


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
芥菜型油菜多室基因 Bjmc2的精细定位
[王刚, 张向向, 徐平, 吕泽文, 文静, 易斌, 马朝芝涂金星, 傅廷栋, 沈金雄*  ]
]
]
|
|


第一作者联系方式: E-mail: wanggang2014@webmail.hzau.edu.cn, Tel: 18963949812
芥菜型多室油菜的产量比普通两室油菜更高, 定位乃至克隆多室基因可为油菜遗传改良及解释多室角果形成机制创造条件。本研究通过验证JD11-2家系衍生群体仅在 BjMc2位点上存在差异, 可用于 BjMc2的定位。采用AFLP结合BSA法分析BC5和BC6群体, 筛选到1个与 BjMc2连锁的AFLP标记并转化为SCAR标记SC1。基于该AFLP标记序列信息, 利用白菜同源序列设计SSR引物和SCAR引物, 获得11对SSR标记和1对SCAR标记。通过在芥菜型油菜BAC文库中的挑选, 获得2个覆盖目标区域的单克隆, 由此开发1个SSR标记。将获得的SCAR和SSR标记扫描BC7群体, 构建了两室性状基因 BjMc2的遗传连锁图, 两侧最近标记ZX17和BACsr96与目标基因之间的遗传距离分别为0.048 cM和0.340 cM, 并定位到白菜A7 Scaffold 000019的946~1014 kb之间, 约68 kb物理距离。
Multilocular plants have a higher seed yield than bilocular plants in B. juncea, mapping and cloning the multilocular gene(s) might be helpful for rapeseed genetic improvement and explanation of the multilocular trait development mechanism. This study verified backcross populations derived from JD11-2 family different at BjMc2 locus only can be used for BjMc2 mapping. Using AFLP assay and BSA method in BC5 and BC6 generations, one AFLP marker linked to the target gene was obtained and converted to SCAR marker (SC1). On the basis of homologous sequences of the AFLP maker in B. rapa, 11 SSR markers and one SCAR marker were identified. Through the screening in the ZBjH BAC library of Brassica juncea, two BACs covered the target area were selected and one SSR marker was developed. All the developed SCAR and SSR markers were then used to detect the BC7 population, and a linkage map for the bilocular gene BjMc2 was built. ZX17 and BACsr96, the closest flanking markers, were mapped at 0.048 cM and 0.340 cM distant from the BjMc2 gene, respectively. Bjmc2 is positioned of 68 kb between 946 kb and 1014 kb in the Scaffold 000019 physical map of A7 in B. rapa. This result would lay a foundation for cloning polycyetic gene Bjmc2and selecting polycyetic lines by marker-assisted selection.
