
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
烟草ATP合酶F0部分4个亚基基因转录本编辑位点分析
[陶瑶1 , 王瑜1, 3 , 钟思荣1 , 吴凌敏1 , 谢丽娟1 , 聂亚平1 , 周玮2 , 王建革4 , 刘齐元1, *  ]
]
]
|
|


第一作者联系方式: E-mail: 1240447330@qq.com
RNA编辑是高等植物线粒体基因转录后表达调控的一种重要方式。为探究ATP合酶F0部分的4个亚基基因与植物雄性不育性的关系, 本研究以3个烟草雄性不育系(MS中烟90、MS云烟85和MS K326)及其同型保持系为供试材料, 比较分析 atp6、 atp9、 orf25和 orfB线粒体基因转录本的编辑位点。结果表明, orf25和 orfB基因转录本在不育系和保持系中发生的编辑位点是一致的。 atp6基因在不育系中未发生编辑, 在保持系中共有6处发生了RNA编辑。与保持系相比, atp9基因在不育系中除8处共同的C→T编辑外, 还缺少2个C→T的特异编辑位点, 其中1个导致氨基酸类型的改变。推测不育胞质因缺少特异的线粒体基因转录本编辑而导致烟草的细胞质雄性不育。
RNA editing exits extensively in mitochondria of higher plants and is one of the most important post-transcriptional regulation methods of gene expression in mitochondrial genomes of higher plants. At the same time, it is an essential process for forming function proteins. RNA editing can induce mutations in mitochondrial genes including nucleotide insertion, substitution, or deletion, which further affects the splicing and processing of primary transcripts, ultimately resulting in cytoplasmic male sterility (CMS). The results of research using multiple species showed that there is an obvious relationship between the four subunit genes of F0-ATPase and CMS. To explore the relationship, we studied RNA editing status of four mitochondrial genes atp6, atp9, orf25, and orfB from three tobacco male sterility lines (MS Zhongyan 90, MS Yunyan 85, MS K326) and their corresponding fertile lines . The four mitochondrial genes atp6, atp9, orf25, and orfB and their cDNA were distinctively amplified by PCR from six tobacco lines. After that, by means of making a comparison between the DNA sequences and the cDNA sequences of target genes to find RNA editing sites. The orf25and orfB gene transcripts had the same RNA editing sites between male sterile and fertile lines. For atp6 gene, RNA editing didn't occur in male sterile lines, while there were six RNA editing sites in fertile