


{kind=link}
{kind=link}
{kind=link}
{kind=link}
甘蓝型油菜结角高度与荚层厚度的全基因组关联分析
[卢坤1, *  , 王腾岳
, 王腾岳1, * , 徐新福1 , 唐章林1 , 曲存民1 , 贺斌2 , 梁颖1 , 李加纳1, * ]
, 王腾岳, 徐新福]
|
|


第一作者联系方式: 卢坤, E-mail: drlukun@swu.edu.cn; 王腾岳, E-mail: 18306077869@163.com **同等贡献(Contributed equally to this work)
角果是油菜重要的光合作用和种子存储器官, 对油菜产量具有重要贡献。本研究以412份具有代表性的甘蓝型油菜品种(系)为材料, 利用芸薹属60K Illumina Infinium SNP芯片对其基因型分析, 并对油菜结角高度和角果层厚度进行全基因组关联分析。结果共检测到16个显著关联的SNP, 其中重庆环境下分别检测到2个和4个SNP与结角高度和结角层厚度显著关联, 单个SNP解释的表型变异为5.61%~5.69%和5.94%~6.31%。云南环境下分别检测到5个和1个显著关联的SNP, 单个标记解释的表型变异为12.66%~13.97%和22.43%。对2个环境的结角高度差和结角层厚度差共检测到3个和1个与性状显著相关的SNP, 它们对表型变异的解释率分别为17.33%~20.32%和29.05%。其中, 环境间结角厚度差的关联SNP与重庆环境结角层厚度的1个显著关联SNP位于同一LD区间。各显著关联标记LD区段的多个基因调节植物细胞组织发生、花分生组织发育、角果数目和多器官发育, 如 NSN1、 TPST和 SAC1等, 它们可能通过上述功能影响油菜花序或角果的生长发育, 导致结角高度或结角层厚度差异。本研究发掘的这些位点和候选基因可作为影响油菜结角高度和角果层厚度的重要候选区域和基因, 为揭示油菜结角性状的遗传基础和分子机制, 提高油菜单位面积产量奠定了基础。
, WANG Teng-Yue, XU Xin-Fu
Layer of pod canopy is an important photosynthetic and seed storage part in rapeseed, providing important contribution to yield. In this study, 412 representative Brassica napus varieties (or lines) were genotyped using the Brassica 60 K Illumina Infinium SNP array by genome-wide association analysis of the height of podding (HP) and thickness of pod canopy (TPC). A total of 16 significant SNPs were identified, including two and four SNPs associated with HP and TPC in Chongqing, each of them explained 5.61-5.69% and 5.94-6.31% of phenotypic variation, respectively. Five and one significant SNPs accounting for 12.66-13.97% and 22.43% of the phenotypic variation for HP and TPC in Yunnan, respectively, were also detected. Three and one significant SNPs associated with the difference of HP and TPC between two environments were detected, explaining 17.33-20.32% and 29.05% of phenotypic variation, respectively. The latter SNP marker was located in the same linkage disequilibrium (LD) interval with one of significant SNPs related to TPC in Chongqing. Functional annotation of genes within the LD intervals containing significant markers showed that several genes involved in regulation of cell organization and biogenesis, floral meristem development, number of silique, and multicellular organismal development existed, such as NSN1, TPST, and SAC1, which might result in the variation of HP and TPC through affecting the growth and development of flower or silique in B. napus. These loci and genes could be regarded as important candidate regions and genes for HP and TPC of B. napus. The results lay the foundation for revealing the genetic basis and molecular mechanism for podding traits, and improving the yield per unit area of B. napus.