油菜是重要的油料作物, 高产是油菜遗传育种的首要目标。油菜单株产量构成因素主要包括全株角果数、每角粒数和千粒重。宋稀等[1]和Zhu等[2]认为今后一段时间增加每角粒数是油菜改良重点之一。普通油菜角果由两心皮发育成两室, 中间为假隔膜, 每角粒数一般15~20粒, 极少数品种超过30粒。而多室油菜角果由3个以上心皮发育而成, 中间至少存在2个假隔膜将果室分隔为三室、四室或五室, 每角粒数多在30粒左右, 最多可达40余粒[3, 4, 5, 6]。显然, 多室材料的显著特征是角果粒数多于两室材料。Katiyar等[3]、赵洪朝等[4]和吕泽文等[7]研究表明, 尽管多室油菜的千粒重和单株有效角果数均低于普通两室油菜, 但因多室油菜每角粒数更多, 结果多室材料的单株或小区产量均显著高于两室材料(一般高10%~40%, 最高达70%)。因此, 研究和利用多室性状对提高油菜产量具有重要意义。
定位甚至克隆多室基因是研究和利用多室性状的重要基础。分子标记技术已广泛用于油菜重要农艺性状相关基因的图谱定位与克隆。由于油菜属于异源四倍体作物, 基因组比较复杂, 开发连锁标记较困难, 目前油菜中仅有少数几个多室基因被定位甚至克隆。Paritosh等[8]以白菜型多室材料黄籽沙逊Tetra及其与Chiifu杂交后代衍生的RIL群体为材料, 利用RNA-seq开发了大量SNP标记, 将多室基因tet-o定位到A04染色体, 两侧最近的SNP标记分别为0.1 cM和0.3 cM。Fan等[9]证实位于A04染色体上的BrCLV3是控制白菜型油菜黄籽沙逊ml4突变体多室表型的关键基因, 由于BrCLV3的第3个外显子区单碱基突变, 致使CLV-WUS负调控途径异常, 最终导致多室角果的形成。Xiao等[10]认为芥菜型油菜duoshi的多室性状遗传受两对独立遗传的隐性核基因控制, 并将多室基因Bjln1定位到A7染色体的208 kb区段内。吕泽文[11]研究认为芥菜型油菜J163-4多室性状受2对隐形核基因(Bjmc1和Bjmc2)控制, 无细胞质效应, 且开发了与Bjmc1基因连锁的SSR、SCAR标记, 分别距目标基因11.5 cM和14.9 cM。Xu等[12]在吕泽文研究基础上扩大群体将Bjmc1定位在A7的一个区段内, 其两侧最近的标记距离分别为1.1 cM和1.6 cM, 而对于多室基因Bjmc2还未开发连锁标记。近几年, 多种芸薹属作物的全基因组测序相继完成, 并建有相应数据库, 为开发与Bjmc2连锁的分子标记提供了捷径。
本研究通过构建BjMc2位点回交群体, 拟利用AFLP标记技术及BSA法、芸薹属数据库, 结合田间表型数据计算标记遗传距离, 旨在构建BjMc2位点的遗传连锁图; 从而为Bjmc2的克隆和分子标记辅助选育多室品种奠定基础。
亲本材料是芥菜型油菜两室材料J248-2和多室材料J163-4, 均由华中农业大学油菜研究室杂优组提供, 均经多代连续自交, 遗传稳定。Bjmc2位点的定位群体的构建方法如图1, 由J248-2与J163-4杂交得F1, F1与J163-4回交两代, 选择BC2世代中JD11-2家系(其两室单株与多室单株比例为1∶ 1), 其后选择两室单株与多室单株进行多代兄妹交保存该家系。
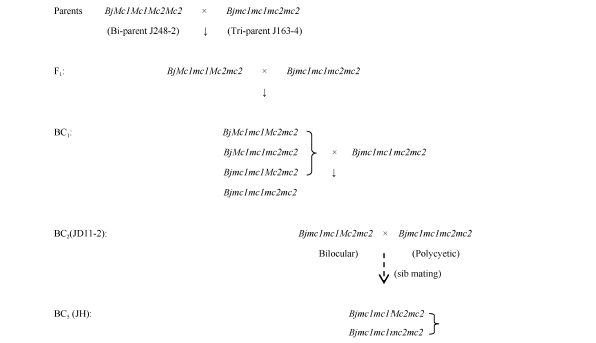 | 图1 芥菜型油菜多室位点Bjmc2群体的构建Fig. 1 Construction of populations with polycyetic trait gene Bjmc2 in Brassica juncea |
种植由JD11-2家系两室单株与多室单株多代兄妹交繁衍的BC5(JH)世代, 然后用与BjMc1连锁的分子标记(SR02, 340 bp[12])验证该群体, 并且选择两室单株套袋自交, 另外还与构建的BjMc1位点回交分离群体中两室单株杂交。在其后代家系中, 选两室单株自交且同时与多室单株兄妹交。角果表型分离的均用χ 2测验, 以检验其分离比适合性。
在芥菜型油菜苗期选取新鲜嫩叶, 采用CTAB小样法提取基因组DNA[13]。用分光光度计测定DNA溶液浓度, 用超纯水将浓度稀释到50 ng μ L-1。根据BSA法, 将BC5(JH)世代等量的18株两室单株和18株多室单株的DNA溶液分别混成3个两室Bulk池和3个多室Bulk池(6株为一池)用于AFLP和SSR标记的筛选[14]。