lines, which all caused changes in the type of amino acids and there were four editing sites enhancing hydrophobicity of the amino acids. It inferred that the difference of protein's hydrophobicity was most likely to cause CMS. The atp9 gene had ten RNA editing sites in fertile lines, eight of which were the same as those in male sterile lines, while two C→T unique editing sites were absent in male sterile lines, of which one caused changes in amino acid. The nucleotide variations at 223 site of atp9 gene resulted in producing a termination code, which might be the necessary RNA editing to produce normal functional protein. These results suggest that lacking of the unique RNA editing sites might contribute to CMS property in tobacco.
RNA编辑是指DNA转录成RNA后通过核苷酸的删除、插入或替换等方式改变遗传信息的一种普遍现象, 是线粒体基因转录后调控的重要过程, 是对中心法则的重要补充[1]。RNA编辑现象普遍存在于植物、动物、微生物等各生物有机体中, 在细胞核、线粒体和叶绿体具有自身遗传系统的细胞器中均可发生。植物线粒体RNA编辑是具有普遍生物学意义的, 它的发现合理解释了线粒体中存在的标准密码使用规律不一致的现象[2]。研究表明, 绝大多数的RNA编辑会导致氨基酸类型的改变, 进而使基因组的遗传信息不断丰富。编辑作用可构建或除去起始或终止密码子, 改变转录本的长度[3]。此外, 蛋白质编码区的编辑一定程度上还能提高转录本的稳定性和编码蛋白的疏水性, 由此可见RNA编辑已成为线粒体产生功能蛋白所必不可少的过程, 同时也成为细胞核调控线粒体基因表达的重要方式之一[4]。
细胞质雄性不育(cytoplasmic male sterility, CMS)广泛存在于高等植物中, 其在杂种优势利用和研究植物发育过程中核质互作方面具有重要的理论和实际意义[5, 6, 7, 8]。国内外大量研究结果表明, 不育和可育胞质的线粒体是有差别的, 随着人们对植物生殖发育的分子生物学和植物雄性不育机制的深入研究, 使人们确信线粒体基因组分子内或分子间频繁重组所形成的异常嵌合基因或开放阅读框是产生CMS的分子基础。近年研究表明, CMS与高等植物线粒体基因的RNA编辑有重要关系[9, 10]。在RNA编辑中由于碱基的插入、缺失或替代, 进而影响初始转录物的剪接和加工, 导致基因结构或表达调控模式改变, 或者形成嵌合基因等, 这些异常的编辑产物可能会导致线粒体功能不能正常发挥而形成CMS[11]。此外, 异常的ATP酶复合体也会导致线粒体的功能缺陷, 从而导致雄性不育[12, 13]。atp6、atp9、orf25和orfB是组成线粒体ATP合酶F0部分的4个亚基, 而F0部分是ATP合酶酶蛋白的膜内区域, 具有质子跨膜传输功能, 在线粒体合成ATP时起很重要的作用。多个物种的研究结果表明, ATP合酶F0部分的4个亚基基因的RNA编辑与CMS之间存在着明显的联系。