甘蓝型油菜(Brassica napus)是全球四大油料作物之一, 也是我国主要的油料、饲料和能源作物, 常年播种面积和总产均居世界首位[1]。作为我国传统食用油, 菜籽油已占国产食用植物油55%以上, 成为国产食用植物油第一大来源[2]。中国种植业信息网农作物数据库(http://www.zzys.moa.gov.cn/)统计数据表明, 近年国内食用植物油需求急剧上升, 国产食用油自给率降低至26%左右, 是当前我国大宗农产品中对国际市场依存度最大, 安全形势最严峻的作物。作为冬季油料作物, 油菜不与粮争地, 如果能够利用我国长江流域近700万公顷冬闲田发展油菜生产, 将有效缓解我国植物油供需矛盾, 保障我国食用植物油供给安全[2]。
作物的产量性状是极其复杂的数量性状, 是作物整个生命周期内一系列生长发育过程和环境互作的最终产物, 通常由多基因控制。对油菜来说, 产量由4个主要产量构成因子决定, 包括种植密度、单株角果数、每角粒数和千粒重[3, 4, 5]。一些与产量相关的株型和角果性状, 如株高、角果长度、角果密度、分枝数、分枝起点高度、分枝角果数和结角层厚度等也会通过影响产量构成因子或其他未知机制影响油菜产量[6, 7]。各产量构成因子和产量相关性状间存在复杂的互作关系, 它们决定了油菜产量潜力, 如千粒重、每角粒数、主序有效角果数、株高、角果层长度、角果层宽度均与单株产量呈极显著正相关, 但千粒重与单株有效角果数和每角粒数均为显著负相关[8, 9], 单株有效角果数与每角粒数也呈极显著负相关[10]。前人利用DNA分子标记技术和QTL (quantitative trait loci)作图方法对油菜单株角果数、每角果粒数和千粒重、株高、分枝起点等性状开展了大量QTL定位研究, 鉴定出多个控制性状变异的主效QTL[4, 7, 11, 12, 13]。但目前的研究主要集中在油菜产量构成因子的QTL定位中, 对密切影响产量和机械收获效率的结角高度(最低有效角果的高度)和结角层厚度的定位研究还非常少。全基因组关联分析(genome-wide association study, GWAS)是检测全基因组范围的遗传变异, 从而寻找与复杂性状相关的遗传因素的研究策略, 目前已在人类重大疾病研究中发挥了重要作用, 对植物复杂数量性状的遗传研究也起到了积极的推动作用。与传统QTL定位方法相比, GWAS采用遗传背景丰富的自然群体为材料, 不需要花多年时间构建特定的分离群体, 具有作图定位精度高、能同时扫描控制目标性状所有关联位点的优点。随着甘蓝型油菜全基因组序列的公布[15]及芸薹属60 K SNP (single nucleotide polymorphism)芯片的开发, GWAS在油菜重要数量性状的遗传研究中也发挥着重要的作用, 鉴定出控制种子重量、单株产量、含油量、株高、一次分枝数、种子萌发和活力相关性状的位点及候选基因[15, 16, 17, 18]。为了研究控制油菜结角高度和角果层厚度的QTL, 本研究采用412份甘蓝型油菜自交系构建自然群体, 结合SNP芯片数据及云南临沧和重庆北碚两地的表型数据, 在最优模型下以GWAS鉴定与目标性状显著关联的SNP, 以连锁不平衡(linkage disequilibrium, LD)分析QTL的LD区间。最后, 根据该区段甘蓝型油菜基因组序列进行基因功能注释, 初步确定可能控制目标性状变异的候选基因, 为通过分子标记辅助选择等方式提高甘蓝型油菜单位面积产量提供理论依据。
