
{kind=link}
{kind=link}
大豆不同环境下脂肪酸组分含量的QTL分析
[雷雅坤1, 2, ** , 刘兵强1, * , 邸锐1 , 闫龙1 , 杨春燕1 , 郝东旭3 , 张孟臣1, *  ]
]
]
|
|


第一作者联系方式: E-mail: yakun0628@163.com **同等贡献(Contributed equally to this work)
大豆油的品质取决于脂肪酸各组分在大豆中的比例, 为发掘控制大豆5种脂肪酸含量的数量性状位点(QTL), 利用冀豆12和黑豆重组自交系群体构建遗传图谱, 采用Windows QTL Cartographer 2.5和QTL Network-2.0软件的CIM和MCIM法对大豆5种脂肪酸组分进行数量性状定位。结果表明, 在石家庄和三亚各环境下共检测到16个QTL, 位于连锁群A2、B2、C2、F、G、I、L上。对2个环境联合分析, 检测到13个QTL, 其中9个用2种方法被检测到, 但这13个位点与环境互作的贡献率明显小于加性效应。其中在B2连锁群Satt168~Satt556控制硬脂酸的QTL Ste-1在河北石家庄和海南三亚均能被检测到, 贡献率均为12%, 在双尾群体和间隔挑选群体中也能检测到控制硬脂酸的QTL Ste-1, 说明这一QTL稳定存在于本组合群体中, 为今后大豆硬脂酸的QTL精细定位奠定了基础。
Soybean is an important crop which contributes about 60% of the world’s oilseed production. The quality of soybean oil depends on the relative composition of fatty acid in seeds. A population of recombinant inbred lines derived from a cross of Jidou 12 × Heidou was grown in two environments and the seed samples from the environments were evaluated for fatty acid contents. Simple sequence repeat (SSR) markers were used to construct genetic linkage map. A total of 16 QTLs underlying fatty acid contents were identified by CIM and MCIM methods using Windows QTL Cartographer 2.5 and QTL Network-2.0 software at Sanya and Shijiazhuang locations, respectively. These QTLs were scattered on linkage groups A2, B2, C2, F, G, I, L. According to the two environment combined data, 13 QTLs were detected using two mapping methods. Nine of them were common in the results from two methods. Another QTL associated with stearic acid content, named as Ste-1, located on LG B2 and flanked by Satt168 and Satt556, was stable across two locations, also. QTL Ste-1 could explain 12% of the phenotypic variation at both locations. This study is helpful to improve fatty acid composition in soybean.