刘海军等[14]比较分析了大豆线粒体基因atp6的编辑位点与CMS的关系, 研究结果表明不育系比保持系中多3个编辑位点, 其中2个位点导致了氨基酸类型的变化, 因此认为其可能与CMS有关。施真等[15]研究发现atp6基因转录本的RNA编辑导致的氨基酸变化与甜菜CMS有一定关系。韩利涛等[16]对大豆N8855型CMS不育系与其保持系的atp6基因的RNA编辑分析发现, 在不育系与保持系的atp6-3基因保守区中均有2个互不相同且编码不同氨基酸的编辑位点, 推测这可能与大豆的CMS有关; Jiang等[17]对大豆的atp9基因进行RNA编辑分析, 发现在cDNA水平上不育系和保持系之间发生了2次C-U转换, 导致二者编码蛋白不同; Mouras等[18, 19, 20]对烟草转入未编辑的atp9基因后, 获得转基因雄性不育烟草植株; 再对其转入反义atp9基因后, 转基因雄性不育烟草的育性得到恢复。Nakajima等[21]研究表明, 胡萝卜线粒体内的orfB基因结构与花瓣表型及花的CMS性存在一定的相关性, 并且它的表达是由转录后水平来调节的。Das等[22]对籼稻不育系及其同型保持系的orfB基因进行RELP检测, 结果表明在不育株系中的orfB基因具有一个更长的转录本。
烟草是我国乃至世界重要的经济作物和植物分子领域常用的模式植物[23, 24, 25]。我们此前从DNA[26, 27]和生物信息学[28, 29, 30]水平上部分报道过其F0-ATP亚基基因与雄性不育性的关系, 本文以3对烟草雄性不育系及其相应的保持系为材料, 研究其线粒体ATP合酶4个亚基基因F0部分atp6、atp9、orf25和orfB的RNA编辑位点, 通过分析烟草CMS不育系和保持系中这些基因在转录水平和基因组水平的差异, 以期进一步探讨它们与烟草CMS性的关系, 为研究烟草CMS分子机制奠定基础。
3对烟草细胞质雄性不育系及其同型保持系是MS中烟90 (MSzy90)和中烟90 (zy90); MS云烟85 (MSyy85)和云烟85 (yy85); MS K326 (MSK326)和K326。
选取烟草线粒体F0-ATP合酶4个亚基基因atp6、atp9、orf25和orfB进行PCR扩增和测序, 扩增引物是根据NCBI中GenBank数据库收录的烟草线粒体完全基因组序列(登录号为BA000042), 利用Primer Premier 5.0软件设计的(表1), 并由生工生物工程(上海)股份有限公司合成。
| 表1 用于CMS相关线粒体基因扩增的特异引物 Table 1 Special primers used to amplify mitochondrial genes related to CMS |
反转录合成cDNA第一链, 具体步骤如下: 取11 μ L总RNA于PCR管中, 加入1 μ L的Oligo dT Primer, 混匀后65℃水浴5 min, 迅速在冰上冷却5 min后加入5× 反应缓冲液4 μ L, 1 μ L RNase 抑制剂, 2 μ L dNTPs, 2 μ L逆转录酶AMV, 混匀后离心3~5 s, 42℃保温60 min后90℃保温10 min以终止反应, 冰上冷却几分钟后于-20℃保存。
PCR扩增DNA体系(25 μ L)含: 2× Taq PCR Master Mix 12.5 μ L、上下游引物各1 μ L (5 μ mol L-1)、模板1 μ L和ddH2O 9.5 μ L。PCR程序为94℃预变性3 min、94℃ 40 s、60℃ 30 s、72℃ 1 min、36个循环、72℃ 7 min。反应结束后4℃保存。PCR扩增cDNA体系(25 μ L)同上, PCR程序为94℃预变性3 min、94℃ 40 s、55℃ 30 s、72℃ 1 min、40个循环、72℃ 7 min。反应结束后4℃保存。
扩增产物经1%的琼脂糖凝胶电泳检测(每个样品重复3次), 并由生工生物工程(上海)有限公司测序。用DNAMAN和BlastN等软件对其核苷酸序列及编码的氨基酸序列比对分析。
比较DNA序列和cDNA序列的测序结果, 找出不育系及其保持系4个目的基因转录本发生的编辑位点, 比较分析不育系和保持系目的基因在编辑位点上的差异。记发生编辑的为“ +” , 未编辑的为“ -” [33]。
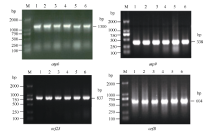
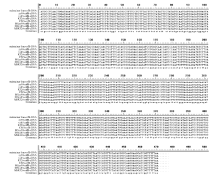
4个目的基因片段的PCR扩增产物在3种供试材料间不存在差异, 且与预期大小相吻合, 扩增效果较好(图1)。PCR扩增后的测序结果表明, 获得的atp6、atp9、orf25和orfB基因片段分别约为1300、338、837和604 bp, 且9个不育系与保持系样品的重复测序结果均各一致。序列比对后发现, 4个基因的保持系与GenBank中的基因序列相同; 但不育系较保持系和GenBank中的基因序列, 存在不同位点的碱基突变, atp6、atp9、orf25和orfB基因分别有6 (图3)、1 (图4)、2 (图5)和1 (图6)个该位点, 且在9个不育系样品中的碱基突变位点完全相同, 而大多数位点的碱基突变都导致该位点所编码氨基酸的变化。