关联分析材料包括国内外油菜主产区的412份具有代表性的油菜种质资源, 其中, 国内材料381份, 主要来自重庆、湖北、湖南、陕西、江苏等地, 国外材料31份, 主要来自德国、加拿大、丹麦、瑞典等国。所有材料均由重庆市油菜工程技术研究中心提供。
2013年9月至2014年5月, 将412份材料中的392份种植于重庆市油菜工程技术研究中心歇马实验种植基地(北纬29º 45'39.99", 东经106º 22'38.47", 海拔238.57 m), 根据前期研究结果, 选择118份与重庆种植生育期一致的材料(与重庆共有材料98份)种植于云南省临沧市农业技术推广站临翔区博尚镇勐准村实验种植基地(北纬23º 43'56.69"', 东经100º 02'4.79", 海拔1819.50 m)。田间试验按3个重复随机区组设计, 每个小区种植2行, 行距40 cm, 株距20 cm。田间管理同常规生产, 确保同一地区所有样本的生长环境一致。待油菜成熟收获时, 从各小区随机选取5株长势一致的株系, 测定各株系的结角高度(height of podding, HP)和结角层厚度(thickness of pod canopy, TPC)。结角高度为子叶节与下部第1个有效角果着生部位的垂直高度差; 结角层厚度为下部第1个有效角果着生的部位与上部最顶端有效角果着生部位的垂直高度差, 以厘米(cm)为单位。为了定位上述性状受环境调控的位点, 同时计算了两地共有材料的结角高度差和结角层厚度差。采用Microsoft Excel 2007软件初步整理表型数据, 后采用SPSS 20软件统计分析。
利用芸薹属60K Illumina Infinium SNP芯片, 严格按照试验操作说明对412份甘蓝型油菜群体材料进行SNP基因分型[18]。采用GenomeStudio (Illumina公司)软件分析SNP基因型, 排除最小基因型频率(minor allele frequency, MAF)低于0.05和SNP得率(call frequency)小于80%的标记, 最终获得31 832个在油菜基因组上具有唯一位置的高质量SNP标记用于后续分析。
根据SNP标记在甘蓝型油菜19条染色体上的位置, 挑取11 368个均匀分布于染色体上的SNP, 以Structure 2.3.4[19]进行群体结构分析获得Q矩阵, 不作数迭代(length of burn-in period)设置为50 000, MCMC重复为100 000, 组群数(K)设定为1~10, 重复5次。利用STRUCTURE HARVESTER (http://taylor0. biology.ucla.edu/structureHarvester/)[20]选择合适的K值作为最适亚群数。将基因型数据导入Tassel 5.2.12[21]后进行主成分分析(PCA矩阵), 亲缘关系分析(K矩阵)和LD分析。然后, 将Q矩阵导入Tassel 5.2.12, 以Q、K和PCA矩阵作协变量, 采用基于一般线性模型(general linear model, GLM)的naive、Q和PCA模型和混合线性模型(mixed linear model, MLM)的K、Q+K和PCA+K共6种模型进行关联分析, 显著关联SNP阈值设为1/31832 = 3.14E-5。根据所有SNP的-lg (P)观察值和期望值, 以GGplot2[22]绘制Quantile-Quantile散点图(QQ plot), 确定每个性状GWAS分析的最佳模型。基于最优模型的关联分析结果, 利用QQman绘制曼哈顿图[23]。