大豆油是主要的食用油之一, 大豆的蛋白、矿物质和维生素含量丰富, 其不饱和脂肪酸含量在油分中也具有很高的比例。因此, 应重视大豆脂肪酸各组分的比例, 以便开展大豆品质育种, 提高脂肪含量[1]。大豆棕榈酸、硬脂酸、油酸、亚油酸与亚麻酸5种脂肪酸含量都属于数量性状[2]。Nickell和Brummer[3]都用RFLP标记分别在连锁群D和B2上定位到了控制棕榈酸和亚麻酸的位点。1992年Diers等[4]检测到影响棕榈酸、油酸和亚油酸的3个QTL, 解释10%以上的遗传变异。2002年Li等[5]通过Cook× N87-2122-4构建的重组自交系F2和F2:3群体, 在连锁群上检测到了控制棕榈酸的2个QTL, 分布在连锁群A1标记Satt684附近和M连锁群标记Satt175附近, F2群体在这2个位置上可解释的遗传变异为38%和8%, F2:3群体在这2个位置上可解释的遗传变异分别为33%和9%; 所检测到的2个连锁群上的区域的主效基因和微效基因互作效应显著。2003年Spencer等[6]通过群体检测到影响硬脂酸的QTL, 位于B2连锁群上标记Satt070、Satt474和Satt556附近。2006年郑永战等[7]用Essex× ZDD2315的回交群体F1的114个单株, 检测到控制5种脂肪酸含量的16个位点, 分布于B2、C1、D1b、D2、E、H、I、L、N等9个连锁群上, 可解释的遗传变异为8.2%~39.3%, 其中在B2上检测到的控制硬脂酸的位点和Spencer检测到的位点一致。在不同群体中所定位到的控制脂肪酸含量的QTL有所差异, 通过对所报道的位点进行整合, 可以从整体上分析大豆脂肪酸的QTL, 选出比较可靠的位点用于育种工作。1996年Orf等[8]对报道过的14个群体进行整理, 在5个群体里控制大豆脂肪酸的QTL有12个遗传贡献率超过了10%。2009年宋万坤等[9]对大豆脂肪酸组分QTL进行了元分析, 影响大豆5种脂肪酸组分的QTL主要分布在A1、B2、D1b、D2、E、G、L这7条连锁群上, 且成簇分布。其中影响大豆棕榈酸、硬脂酸、油酸、亚油酸、亚麻酸含量的QTL分别为3、1、5、2和1个, 还有1个QTL同时影响大豆的棕榈酸、硬脂酸和油酸含量。于福宽[10]利用鲁黑豆2号和南汇早黑豆重组自交系群体对棕榈酸、硬脂酸、油酸、亚油酸、亚麻酸、油分等性状检测到13个QTL。分布于A1、M、G、O、K、B2和N连锁群上, LOD值为2.62~12.70, 遗传贡献率为5.60%~ 21.39%。
目前, 研究者们比较了不同环境中以同一群体定位到的QTL。朱军[11]提出的基于混合线性模型复合区间作图法为分析QTL与环境的互作效应提供了有力手段, 可以直接分析出QTL与环境互作的遗传贡献率。单大鹏等[12]用Charleston× 东农594的RIL群体连续5年对油份进行QTL定位, 检测到11个位点, 其中有2个QTL没有与环境的互作。苗兴芬[13]用Charleston× 东农594的群体通过3个年份3个地点的试验, 检测到5种脂肪酸的68个位点, 其中有19个存在与环境的互作。
本试验利用冀豆12和黑豆组合的F6群体构建更完整的大豆遗传连锁图谱, 连续2年对5种脂肪酸组分含量进行QTL定位, 在三亚环境下对石家庄定位到的数量性状位点进行验证, 找到更加可靠的控制5种脂肪酸组分含量的QTL。对在石家庄和三亚检测到的脂肪酸含量进行了上位效应和QTL与环境互作效应的分析, 有利于得到受环境影响较小, 能稳定遗传的QTL。
以5种脂肪酸含量均有差异的冀豆12为母本, 黑豆为父本进行杂交。通过单粒传法得到本群体F6的211个家系。2010年冬将收获的F6单株种植于海南三亚实验站, 每个家系1行, 行长2 m, 株行混收, 检测了211个家系脂肪酸主要组分含量。2011年夏, 将收获的F7种植于河北石家庄堤上实验站, 对F8进行脂肪酸主要组分含量的测定。根据F7群体硬脂酸含量的表型数据, 从211个家系中选取含量最高和最低的各30株建立双尾群体; 每隔3株挑选1株建立间隔挑选群体。
以本群体211个家系F6单株的新鲜嫩叶为试验材料, 用SDS法提取大豆基因组DNA, PCR体系含5 μ L DNA、2.0 μ L 10× PCR buffer、2.5 mmol L-1 dNTPs、1.5 μ mol L-1引物、1 U Taq酶, 加ddH2O补足至20 μ L。反应程序为94℃预变性5 min; 94℃变性30 s, 47℃退火30 s, 72℃延伸30 s, 35个循环; 72℃延伸5 min, 4℃保存。