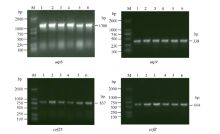
4个目的基因atp6、atp9、orf25、orfB的cDNA扩增产物在3种供试材料间不存在差异, 扩增效果较好(图2), 与4个目的基因的DNA扩增条带(图1)大小相近, 完全满足后续的测序要求。PCR扩增后的测序结果分别约为1300、338、837和604 bp, 且9个不育系与保持系样品的重复测序结果均各一致。说明扩增的片段即为4个目的基因cDNA。通过比较DNA序列和cDNA序列的测序结果发现, 4个目的基因都发生了不同程度的RNA编辑, 且编辑方式均为C→ U(T)的模式。
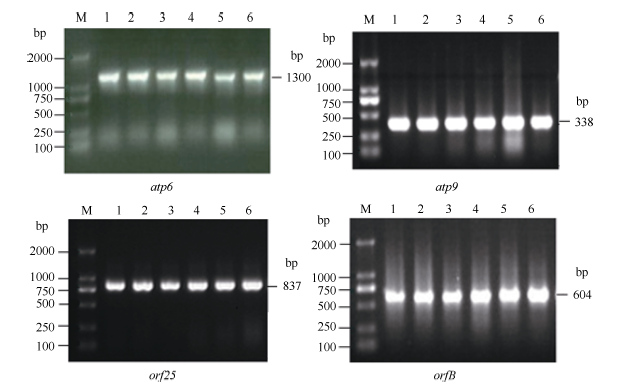 | 图1 烟草线粒体atp6、atp9、orf25、orfB基因PCR扩增结果Fig. 1 PCR amplification results of atp6, atp9, orf25, and orfB genes in tobacco mitochondrionM: 2000 DNA marker; 1: MSzy90; 2: zy90; 3: MSyy85; 4: yy85; 5: MSK326; 6: K326. |
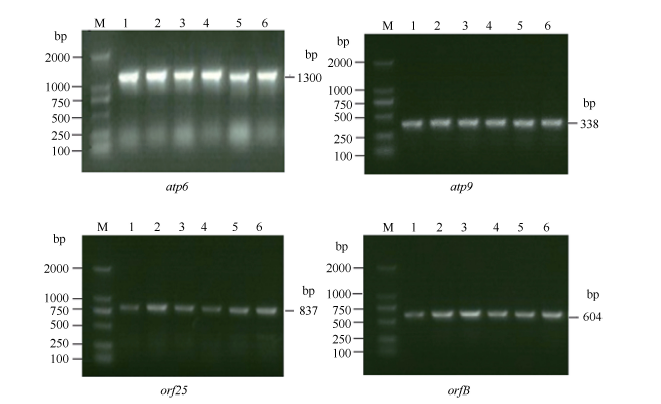 | 图2 烟草线粒体atp6、atp9、orf25、orfB基因cDNA序列PCR扩增结果Fig. 2 cDNA PCR amplification results of atp6, atp9, orf25, and orfB genes in tobacco mitochondrionM: 2000 DNA marker; 1: MSzy90; 2:zy90; 3: MSyy85; 4: yy85; 5: MSK326; 6: K326. |
2.2atp6基因的编辑位点分析
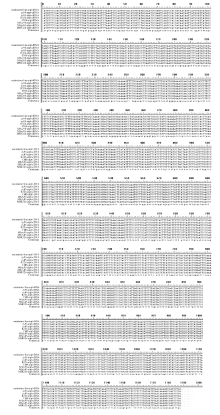
在3种烟草不育系材料中, atp6基因的转录本保守区都没有发生RNA编辑; 而在保持系中, atp6基因的转录本编辑位点共有6个, 编辑方式均为C→ U(T)的模式, 其中2个位点发生在密码子的第1位, 4个发生在密码子的第2位, 3个保持系材料发生的编辑位点均一致, 编辑后的基因在其氨基酸水平上均发生了改变, 使得不育系和保持系编码的蛋白完全不同, 这6个RNA编辑位点引起5种氨基酸类型的变化, 增强氨基酸疏水性的编辑位点有4个, 推测败育的形成可能与氨基酸疏水性的差异有关(图3)。
 | 图3 不育系及其保持系烟草中atp6基因的RNA编辑位点及基因突变位点466、545、602、658、683、1100位点为保持系atp6基因的RNA编辑位点; 59、92、253、418、768、1157位点为不育系atp6基因的基因突变位点。Fig. 3 RNA editing sites and gene mutation sites ofatp6 gene between cytoplasmic male sterility and maintainer lines in tobaccoRNA editing sites ofatp6 gene in maintainer lines were sites 466, 545, 602, 658, 683, 1100. Gene mutation sites ofatp6 in cytoplasmic male sterility were sites 59, 92, 253, 418, 768, 1157. |
2.3atp9基因的编辑位点分析
在不育系中, atp9基因转录本编辑位点共有8个, 且在3个不育系材料中编辑位点保持一致, 其中2个发生在密码子的第1位, 5个在密码子的第2位, 1个在密码子的第3位, 除第90个位点外, 其余位点均引起氨基酸类型的改变, 且有5个编辑位点增强了氨基酸的疏水性; atp9基因的保持系比不育系多2个编辑位点, 其中81位点的C→ T转换为缬氨酸的同义转换, 而191位点处发生脯氨酸(不育系)→ 亮氨酸(保持系)的转换, 推测这1个编辑位点对atp9 转录本的翻译及蛋白功能的正常发挥起重要作用。此外, 二者在第223个编辑位点上都产生了一个终止密码子, 缩短了转录本长度, 由于其在不育和保持系中都存在, 推测其可能是产生正常功能蛋白所必需的编辑(图4)。