利用Tassel 5.2.12分析LD在甘蓝型油菜各染色体上的分布, 绘制各染色体的LD衰减图, LD类型参数设置为Full Matrix。为确定候选基因区间, 以Haploview 4.2[24]计算显著关联SNP所在染色体的LD, HW阈值(Hardy Weinberg P-value cutoff)设定为0.001; 非缺失标记的比例为75%, MAF设定为0.05; 单倍型块(haplotype block)分析采用solid spine of LD算法, D′ 设为0.8, 采用显著关联SNP所在单倍型块作为候选基因所在LD区间。若显著关联SNP未位于单倍型块内, 则以其两侧SNP标记上下游50 kb的侧翼序列区间作为LD区间, 用于候选基因预测和功能注释分析。
根据LD区间在油菜基因组中的位置, 以法国公布的甘蓝型油菜品种“ Darmor-Bzh” 的基因组[15]注释信息(http://www.genoscope.cns.fr/brassicanapus)分析LD区间内的基因数目和基因编码蛋白序列。利用本地Blast程序, 将待分析的蛋白序列与拟南芥(Arabidopsis thaliana)所有蛋白序列进行BlastP比对, E-value阈值为1E-10, 以同源性最高的拟南芥基因注释候选基因功能。为了更准确地了解LD区间候选基因功能, 在拟南芥信息资源网(The Arabidopsis Information Resource, https://www.arabidopsis.org/)对拟南芥同源基因的基因表达模式、亚细胞定位、可能的共表达基因和互作蛋白进行了分析, 以筛选控制目标性状的重要候选基因。
对重庆和云南环境共412份自然群体材料结角高度与结角层厚度统计分析(图1和表1)表明, 目标性状均表现出广泛的表型变异, 在重庆的变异幅度分别为62.00~153.00 cm和7.13~142.88 cm, 变异系数为13.84%和30.42%; 而在云南的变异幅度分别为70.60~183.00 cm和11.20~99.40 cm, 变异系数为19.03%和20.11%。以两地均种植的98份材料进行差值分析(重庆性状值-云南性状值), 结果表明结角高度差与结角层厚度差的变异范围分别为-136.00~ -19.00 cm和-70.08~86.50 cm, 说明2个性状在环境之间存在较大变异。Shapiro-Wilk检验表明, 所有性状W值均在0.96~0.99之间, 除云南和重庆的结角层厚度外, 其他表型P-value值均未达到显著差异水平, 符合正态分布。虽然云南和重庆结角层厚度未通过正态分布检验, 但从频次分布图(图1)和Shapiro-Wilk检验W值(表1)可看出, 其频次分布具有明显的正态分布特征, 表明本研究性状均为数量性状, 采用GWAS分析方法可有效促进目标性状定位。