采用安捷伦气相色谱6890N, 将供试材料用研磨仪充分研磨, 磨好后取0.04 g豆粉放入2.0 mL离心管中, 加1 mL脂肪提取液, 振荡混匀10 min, 10 625× g离心后取上清液, 加0.5 mol L-1甲醇钠甲酯化1 mL, 静置10 min, 加饱和NaCl溶液至样品分层。吸取上清液进行气相色谱分析。选用FID检测器, FFAP弹性石英毛细管柱, 恒压的模式, 载气为高纯氮气, 流速为1 mL min-1; H2气流速为35 mL min-1; 空气流速为350 mL min-1。进样量为1 μ L。分流比为10∶ 1, 进样口的温度为250℃。检测器的温度为250℃。程序温为180℃ 3 min, 9℃ min-1升到225℃ min-1, 保持25 min[14]。
利用Map Manager QTXb20 (P=0.0001, 函数Kosambi)软件对冀豆12× 黑豆群体进行遗传连锁图谱的构建和分析。用Windows QTL Cartographer 2.5软件及复合区间作图法, 取LOD=2.5为阈值, Walk Speed=1.5 cM, 对由211个家系组成的大群体、双尾群体和间隔挑选群体的5种脂肪酸含量进行QTL检测, 同时分析各位点的遗传贡献率和加性效应。用QTL Network-2.0软件及基于混合线性模型的复合区间作图法, 对2个环境下的5种脂肪酸含量进行联合分析, 并分析所检测到位点的加性效应和与环境的互作效应。
从表1可看出, 在不同环境中2个亲本的5种脂肪酸组分含量有一定的变化。后代群体的5种脂肪酸含量中除三亚环境的油酸最小值为14.86%, 比低亲的父本黑豆高0.81个百分点外, 其余性状都具有双向超亲的现象, 且群体后代的平均值均位于两亲本之间, 在2个环境中硬脂酸的平均值分别为3.36%和3.15%, 均倾向于低亲值, 油酸的平均值分别为17.89%和17.19%, 均倾向于高亲值。在5种脂肪酸组分中, 硬脂酸的变异幅度较小, 油酸和亚油酸的变异幅度较大, 表明在这2个性状上后代群体有一定差异, 对提高油酸和亚油酸在大豆5种脂肪酸组分中所占的比例有一定意义。5种脂肪酸组分的最大值和最小值差异较明显, 说明在后代分离群体中的表现分离程度较大, 且变异连续, 在2个环境中5种脂肪酸组分偏度的绝对值都小于1, 基本呈正态分布, 是属于多基因控制的数量性状。
| 表1 亲本及后代群体脂肪酸性状统计结果 Table 1 Statistics results of fatty acids traits of parents and descendant population |
表2表明, F7棕榈酸和油酸、亚油酸呈极显著负相关, 硬脂酸和亚麻酸呈显著负相关, 油酸和亚油酸、亚麻酸呈极显著负相关, F7脂肪酸间的相关除了硬脂酸和亚麻酸的相关程度外, 其余均和F8脂肪酸间的相关性一致。棕榈酸、硬脂酸、油酸、亚油酸和亚麻酸F7和F8之间都呈极显著正相关。说明F7和F8的脂肪酸组分含量受环境因素的影响较大。
| 表2 群体F7和F8两代间脂肪酸的相关 Table 2 Correlatin coefficient of five types of fatty acids between F7 and F8 populations |
参照Cregan等[15]公布的大豆遗传连锁图谱, 在20条连锁群上, 约每10 cM挑选2~3对引物, 共筛选了495对引物, 其中有多态的为221对, 从中选取130对分布均匀的引物, 利用构建遗传连锁图谱, 能得到条带清晰的为118对, 与冀豆12带型一致的记为A, 与黑豆带型一致的记为B, 杂合的带型记为H, 缺失的记为-。
利用SSR标记对冀豆12× 黑豆群体构建大豆遗传连锁图谱, 共包括21条连锁群。与大豆公共图谱相比较, 连锁群与公共图谱有很好的对应, 标记顺序与公共图谱完全一致。遗传连锁图谱的标记为117个SSR标记, 每个连锁群上平均包括6个标记, 图谱总长度为1501.0 cM, 标记间平均距离为15.6 cM[16]。
采用复合区间作图法分析石家庄和三亚2个环境大豆5种脂肪酸含量的数据, 共检测到16个QTL (图1和表3)。
在石家庄环境可以检测到4个控制硬脂酸的QTL, 分别位于连锁群B2、C2和I, 在三亚环境还可以检测到2个QTL, 分别位于连锁群B2和C2 (表3), LOD值为2.50~ 7.03, 遗传贡献率为11%~14%。其中QTL Ste-1在2个环境中都被检测到, 增效基因均来自母本冀豆12, 且可解释的遗传变异都为12%。
 | 图1 冀豆12× 黑豆中检测出的5种脂肪酸含量QTL在连锁群的分布Fig. 1 Distrbution of QTL for five types of fatty acids detected from Jidou 12× Heidou on linkage map |