采用Negi等[15]方法并略有改动筛选AFLP标记。采用EcoR I/Mse I、Pst I/Mse I两种组合, 充分酶切后, 加入接头和连接缓冲液, 4℃过夜, 将连接产物稀释10倍后作为预扩模板, 采用EA/MC、EA/MG、EC/MA、EC/MG、P0/MC和P0/MG组合预扩, 将预扩产物稀释20倍作为选择性扩增模板。用选扩引物扩增后的样品100℃水浴变性后采用6%聚丙烯酰氨凝胶电泳分离检测。
参考Yi等[16]方法将筛选的AFLP标记差异片段挖胶回收并测序, 用Lasergene软件将序列去载体, 和对应选扩引物比对得到差异片段序列, 利用Primer5软件设计引物, 检测两室基因池和多室基因池是否存在差异。
将差异AFLP标记序列在Brassica Database (BRAD)数据库中进行Blast比对分析, 根据序列信息筛选白菜scaffolds, 并由此利用http://wsmartins. net/websat/网站设计SSR引物, 用两室和多室基因池进行筛选, 参照吕泽文[11]硕士论文的PCR反应体系和参数。
用与BjMc2紧密连锁的标记筛选湖南农业大学构建的芥菜型油菜ZBjH BAC文库[17]。由上海生工釆用Sanger方法进行BAC末端测序, 测序引物为M13R和S1。在Brassica Database (BRAD)数据库中以Blast比对分析获得的BAC末端序列。釆用二代Illumina测序平台进行BAC全长测序, 并进行Denovo组装全长。得到目标序列后再利用http:// wsmartins.net/websat/网站设计SSR引物, 用两室和多室Bulk池进行筛选, 以获得多态性标记。
利用BC5(JH)世代301单株、BC6世代252单株和BC7世代6207单株检测所有与BjMc2连锁的SCAR、SSR标记, 根据田间角果表型计算连锁标记与目标基因的遗传距离, 用MapDraw软件作遗传连锁图[18]。
由表1和表2可见, BC5(JH)世代群体中两室单株自交, 第1代两室和多室单株数比例接近3∶ 1; 第2代中, 两室自交的后代中两室和多室单株数比例为3∶ 1或全是两室单株, 未出现两室和多室单株数比例为15∶ 1; 与多室兄妹交的后代中两室和多室单株数比例为1∶ 1或全是两室单株。同样, BC5(JH)世代中两室单株与BjMc1位点的回交分离群体中两室单株杂交, 第1代两室和多室单株数比例接近3∶ 1; 第2代中, 两室自交后代中两室和多室单株数比例为15∶ 1或3∶ 1; 与多室兄妹交后代中两室和多室单株数比例为3∶ 1或1∶ 1。另外, 如图2用BjMc1基因连锁标记SR02鉴定BC5(JH)回交群体, 两室和多室单株未出现差异。以上结果进一步验证芥菜型油菜J163-4多室性状由两对基因控制, 两室相对多室为显性; 还证实由JD11-2家系衍生的BC5(JH)回交群体中两室和多室单株仅在BjMc2位点上存在差异, 可用于构建群体定位BjMc2。
| 表1 BjMc1与BjMc2间的等位性分析(自交或杂交第1代分离表现) Table 1 Allelic analysis between BjMc1 gene and BjMc2 gene (segregation in selfing or hybrid first generations) |
| 表2 BjMc1与BjMc2间的等位性分析(自交或杂交第2代分离表现) Table 2 Allelic analysis between BjMc1 gene and BjMc2 gene (segregation in selfing or hybrid second generations) |
 | 图2 与BjMc1连锁标记SR02在不同基因型单株中的扩增1~2:BjMc2位点定位群体中两室单株和多室单株; 3~4:BjMc1位点定位群体中两室单株和多室单株。Fig. 2 Amplification in diverse genotypes using marker SR02 linked toBjMc11-2: bilocular and polycyetic plants in population with BjMc2; 3-4: bilocular and polycyetic plants in population with BjMc1. |
通过筛选1536对AFLP引物组合, 获得1个与BjMc2连锁的AFLP标记(P10MC4)(图3)。进行差异片段回收测序, 在Brassica Database (BRAD)数据库中Blast比对分析序列, 结果与白菜A7同源性很高, 比对在白菜Scaffold 000019的1435 kb位置上, 根据序列设计引物成功转化成SCAR标记(SC1) (图4)。