| 表1 甘蓝型油菜结角高度与结角层厚度的统计分析 Table 1 Statistical analysis of HP and TPC in B. napus |
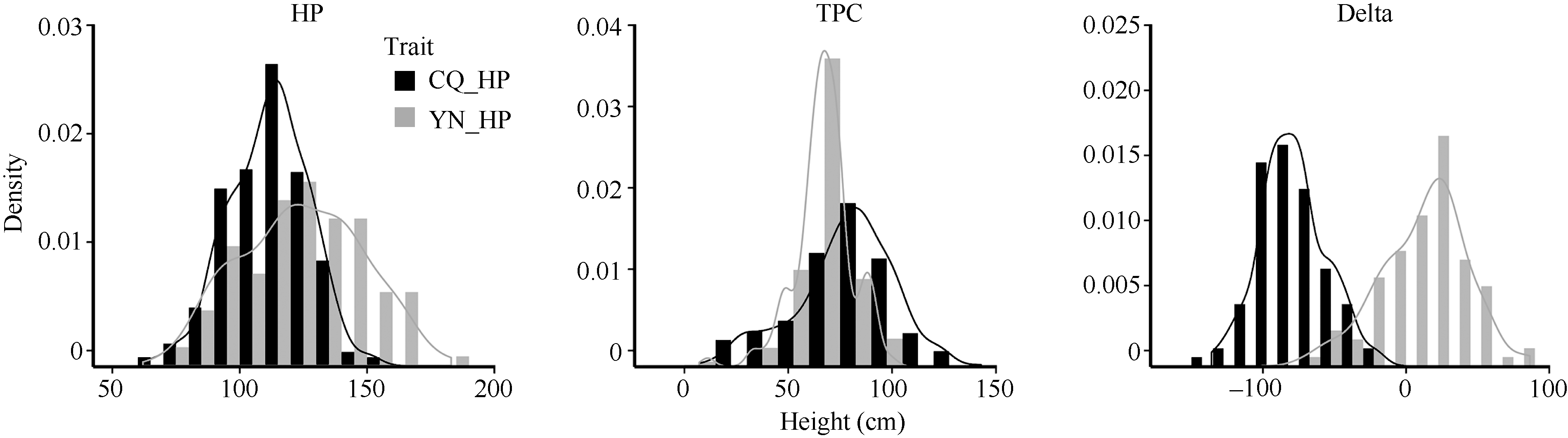 | 图1 自然群体结角高度与结角层厚度的频次分布Fig. 1 Frequency distribution of HP and TPC from the natural population |
对重庆环境结角高度(图2-A)及2个环境结角层厚度(图2-D和2-E)分析表明, MLM+K、MLM+K+Q和MLM+K+PCA模型均能较好地控制假阳性, 但MLM+K+PCA模型检测到P-value较其他模型更接近期望值。云南环境结角高度(图2-B)及两地结角层厚度差(图2-F)以GLM+PCA模型为最佳。对两地结角高度差(图2-C), GLM模型能较好地控制假阳性产生概率。
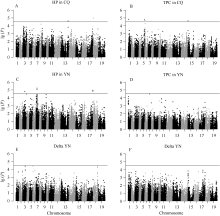
结角高度和结角层厚度GWAS (表2和图3)结果表明, 共检测到16个显著关联的SNP。重庆环境下检测到2个和4个SNP与结角高度和结角层厚度显著关联, 单个SNP解释的表型变异为5.61%~ 5.69%和5.94%~6.31%, 分别位于A03、A06、A07、C03和C08染色体。云南环境下对结角高度和结角层厚度分别检测到5个和1个显著关联的SNP, 单个标记解释的表型变异为12.66%~13.97%和22.43%, 分别位于A03、A07和C07染色体。2个环境下对结角高度差和结角层厚度差共检测到3个和1个显著相关的SNP, 它们对表型变异的解释率分别为17.33%~20.32%和29.05%, 分别位于A03、A06、C02和C08染色体。
从图4可以看出, 甘蓝型油菜A基因组的平均LD衰减距离小于C基因组。以决定系数(coefficient of determination) r2=0.1为衰减阈值时, A基因组的A08、A09和A10染色体LD衰减较快, 衰减距离约为2 Mb, 而其他A基因组的LD衰减距离约为500 kb; C基因组中C01和C02的衰减速度最慢, 衰减距离为10 Mb, C06的衰减速度最快, 衰减距离约为1 Mb。这些结果表明, A基因组较C基因组衰减速度更快, 可能与中国半冬性甘蓝型油菜A基因组在育种中发生大规模重组, 打破连锁不平衡有关。
 | 图2 比较6种模型的结角高度和结角层厚度GWAS的QQ图Fig. 2 Quantile-quantile plots of estimated -lg (P) from association analysis using six methods for HP and TPC traits |
| 表2 结角高度与结角厚度的显著关联标记 Table 2 Significant association markers of HP and TPC |
 | 图3 结角高度和结角层厚度全基因组关联分析的曼哈顿图Fig. 3 Manhattan plots of GWAS for HP and TPC traits |
