in plant resistance to biotic and abiotic stresses. The DELLA gain-of-function mutant Aibian 1 has been widely applied in wheat breeding for dwarfness, but its tolerance to salt is unclear. In this study, Aibian 1 was compared with Chinese Spring, Jing 411, and Aiganzao in salt tolerance using Hoagland’s hydroponic culture with 200 mmol L-1 NaCl. Total chlorophyll content, relative water content, superoxide dismutase (SOD) activity and malondialdehyde (MDA) content were measured to evaluate the ability of salt tolerance. Immunoblot assay was employed to assess the DELLA protein abundance in wheat seedlings. After seven-day NaCl treatment, Aibian 1 showed no clear withering leaves, whereas the other three cultivars displayed obviously wilting and yellowing leaves. Among the four cultivars, Aibian 1 had the least loss on chlorophyll content and relative water content, the highest SOD activity, and the lowest levels of MDA content after salt treatment. Immunoblot assay indicated the highest accumulation of DELLA protein in Aibian 1 among the four cultivars. Thus, we consider the high level of DELLA protein is closely related to salt tolerance in Aibian 1.
我国农用耕地中约有1亿公顷的盐碱地, 是制约小麦播种面积和进一步增产增收的重要环境因素之一[1]。因此, 开展小麦抗盐机理研究、培育耐盐品种对我国扩大小麦播种面积、高产稳产, 乃至粮食安全具有重要的意义。
20世纪60年代, 随着矮秆基因Rht的广泛应用, 兴起了第一次“ 绿色革命” 。目前, 已定名的Rht主效矮秆基因多达25个[2], 但在生产中有利用价值的却不超过10个, 利用最广泛的是来源于农林10号的Rht1 (Rht-B1b)和Rht2 (Rht-D1b), 分别位于小麦4B和4D染色体短臂[3]。Rht1和Rht2编码赤霉素(GA)信号途径中的负向调控因子DELLA蛋白。该类蛋白主要存在于细胞核内[4], 属于转录调控因子GRAS (GAI、RGA和SCR)蛋白家族的一个亚家族[5]。DELLA蛋白主要由位于N端的GA信号感知区和近C端的GA信号调节区两部分构成。GA信号感知区包含2个非常保守的酸性结构域VHYNP和感受GA信号所必须需的DELLA结构域[6]; GA信号调节区由NLS (nuclear localization signal)、LZ (Leucine zipper)结构域、poly S/T/V (poly Ser/Thr/Val)调节结构域、VHIID (Val-His- Ile-Ile-Asp) [7, 8]、SAW (SCARECROW)等功能域构成[4]。