 | 图4 不育系及其保持系烟草中atp9基因的RNA编辑位点及基因突变位点20、50、82、90、92、182、212、223位点为不育系atp9基因的RNA编辑位点; 20、50、81、82、90、92、182、191、212、223位点为保持系atp9基因的RNA编辑位点; 126位点为不育系atp9基因的基因突变位点。Fig. 4 RNA editing sites and gene mutation sites ofatp9 gene between cytoplasmic male sterility and maintainer lines in tobaccoRNA editing sites ofatp9 gene in cytoplasmic male sterility were sites 20, 50, 82, 90, 92, 182, 212, 223; RNA editing sites ofatp9 gene in maintainer lines were sites 20, 50, 81, 82, 90, 92, 182, 191, 212, 223; Gene mutation sites ofatp9 in cytoplasmic male sterility were site 126. |
2.4orf25基因的编辑位点分析
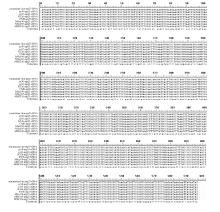
无论在保持系还是不育系中, orf25基因的转录本保守区中均存在10个RNA编辑位点, 编辑方式都属于C→ U(T)的模式, 且18个样品中发生的编辑位点一致, 编辑位点均发生在密码子的第2位, 且都导致了该位点所编码氨基酸类型的变化, 其中有8个氨基酸的改变均转换成亮氨酸, 有6个亲水氨基酸转换成疏水氨基酸。编辑结果提高了编码蛋白的疏水性, 同时也增加了编码蛋白在氨基酸序列上的保守性(图5)。
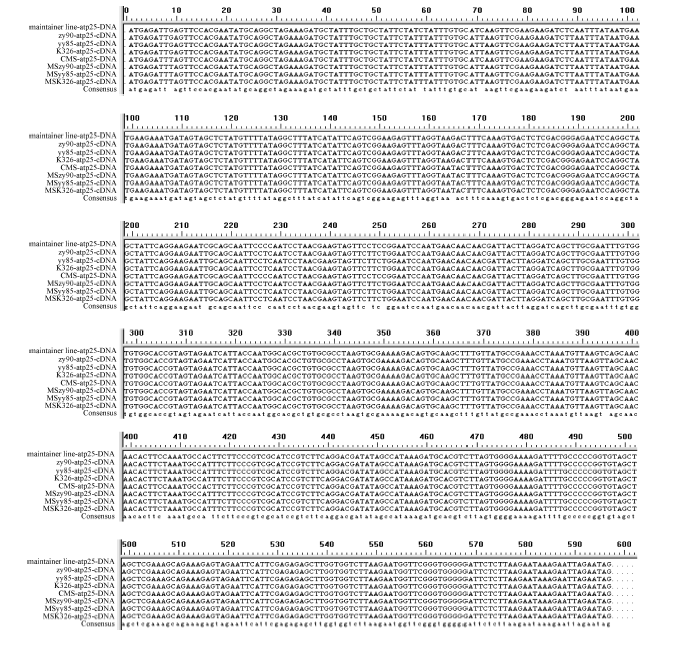 | 图5 不育系及其保持系烟草中orf25基因的RNA编辑位点及基因突变位点59、71、89、215、227、248、251、395、407、416位点为不育系和保持系orf25基因的RNA编辑位点; 9、165位点为不育系orf25基因的基因突变位点。Fig. 5 RNA editing sites and gene mutation sites oforf25 gene between cytoplasmic male sterility and maintainer lines in tobaccoRNA editing sites oforf25 gene between cytoplasmic male sterility and maintainer lines were sites 59, 71, 89, 215, 227, 248, 251, 395, 407, 416; Gene mutation sites oforf25 in cytoplasmic male sterility were sites 9, 165. |
2.5orfB基因的编辑位点分析
在保持系和不育系中, orfB基因的转录本均存在4个RNA编辑位点, 且18个样品中发生的编辑位点均一致。其中有2个位点发生在密码子的第1位, 另外2个位点发生在密码子的第2位, 这些编辑位点都会导致氨基酸种类的变化(图6)。
 | 图6 不育系及其保持系烟草中orfB基因的RNA编辑位点及基因突变位点47、58、76、443位点为不育系和保持系orfB基因的RNA编辑位点; 363为不育系orfB基因的基因突变位点。Fig. 6 RNA editing sites and gene mutation sites oforfB gene between cytoplasmic male sterility and maintainer lines in tobaccoRNA editing sites oforfB gene between cytoplasmic male sterility and maintainer lines were sites 47, 58, 76, 443; Gene mutation sites oforfB in cytoplasmic male sterility were site 363. |