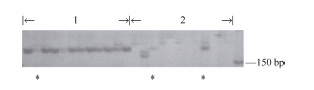 | 图3 AFLP引物组合P10MC4在12个两室单株和12个多室单株中的扩增情况1~2: 两室单株和多室单株, * 为交换单株。Fig. 3 AFLP primer pair P10MC4 profiles from 12 biloculars and 12 triloculars1-2: bilocular and polycyetic plants, * recombinant type. |
 | 图4 标记SC1在bulk池和单株的扩增结果1~2: 两室bulk池和多室bulk池。Fig. 4 Amplification results of SC1 on bulk pools and individuals plants1-2: bilocular bulk pools and polycyetic bulk pools. |
由AFLP标记(P10MC4)的差异片段回收测序所得序列, 根据比对结果在Scaffold 000019的1435 kb位置左右每约100 kb设计1对SSR引物, 且在Brassica Database (BRAD)数据库下载白菜A7同源区段的已有SCAR标记。用3个两室Bulk池和3个多室Bulk池去验证这些引物, 获得11对SSR标记和1对SCAR标记(SC8)与BjMc2连锁(表3)。
| 表3 与BjMc2连锁的SSR标记和SCAR标记的特征 Table 3 Characterization of SSR and SCAR markers linked to BjMc2 |
用3个BjMc2两端较近的连锁标记(ZX4、SSR344和ZX10)在芥菜型油菜ZBjH BAC文库中, 筛选到2个BAC单克隆(002-O-21和009-M-2)能够覆盖目标基因区域。BAC末端测序结果都能比对到Scaffold 000019上, 002-O-21两端分别比对在832 kb和979 kb; 009-M-2两端分别比对在1104 kb和941 kb。推测002-O-21 BAC单克隆约150 kb, 009-M-2 BAC单克隆约160 kb, 与该文库平均长度插入相符。由BAC全长测序结果开发更近的SSR标记, 成功得到BACsr96标记与BjMc2连锁。
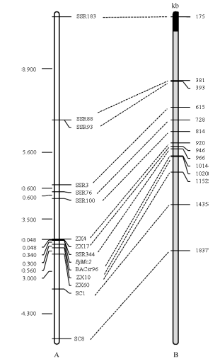
用BC5(JH)世代301个单株、BC6世代252个单株和BC7世代6207个单株检测所有SCAR、SSR标记与BjMc2的遗传距离, 由构建的遗传连锁图可知, 所有连锁标记覆盖在BjMc2周围27.196 cM区域内, 包括12个SSR标记和2个SCAR标记。在所有连锁标记中, ZX17和BACsr96与BjMc2遗传距离最近, 分别为0.048 cM和0.340 cM, 其中SSR344与目标基因共分离(图5-A)。
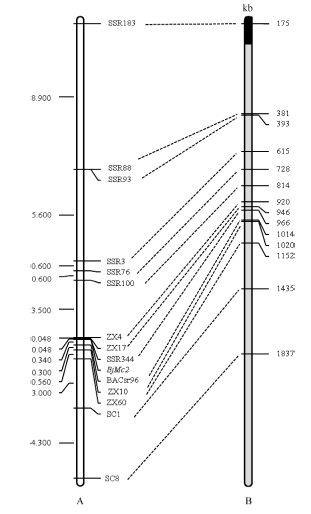 | 图5 芥菜型油菜两室基因BjMc2的遗传连锁图和白菜A7同源Scaffold物理图谱A是BjMc2的遗传连锁图; B是白菜A7同源Scaffold物理图谱, 黑色代表Scaffold 000101, 灰色代表Scaffold 000019。Fig. 5 Genetic linkage map of bilocular gene BjMc2 inB .junceand physical map of homologous Scaffolds of A7 in B. rapaA is the genetic linkage map of the bilocular gene BjMc2; B is physical map of homologous Scaffolds of A7 in B. rapa, black represent Scaffold 000101, gray represent Scaffold 000019. |
因多室油菜能提高产量, 故越来越受到研究者关注[4, 5, 6, 7, 8, 9, 10, 12, 19, 20]。目前报道的几个油菜多室材料, 其表型及稳定性都不尽相同。何余堂等[5]发现白菜型多室油菜有多种表型(三室和四室), 且同一单株有两室、三室和四室并存现象(嵌合现象), 早期子房由4个心皮组成, 子房横切面假隔膜呈“ +” 型, 将子房