植物感知GA信号后, 位于质膜上的GA受体GID1与GA形成GA-GID1复合体, 该复合体进入核中与DELLA蛋白的N端结合后, DELLA蛋白的C端GRAS结构域识别E3连接酶SCFSLY1/GID2的F-box亚基, 从而形成GA-GID1-DELLA-E3连接酶复合体, 导致DELLA蛋白通过26S蛋白酶体途径降解, 打开GA信号途径, 从而调控植物的生长发育进程[9]。小麦Rht-B1b和Rht-D1b两个基因编码的蛋白均是N端DELLA区段的缺失导致GA信号感知区遭到破坏, DELLA蛋白不能被降解, 从而阻断GA信号转导, 致使小麦植株矮化[10]。
DELLA蛋白不仅参与调节植物的株高[11], 还能够响应各种逆境胁迫, 在作物抵抗逆境中发挥着重要的作用。盐胁迫条件通过诱导GA2氧化酶导致内源GA水平降低、DELLA蛋白大量积累、植物生长受抑、植物抗盐能力提高[12, 13, 14]。响应低温的转录因子CBF1/DREB1b能够提高GA合成途径中GA2ox3和GA2ox6的表达水平, 从而减少内源GA的含量, 稳定DELLA蛋白, 提高植物的抗冷能力[15]。植物营养不足, 例如磷缺乏, 导致GA含量降低, 促进DELLA蛋白的积累, 从而调控植物低磷胁迫下的适应能力[16]。已经发现的DELLA蛋白包括拟南芥的GAI、RGA、RGL1、RGL2和RGL3, 大麦的SLN1, 水稻的SLR1和OsGAI, 小麦的Rht和葡萄的L1等[17]。对拟南芥DELLA蛋白的抗逆研究有很多报道, 但对小麦DELLA蛋白目前主要报道其株高调节功能, 鲜有其他方面的研究。
矮变1号是西安市农业科学研究所于1972年从小麦品种矮秆早中选出的天然Rht突变体, 株高26~29 cm, 是世界上最矮的小麦品种之一[18]。该突变体携带不完全显性矮杆基因Rht-D1c, 由2个拷贝的Rht-D1b串联复制而成, 其致矮效果是Rht-D1b的3倍以上[3]。利用矮变1号与太谷核不育杂交选育出矮败小麦, 并且已建立了矮败小麦轮回选择育种体系[19]。本研究采用含NaCl (200 mmol L-1)的Hoagland营养液处理4个小麦品种的幼苗7 d, 比较其总叶绿素含量、相对含水量、SOD活性和MDA含量等生理指标, 并借助免疫印迹检测DELLA蛋白水平, 明确DELLA蛋白与小麦耐盐性之间的关系, 为探讨矮变1号在抗盐育种中的利用价值提供依据。
4个供试品种均由中国农业科学院作物科学研究所提供, 本实验室扩繁并保存。其中, 矮秆早由麦类种质资源课题组提供, 中国春、京411和矮变1号由矮败小麦课题组提供。
实验在中国农业科学院人工温室内进行, 以消除光照和温度等条件造成的误差。培养条件为16 h光照/8 h黑暗, 光照强度30.0 µ mol m-2 s-1, 恒温22℃, 空气相对湿度50%。选取无病饱满的小麦种子, 表面消毒后用无菌水浸种12 h, 28℃催芽24 h, 将萌动露白一致的种子播于蛭石中。4 d后挑选生长健壮、株高一致的幼苗, 洗净根部, 用长条海绵固定于带孔的泡沫板上, 并于Hoagland营养液(pH 6.0)中培养。将4个品种置同一块泡沫板上, 每品种重复3孔, 每孔2株。幼苗在营养液中适应2 d后, 转入含200 mmol L-1 NaCl的营养液中, 以不含NaCl的营养液为对照。每3 d更换一次营养液, 处理7 d后取叶片测定各种生理指标。设3次独立的生物学实验。
未经NaCl处理时, 4个品种均生长良好, 叶片为绿色且竖直向上生长(图1-A, C); 而200 mmol L-1 NaCl处理7 d后, 中国春、京411和矮秆早的株高明显下降, 叶宽变窄, 叶片萎蔫与失绿的程度依次加重, 而矮变1号的叶色与生长状况与未胁迫处理的生长状况无明显差异, 也没有出现明显萎蔫(图1-B, D)。