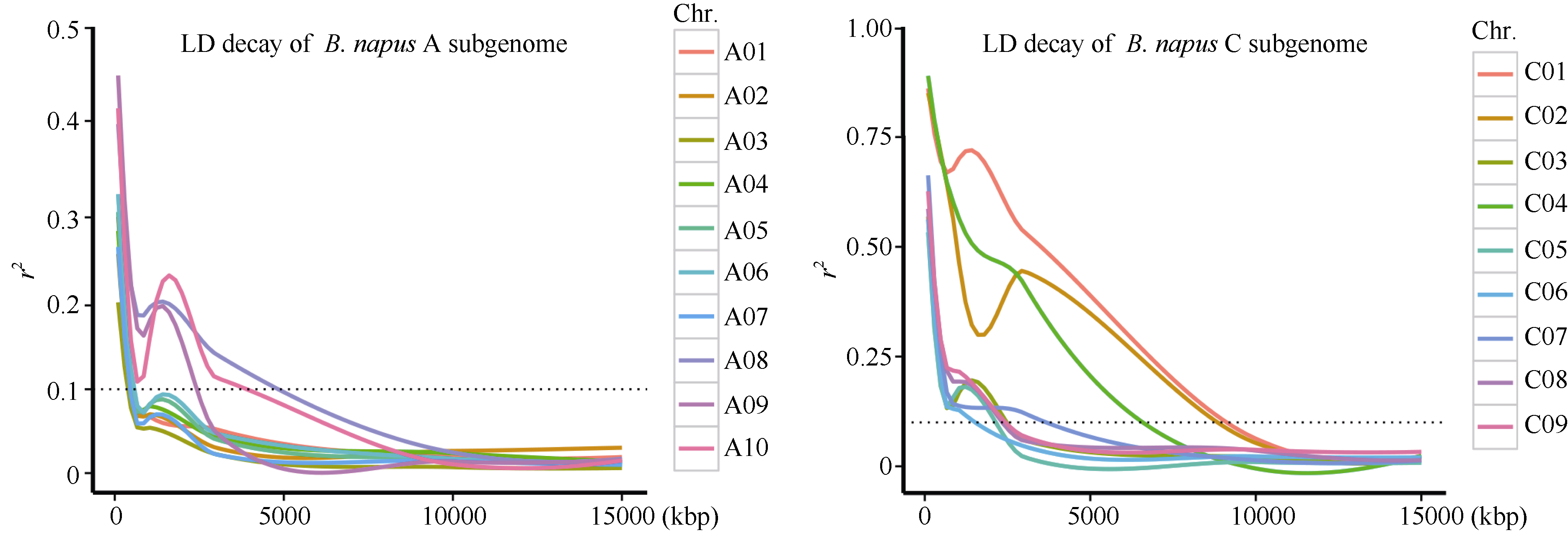 | 图4 甘蓝型油菜A和C基因组不同染色体的LD衰减Fig. 4 LD decay of different chromosomes for A and C subgenomes in B. napus |
显著关联SNP的LD区间和候选基因分析结果如表3所示。共检测到5个单倍型区块, 包括6个SNP, 其中A07染色体上与云南结角高度相关的2个SNP位于同一单倍型块(表3)。虽然C07上与云南结角高度相关的2个SNP并未在同一单倍型块中, 但2个SNP仅相隔450 bp, 因此将其作为同一LD区间。与重庆结角层厚度相关的Bn-scaff_16414_ 1-p836176和两环境结角层厚度差相关的Bn-scaff_ 16214_1-p321530位于C05染色体同一LD区间内, 表明该区间存在与结角层厚度相关的重要候选基因。但对结角高度来说, 重庆、云南及两地差值相关SNP无重叠LD区间, 说明结角高度受环境影响较大, 可能需要多年多点试验才能鉴定出稳定的显著关联SNP或QTL。
基于油菜候选基因序列及拟南芥同源基因的功能分析[14], 对LD区间的候选基因进行了功能预测(表3)。结果表明, 结角高度相关LD区间的基因分布于A03、A06、A07、C03、C07和C08染色体, 基因数为42、28、44、11、46和34个; 结角层厚度相关LD区间的基因位于A01、A05、A06、A07和C05染色体, 基因数分别为38、105、48、51和38个。在上述基因中, 存在多个调节植物细胞组织发生(cell organization and biogenesis)、花分生组织发育(floral meristem development)、角果数目(number of silique)和多器官发育(multicellular organismal development)的基因, 它们可能通过上述功能影响油菜花序或角果的发育, 导致结角高度或结角层高度差异。
| 表3 显著关联标记所在LD区间及候选基因 Table 3 LD intervals and candidate genes of significant association markers |
油菜结角层是由角果和果序轴形成的冠层结构, 包含了油菜所有的产量构成因素(角果数、每角粒数、粒重)和后期的源库结构关系, 既是油菜生育后期主要的光合器官, 也是光合产物贮藏器官和种子存储器官[25]。研究表明, 油菜结角层为籽粒产量贡献了50%~70%的光合生产力[26], 是籽粒产量的主要来源, 而茎秆和叶片的光合产物对产量的贡献较少, 均只有10%左右[27]。虽然结角层对油菜产量具有重要贡献, 但目前关于油菜结角层的研究还停留在对理想株型的分析上, 遗传和定位研究还非常少。漆
丽萍[9]利用181个DH株系对油菜株型及产量相关的16个性状分析, 表明角果层长度(对应本研究的结角厚度)和角果层宽度呈显著正相关, 2个性状与单株产量也极显著正相关, 相关系数分别为0.70和0.61, 表明这2个性状与产量的关系密切; 检测到10个角果层长度相关QTL, 分布在A01、A03、A06、A09、C03和C08连锁群, 单个QTL解释的遗传变异为5.1%~11.8%; 20个角果层宽度相关QTL分布在A01、A02、A03、A05、A07和A09连锁群, 单个QTL解释的遗传变异为4.2%~31.3%。本研究在A01和A06染色体检测到结角层厚度相关SNP, 与漆丽