RNA编辑作为高等植物线粒体基因组转录后水平基因表达调控的一种重要方式, 在植物的生长发育过程中发挥着重要的作用[34]。它的研究使人们认识到, mRNA中的序列并不一定都是从DNA直接转录而来的, 对于有些蛋白质, 仅仅从其DNA序列来推断蛋白质序列已成为历史。RNA编辑位点的发现和鉴定是研究其生物学功能的前提, 也为深入研究其分子机制、揭示其作用机理奠定了基础。本研究将3对烟草雄性不育系及其相应保持系4个线粒体基因的DNA与cDNA碱基序列对比分析发现, 不育系和保持系中4个目的基因的RNA编辑方式均为C→ U(T)的转换, 研究表明该方式是通过基因组编码的C残基脱氨基来完成的[35]。在4个目的基因中, RNA编辑位点大多数都发生在密码子第1和第2位点, 所造成的核苷酸的改变均会改变编码氨基酸的种类。发生在密码子第3位点的编辑并未引起氨基酸类型的改变, 从总体上看, 这些RNA编辑都增加了蛋白质疏水性, 同时也增强了物种间的保守性。这与前人的研究结果是一致的[36]。从4个目的基因发生编辑的总体情况来看, 与保持系相比, 不育系中atp6基因的编辑位点完全缺失, 保持系比不育系多了6个RNA编辑位点, 从而认为未编辑的atp6基因可能是CMS的关键因子。研究表明未编辑的atp9基因的表达与烟草CMS的形成有关[18, 19, 20]。本研究结果发现atp9基因的保持系比不育系多2个编辑位点, 其中81位点的C→ T转换为缬氨酸的同义转换, 而191位点处发生脯氨酸→ 亮氨酸的转换极有可能与整个atp9亚基的结构和功能有关, 因此烟草有可能正是由于缺少这一特异转录本编辑位点而影响atp9基因功能的正常发挥。易平等[37]发现红莲(HL)型水稻线粒体atp6基因发生的18个编辑位点在不育系和保持系中无差异, 但不育系的编辑频率低于保持系, 二者相差不显著。而orf25和orfB基因的编辑位点在保持系和不育系中无变化, 为完全编辑, 但是否在编辑频率上存在差异而影响ATP合酶的正常功能还有待进一步研究。因此, RNA的正常编辑对线粒体功能的正常发挥起着重要作用, 编辑不充分的转录产物最终会干扰线粒体功能的正常发挥[37]。
本试验结果与一些报道相吻合, 但与一些研究结果存在差异。在保持系中, atp9基因的特异编辑位点与NCBI中收录的和Hernould[18]报道的完全一致, orf25基因发生的RNA编辑情况也与NCBI中所收录的相吻合; 而atp6和orfB基因编辑位点比NCBI中收录的缺少几个位点, 推测这可能与试验所用材料品种或所取材料部位的差异有关。本试验所用材料为与育性直接相关的花蕾, 而NCBI收录的数据所用实验材料大多数为叶片, 从而表明RNA编辑能力还可能受发育时期和组织特异性的调控[33]。此外, 也可能与本实验所采取的PCR产物直接测序法有关, 该测序方法要求编辑位点是完全编辑或编辑频率达到一定的值[38], 因此可能丢失一些低频率的编辑位点。
此前报道过烟草中未编辑的atp9基因的表达与CMS的形成有关。但目前还未曾见过有关未编辑的atp6、orf25和orfB基因的表达与烟草雄性不育之间关系的报道。从本试验结果及之前对于其他作物的相关报道来看, 对于atp6基因, 保持系比不育系多6个RNA编辑位点, 从而推测未编辑的atp6基因可能是CMS 的关键因子。另外, 虽然orf25和orfB基因在不育系和保持系上的编辑位点完全一致, 但可能编辑频率存在一定的差异, 从而影响ATP合酶功能的正常发挥。推测烟草雄性不育系这4个亚基基因中的一个或者几个或者全部的特异RNA编辑很有可能就是导致烟草CMS的重要原因, 但编辑位点改变是如何影响基因功能的, 还需要进一步深入的研究。下一步我们将利用生物信息学方法比较ATP合酶F0部分的4个亚基基因推导蛋白在不育系和保持系中各结构层次(一级、二级、超二级和三级结构)上的差异, 构建蛋白质互作模型, 预测F0亚基互作特征, 构建F0亚基互作网络, 分析蛋白质水平上联合效应与烟草雄性不育性之间的调控机制。
orf25和orfB基因转录本在不育系和保持系中发生的编辑位点是一致的, atp6基因在不育系中未发生编辑, 在保持系中共有6个RNA编辑位点, atp9基因保持系比不育系多2个C→ T的特异编辑位点, 推测烟草不育胞质可能因缺少特异的线粒体基因转录本编辑而导致细胞质雄性不育。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|