| 表3 2年中用CIM法定位的5种脂肪酸含量QTL Table 3 QTLs mapping of five fatty acid contents with CIM method in two years |
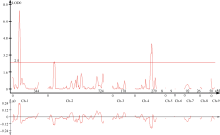
在双尾群体和间隔挑选群体中, 均可定位到控制硬脂酸的QTL Ste-1, 遗传贡献率为15%~27%, 在双尾群体中定位到位于B2和C2上的QTL与石家庄环境中所检测到的位点一致(图2), 其中位于B2上的QTL Ste-3在间隔挑选群体中也被检测到, 说明分布在连锁群B2和C2上的控制硬脂酸的QTL比较稳定。在双尾群体I连锁群标记Satt148附近和在间隔挑选群体H连锁群标记Satt541- Sat-118检测到了控制硬脂酸的位点, 这2个QTL与冀豆12× 黑豆群体定位到的位点不同, 有待进一步验证。
 | 图2 复合区间作图法对硬脂酸的QTL定位似然比图谱Fig. 2 Likelihood ratio maps of QTL tagging for stearic acid using CIM |
在石家庄环境检测到1个棕榈酸含量QTL (QTL Pal-1), 位于F连锁群, LOD值为2.69, 可解释6%的遗传变异。在石家庄环境检测到2个油酸含量QTL, 都分布在I连锁群的Satt162附近, 遗传贡献率分别为9%和10%, 其增效基因均来自母本冀豆12, 2个QTL距离较近, 在连锁群I上可能有控制油酸的主效QTL存在, 在海南环境检测到1个QTL, 位于连锁群L。在石家庄环境下只检测到2个亚油酸含量QTL, 分别位于连锁群C2和I, 可解释的遗传变异分别为12%和6%。在石家庄环境检测到3个控制亚麻酸的QTL, 分别位于连锁群A2、C2和G上, LOD值为2.44~5.07, 遗传贡献率为7%~15%。其中位于连锁群G标记Satt612-Sat_372之间的QTL Len-5, 在石家庄环境本群体的F4中也曾被检测到, 其增效基因来自父本黑豆。在三亚环境检测到2个控制亚麻酸的QTL, 分别位于连锁群C2和I, 可解释的遗传变异为5%和8%。
利用F6群体的分子数据及石家庄和三亚2个环境下5种脂肪酸含量的数据, 用QTLNetwork-2.1软件基于混合线性模型的复合区间作图法对2个环境的大豆脂肪酸组分含量进行联合分析(表4), 共检测到13个QTL, 可解释的遗传变异范围为3.8%~15.4%, 和CIM法相比, 2种方法都能检测到的QTL有9个, 但贡献率较CIM法有所下降。
13个QTL中有12个与环境具互作效应, 其效应值为0.0001~0.4458, 2个环境对单个位点的效应值大致相等, 但作用相反。位点与环境的互作均没有达到显著水平。其中分布在I连锁群标记Satt049~Sat_418控制亚油酸的QTL与环境互作效应的贡献率为0, 说明该位点与环境没有发生互作效应。控制亚油酸的QTLN Lei-2与环境互作的遗传贡献率最大, 为0.89%。13个QTL与环境互作效应总的遗传贡献率为3.04%, 明显小于自身加性效应的遗传贡献率。说明本群体所定位的QTL受环境影响较小, 比较稳定、可靠。从表4可知, 除了3个控制油酸位点的加性效应都为正值, 增效基因均来自父本外, 控制其余4种脂肪酸位点的加性效应同时存在正值和负值, 这也说明了5种脂肪酸组分除F7三亚环境的油酸没有负向超亲外其余均有双向超亲分布, 且最大值和最小值之间差异显著。
| 表4 2年中用MCIM法定位的5种脂肪酸含量QTL及其与环境的互作 Table 4 QTLs mapping of five fatty acid contents QE interaction with MCIM in two years |