分隔成大小均匀的4个腔室, 但在后期发育过程中假隔膜退化, 有的退化为三室, 横切面呈“ Y” 型; 有的直接退化为两室。Paritosh等[8]、Fan等[9]研究材料同样存在嵌合现象。朱彦涛等[6, 19]发现甘蓝型多室油菜三棱角果横截面为三角形, 假隔膜呈现“ Y” 型, 但该材料也存在三棱角果与正常两室角果同时存在的嵌合现象。赵洪朝等[4, 20]、Xiao等[10]报道的芥菜型油菜多室材料duoshi其角果横切面的假隔膜呈“ +” 型, 且其表型不稳定, 存在两室和多室嵌合现象。而本研究利用的芥菜型油菜多室材料J163-4, 其子房横切面的假隔膜呈“ II” 型, 与上述白菜型油菜、芥菜型油菜及甘蓝型油菜多室材料的横切面的假隔膜形态均不同且表型稳定, 未发现有两室和多室角果并存的嵌合现象[7, 12]。J163-4的表型和稳定性不同于其他油菜多室材料可能与其受控基因的不同/或基因结构变异有关。
本研究开发的所有标记都与白菜A7同源, 且将J163-4多室性状基因Bjmc2定位到2个最近连锁标记ZX17和BACsr96之间, 遗传距离分别为0.048 cM和0.340 cM。这2个标记在白菜Scaffold 000019同源位置是946 kb和1014 kb (~68 kb) (图5-B)。此前Xu等[12]报道BjMc1定位区段也与A7同源, 但等位性分析和SR02标记检测结果表明BjMc1和BjMc2是2个独立分离的位点。2个位点区域都与A7同源可能是由于A、B和C基因组本身有同源关系或同一个基因在不同基因组有不同拷贝。Truco等[21]对3个二倍体物种的连锁图比较发现, 3个基因组间有着广泛的相同保守区域, 但伴随着大量的序列重排现象, 也广泛存在A、B、C基因组内部的同源保守区段, 由此推测出3个基因组的形成可能是由一个基因组较小的祖先物种经过多次重复和重排形成的。现已完成甘蓝型油菜全基因测序[22], 借助与亲本种甘蓝和白菜基因组的比较, 发现该基因组进化的突出特点是2个亚基因组(A和C)存在着广泛的相互基因或DNA序列置换, 这从序列上解释了甘蓝型油菜异源四倍体进化关系。另外, Xiao等[10]报道的芥菜型油菜duoshi的多室基因Bjln1定位区段也与白菜A7 Scaffold 000019同源。但是, 由于多室材料J163-4和duoshi的角果表型不同, 且J163-4的多室表型稳定, 均为三室, 不存在嵌合现象, 因此Bjmc2与Bjln1尽管定位在同一区段, 可能为同一位点/等位基因, 但其基因结构不同, 基因表达效应不同, 最终导致表型和稳定性明显差异。再者, 本研究把BjMc2定位在Scaffold 000019的946~1014 kb区段, 该区段可能含有参与拟南芥顶端分生组织发育反馈调控途径(CLV-WUS)关键基因CLV1的同源基因Bra015812 [Brassica Database (BRAD)数据库]。关于clv1, 在拟南芥中已有大量研究和报道, 其突变能够导致子房出现多心皮, 从而出现角果多室性状[23, 24]。在拟南芥中clv1的clv1-1[23]、clv1-2至clv1-3[24]、clv1-4至clv1-7[23]、clv1-8至clv1-9[24]和clv1-10至clv1-13[25]的不同突变体的表型均不同, 心皮数为3~9个。在芥菜型油菜中, CLV1同源拷贝数及突变类型可能会更多, 故在异源四倍体的芥菜型油菜中, 该反馈调控途径会更复杂, 不同的材料可能是不同的拷贝或突变类型。因此, 只有克隆多室基因, 分析基因结构和表达模式才能更好地研究和利用多室性状。另外, 通过已有优良芥菜型油菜或甘蓝型油菜品种与J163-4杂交, 再利用本文已开发与BjMc2紧密连锁的分子标记检测后代单株, 可以辅助选育含有多室基因的家系, 从而加快选育芥菜型油菜或甘蓝型油菜多室品种。
通过JD11-2衍生的回交定位群体, 将BjMc2锚定到遗传距离分别为0.048 cM和0.340 cM的ZX17和BACsr96标记内, 与白菜Scaffold 000019的同源位置是68 kb物理距离。
致谢: 本研究所用芥菜型油菜BAC文库得到湖南农业大学刘忠松教授的帮助, 在此谨表谢意。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|