 | 图1 盐胁迫前后的小麦幼苗形态 小麦种子28℃催芽24 h后在蛭石中22℃生长4 d (A和B), 然后在Hoagland液(pH 6.0)中缓冲2 d, 再分别转入不含(C)或含(D) 200 mmol L-1 NaCl的Hoagland培养液(pH 6.0)生长7 d。E图是D图方框部分的局部放大。1: 中国春; 2: 京411; 3: 矮秆早; 4: 矮变1号。Bar = 2 mm。Fig. 1 Morphology of wheat seedlings in response to salt stress Wheat seeds were germinated at 28 ° C for 24 h and grown in vermiculite for 4 d in greenhouse at 22 ° C (panels A and B). Wheat seedlings were carefully transferred into the Hoagland’ s solution (pH 6.0) for 2 d to buffer and finally cultured in Hoagland’ s solution without (panel C) or with (panel D) 200 mmol L-1 NaCl for 7 d. Panel E is the enlargement of framed parts in panel D. 1: Chinese Spring; 2: Jing 411; 3: Aiganzao; 4: Aibian 1. Bar = 2 mm. |
未经NaCl胁迫处理的4个品种的总叶绿素含量和相对含水量均处于较高水平, NaCl处理后呈明显下降趋势(图2), 总叶绿素含量下降幅度为中国春63%、矮秆早45%、京411 20%、矮变1号17%, 相对含水量下降幅度为中国春27%、矮秆早14%、京411 12%、矮变1号4%。可见, 矮变1号耐盐胁迫的能力较强。
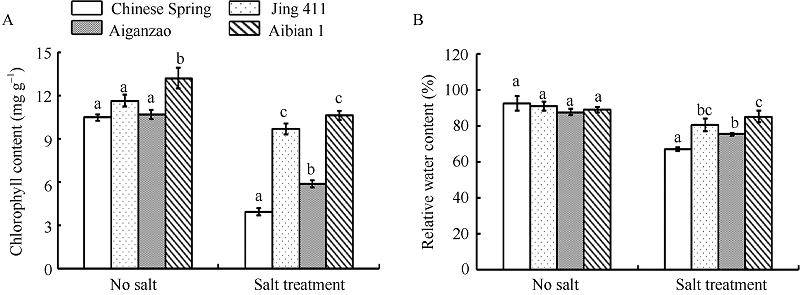 | 图2 盐胁迫和无胁迫下4个小麦品种的叶绿素含量(A)和相对含水量(B)变化 数据为3次生物学重复的平均值和标准差。Fig. 2 Changes of chlorophyll content (A) and relative water content (B) in four wheat cultivars under treatment with or without salt stress Data are the means and standard deviations of three biological repeats. |
NaCl处理后, 小麦幼苗中MDA含量和SOD活性较未胁迫处理时升高(图3), 其中MDA含量增加幅度为中国春2.9倍、矮秆早1.9倍、京411 1.3倍、矮变1号1.1倍, SOD活性增长率为中国春23%、矮杆早25%、京411 38%、矮变1号46%, 表明矮变1号清除自由基和稳定细胞膜的能力最强。
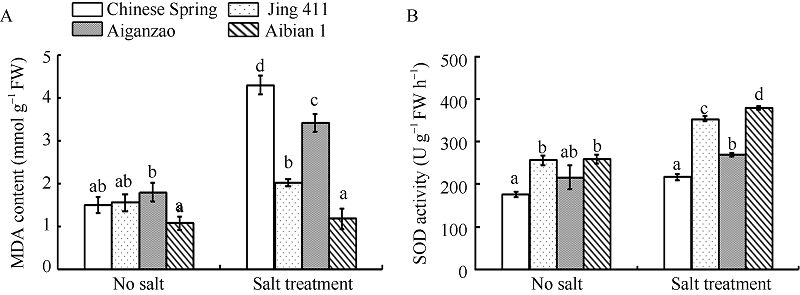 | 图3 盐胁迫和无胁迫下4个小麦品种中MDA含量(A)和SOD活性(B)的变化 数据为3次生物学重复的平均值和标准差。Fig. 3 Changes of MDA content (A) and SOD activity (B) in four wheat cultivars under treatments with or without salt stress Data are the means and standard deviations of three biological repeats. |