由于大豆脂肪酸组分含量属于数量性状, 环境和年份对其影响较大, 从2年的脂肪酸组分含量定位结果可知, 控制5种脂肪酸组分含量的QTL较集中分布在B2、C2和I连锁群。其中, 在B2连锁群标记Satt168-Satt556控制硬脂酸的QTL在2个环境中可被重复检测到, 且2年的遗传贡献率都为12%。这一位点和Spencer等[6]利用普通硬脂酸含量品种Dare和高硬脂酸含量品种FAM94-41的重组自交系F2、F2:3报道过的位点一致, 郑永战等[7]也曾用高含油量品种Essex和低含油量品种ZDD2315的回交群体报道过这个位点。在不同年份地点、环境下得到的相同的基因位点, 是硬脂酸的重要基因位点。在石家庄环境B2连锁群上还可以检测到一个控制硬脂酸的QTL位点, 和2006年郑永战等[7]利用回交群体报道的位点一致。本试验定位的区间和曾报道的相比, 遗传距离在公共图谱上缩短了1.13 cM。三亚环境下检测到的在L连锁群上控制油酸的位点和Maria等[17]在2008年用G99~G725和N00~3350的F2:3定位到的位点都与标记Satt462相连锁。在C2连锁群标记Satt286-Satt460控制硬脂酸的位点和在G连锁群上控制亚麻酸的位点曾在本群体F2:4中被检测到[18], 且这一位点和邹筱[19]报道的与脂肪、硬脂酸、油酸、亚油酸含量相关的Satt079标记的遗传距离较近。由于数量性状受环境和群体的影响, 所以, 在不同群体和同一群体不同环境中重复检测到的位点比较可靠, 挖掘这些有利基因有着重要的意义。
本试验检测到控制5种脂肪酸组分含量的QTL位点中, 在不同年份所检测到的控制同一性状的位点分布在同一个连锁群不同标记之间且遗传距离很近, 如在C2连锁群上控制硬脂酸的QTL, 在2008年三亚环境下和2009年石家庄环境下被定位在标记Satt316-Satt307, 在2010年三亚环境下被定位在标记Satt643-Satt286, 在2011年石家庄环境下被定位在标记Satt286-Satt460, 遗传贡献率分别为7%, 11%, 14%。在G连锁群上控制亚麻酸的QTL位点, 在2008年石家庄环境下被定位在标记Satt517- Satt612, 在2011年石家庄环境下被定位在标记Satt612- Sat_372, 遗传贡献率分别为20%和15%。
以上几对分布在连锁群C2和G上的QTL在公共图谱中的遗传距离很近, 由于环境对定位造成的影响, 这几个QTL可能是一个, 但在QTL的初定位中没有明确的结论, 是否为同一个QTL只能通过这一区域引物的加密, 在不同环境反复试验继续验证。Fulton等[20]认为, 在多种环境中能同时检测到的QTL可能比那些只能在一种环境中检测到且效应值更高的QTL更有利用价值, 这样的QTL在转移到新的遗传背景或在不同的环境条件下评价时更稳定。同时分析多个环境下的数据, 能增大QTL的检测强度, 准确估计QTL的位置和效应[21]。对于遗传距离较近的QTL, 单大鹏等[10]将其归为基本一致的QTL, 但对可以归为一个QTL的遗传距离没有做出限制, 对于本试验中在硬脂酸和亚麻酸上检测到的这2个标记区域可以作进一步研究, 以利于说明这个问题。
在数量性状基因定位时经常将控制不同性状的基因定位到连锁群的同一区域, 本研究发现, 控制亚麻酸和与其相关的油酸、亚油酸3个性状的QTL有部分重叠现象(表3和图1)。在I连锁群标记Satt162-Satt148同时控制油酸和亚麻酸的含量, 在I连锁群标记Sat_418附近检测到控制油酸和亚油酸这2个性状的QTL。由表2可知, 在F8中, 油酸和亚油酸、亚麻酸都呈极显著负相关, 与于福宽[10]和邹筱等[19]报道的结果一致, 说明这一相关在本群体F7和不同群体中都稳定存在[22, 23], 试验结果表明, 在连锁群I上很可能存在控制这一相关的基因位点。在F8中, 亚油酸和亚麻酸存在极显著负相关, 这一相关徐豹等[24]曾报道过, C2连锁群Satt457-Satt322的QTL同时控制亚油酸和亚麻酸的含量, 说明2个性状间重叠的QTL与相关性具有一致性。这些位点从分子层面上对油酸、亚油酸和亚麻酸的相关性做出了更好的解释, 也证明了基因一因多效的现象。通过脂肪酸组分间的相关, 对其中一种组分的选择可以影响到遗传距离较近的另一种脂肪酸组分。本研究中具有多效性脂肪酸QTL集中在C2和I连锁群上。同时控制多个数量性状的可能是同一个基因位点, 也可能是多个基因紧密连锁共同对一个性状起作用[25]。目前的QTL定位中精细定位的研究还很少, 随着分子技术的发展尤其是SNP的运用, 遗传连锁图谱的不断加密和完善, 可以把QTL准确定位到大豆遗传连锁图谱中, 把在同一个区域中定位到的距离较近的QTL分解开, 可以通过精细定位和图位克隆技术, 最终将有价值的基因应用到分子育种上[26]。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| 20 |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|