萍[9]检测到qLSL.A1.1和qLSL.A6物理距离分别为600 kb和9.36 Mb, 表明A01上结角层厚度相关SNP与qLSL.A1.1可能位于同一QTL区间, 而A05、A06、A07和C05上检测到的SNP很可能为新的结角层厚度相关位点。
对重庆、云南及两地差值相关SNP的LD区间比较, 发现仅有C05染色体上重庆结角层厚度相关SNP与两环境结角层厚度差相关SNP位于同一LD区间内, 解释的遗传变异为29.05%, 这可能与不同甘蓝型油菜材料在该LD区段存在对重庆环境差异响应位点有关。此外, 该位点可能与云南环境不同材料结角层厚度变异无显著相关性, 导致其只能在重庆环境和两地差值条件下检测到, 在云南环境下无法检测到。重庆和云南环境下特异检测到的多个显著关联SNP可能与基因和环境互作有关。云南临沧是我国早熟油菜高产区, 2014年专家测产, 11.2 km2连片油菜的平均理论产量为5412 kg, 高产田实收产达6745.35 kg km-2, 而相同材料在重庆种植的产量仅为云南临沧的一半。因此, 我们非常关注云南高产环境特异的显著关联SNP, 这些位点可能对云南特殊生产环境的高产具有重要贡献, 进一步精细定位并克隆这些高产相关基因, 不仅能促进油菜高产机制的理解, 而且能为我国长江流域主产区的油菜增产提供高产基因资源。如C03染色体的BnaC03g64230D, 在拟南芥中的同源基因AT3G07050编码类核干细胞因子1 (nucleostemin-like 1, NSN1), 通过调节核小体发生实现植物花序分生组织的发育调控[28]; A06染色体的BnaA06g04490D与拟南芥AT1G08030同源, 编码酪氨酸硫化转移酶(tyrosylprotein sulfotransferase, TPST), 对维持植物根部发育及花和角果数具有重要作用[29]; A07染色体的BnaA 07g10360D编码SAC1 (suppressor of actin 1)蛋白, 可能通过细胞形态发生和细胞壁合成实现对植物花序发育的调节[30], 这些基因可能影响油菜花序或角果发育, 导致结角高度或结角层高度差异。虽然, 基因与环境互作导致环境特异关联SNP很难通过分子标记辅助选择促进其他地区油菜增产, 但通过基因工程手段对部分高产基因进行操作很可能实现其他地区油菜产量的提升, 对我国油菜生产和发展具有重要意义。
重庆和云南环境下分别检测到2个和5个结角高度显著关联SNP, 结角层厚度显著关联SNP 4个和1个; 两地结角高度差和结角层厚度差显著关联SNP为3个和1个。除C05染色体上重庆及两地结角层厚度相关SNP位于同一LD区段外, 其他SNP均为环境特异, 表明其受基因与环境互作影响。LD区段内调节植物细胞组织发生、花分生组织发育、角果数目和多器官发育基因NSN1、TPST和SAC1等可能通过上述功能影响油菜花序或角果的发育, 导致结角高度或结角层厚度差异。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|