不论在黑暗还是在白光条件下, 矮变1号的DELLA含量均明显高于其他3个品种(图4), 结合形态学和生理生化指标, 认为矮变1号的耐盐性与其体内较高的DELLA蛋白表达水平密切相关。
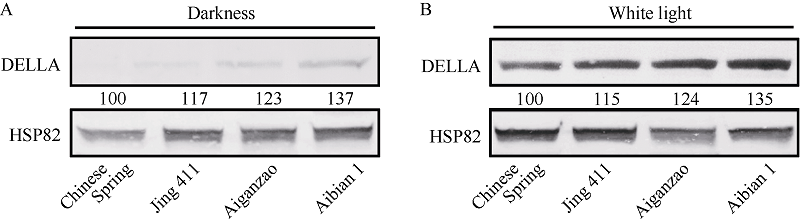 | 图4 4个小麦品种的幼苗在白光或黑暗下DELLA蛋白的免疫印迹检测 A: 22℃白光(30.0 µ mol m-2 s-1)生长6 d; B: 22℃黑暗生长6 d。HSP82免疫印迹显示大约相当的上样量。Fig. 4 Immunoblot analysis of DELLA protein accumulation in four wheat cultivars under white light or darkness A: Seedlings were grown for six days under 22° C and white light (30.0 µ mol m-2 s-1); B: Seedlings were grown for six days under 22℃ and darkness. Anti-HSP82 immunoblots indicates approximately equal loading. |
高盐对植物造成的最直接的伤害是水分胁迫, 植物会因环境的渗透势低于细胞内渗透势导致细胞逐渐失水, 失水量大于吸水量时, 植物出现卷曲和萎蔫症状[24, 25]。叶绿体是植物对盐分最敏感的细胞器, 盐分破坏细胞中色素-蛋白质-类脂复合体的结合强度, 并且增强叶绿素降解酶的活性, 破坏细胞膜系统、促进叶绿素的降解, 导致叶片失绿和黄化, 光合作用受阻, 植物正常的能量供应失调, 造成植物的生长和发育紊乱[24, 26]。本研究发现, 矮变1号在盐胁迫处理前后未出现明显的萎蔫和失绿, 并且植株生长状态良好(图1), 体内的总叶绿素含量和相对含水量均高于其他3个小麦品种(图2), 表明矮变1号缓冲胁迫、维持相对正常的生长发育的能力更强。
在遭受盐胁迫时, 植物细胞内自由基积累, 导致膜脂过氧化水平增高, 体内MDA含量增加引起蛋白质、核酸等生命大分子交联聚合, 细胞内膜系统的完整性遭到破坏, 胞内组分大量外渗, 正常代谢不能维持而导致植物受伤表现出逆境反应乃至死亡。植物体内的SOD等保护性酶类的活性在植物遭遇逆境胁迫时大幅提高, 以减轻MDA积累的伤害, 维持细胞膜系统的稳定性与完整性[27, 28]。胁迫处理后, 矮变1号的SOD活性比处理前增长46% (图3), 明显大于其他3个品种的SOD增幅, 同时矮变1号的MDA上升幅度仅为10%, 变化幅度远低于其他3个品种。这表明在盐胁迫下, 矮变1号通过提高自身SOD酶的活性来增强清除活性氧的能力, 从而增强对盐胁迫的耐受能力。
DELLA蛋白作为GA信号途径中的重要调节因子, 也是外界环境信号与其他植物激素信号通路中一个重要的节点蛋白[29], 能够响应光、乙烯、ABA、非生物胁迫及生长素途径[30]。在拟南芥中, 盐胁迫下DELLA蛋白通过提高SOD等抗氧化酶的活性使植物体内的活性氧(ROS)保持在较低的水平, 从而提高植物的抗盐能力[31]。矮变1号携带Rht-D1b基因, 该基因内部发生碱基替换, GGA突变为TGA, 形成终止密码子, N端的GA信号感知区遭到破坏, 导致DELLA蛋白不被降解[3, 10]。在本研究中, 矮变1号的DELLA含量高于其他3个品种(图4), 且盐处理后矮变1号的SOD活性上升幅度最大, MDA含量上升幅度最小(图2), 表明由DELLA蛋白调控的耐盐机制在小麦和拟南芥中相似, 该机制可能普遍存在于大多数植物中。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|