



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
导入外源玉米C4型 NADP-ME基因对小麦光合效能的影响
[王永霞1, 2  , 杜新华
, 杜新华1, 2 , 许为钢1, 2, * , 齐学礼2 , 李艳2 , 王会伟2 , 胡琳2 ]
, 杜新华, 齐学礼|
|


第一作者联系方式: E-mail: wangyongxia005@163.com, Tel: 0371-65721637
为研究玉米C4型 NADP-ME基因对小麦光合特性的影响, 以T3代转 NADP-ME基因小麦株系10T(9)-1-1和10T(9)-225-4及其对照周麦23为试材, 进行了分子特征鉴定、光合生理特性分析和单株产量性状调查, 并对其作用机制进行了初步解析。结果表明, 外源基因已整合到转基因小麦的基因组中, 并能正确转录和翻译; 与对照周麦23相比, 两个转基因株系在4个测定时期旗叶NADP-ME酶活性均明显提高, 而旗叶净光合速率( Pn)则明显下降; 尤其在花后第7天, 两个转基因株系的酶活性较对照分别提高1.33倍和1.13倍, Pn分别较对照下降17.26%和10.35%; 千粒重和籽粒产量较对照均显著下降。与对照周麦23相比, 转基因株系10T(9)-225-4利用强光和同化CO2能力下降, 光合效率下降; 旗叶气孔张开率和气孔导度均显著下降; 苹果酸含量降低了5.6%, 丙酮酸含量则提高了17.1%; 外施苹果酸可恢复其光合速率。以上研究结果表明, 玉米C4型 NADP-ME基因的导入降低了小麦的光合特性, 苹果酸含量降低引起的气孔导度下降可能是导致转基因小麦光合效能降低的原因。
To explore the physiological characteristics of the transgenic wheat expressing maize C4-type NADP-ME, we introduced NADP-ME into the C3 crop wheat by using particle bombardment transformation. Two transgenic wheat lines (10T(9)-1-1, 10T(9)-225-4) and parental control (Zhoumai 23) were used to study molecular characteristics and photosynthesis property, to reveal the mechanism. The results showed that the NADP-ME sequence was integrated into wheat genome, and the transcription and translation were exactly same as expect. The enzyme activity of NADP-ME in flag leaf in transgenic plants were increased significantly than untransformed plants, for instance it was increased 1.33 and 1.13 times on the 7th day after flowering. Net photosynthetic rate ( Pn) of flag leaf in transgenic plants obviously decreased when compared to the untransformed plants. On the 7th day after anthesis, Pn of transgenic wheat decreased by 17.26% and 10.35%. The yield and 1000-grain weight were decreased than the control. Utilization efficiency on strong light utilizing and ability of CO2 assimilation in transgenic line 10T(9)-225-4 were significantly declined, photosynthesis rate was also decreased. Stomatal opening rate and stomatal conductance were significant decrease, malic acid content of transgenic wheat reducing 5.6% while pyruvate level is raised by 17.1%, and Pn of transgenic wheat can be restored by feeding with exogenous malate. Those results indicated that the transgenic wheat expressing maize NADP-ME gene showed lower photosynthetic characteristics than the control, the reason was maybe the decrease of stomatal aperture caused by decline of malic acid content.
长期以来, 许多科学家都在努力将C4作物的高光效关键基因引入水稻、小麦等C3作物中, 试图利用C4型高光效基因来改良C3作物光合作用的内在遗传基础, 使之具备高光效特性, 从而大幅度提高光合效率, 实现生物学产量和籽粒产量的同步提高[1, 2]。随着分子生物学和植物转基因技术的快速发展, 通过基因工程手段将C4途径光合关键酶基因导入C3植物中, 在国内外已取得了显著进展[3, 4]。特别是在水稻、小麦、烟草等植物中转入C4途径高光效关键基因PEPC和PPDK, 并获得高效表达的转基因植株的成功报道[5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16], 为进一步提升C3植物产量潜力开辟了一条新的道路。NADP-ME也是C4途径的关键酶之一, 具有催化苹果酸脱羧, 释放的CO2富集在Rubisco周围, 从而促进Rubisco的羧化反应, 降低其加氧反应, 提高光合作用效率作用。Ku等[17]认为NADP-ME基因的活性与光呼吸的速率呈负相关, 将NADP-ME基因转入C3植物, 可能是降低C3植物的光呼吸, 提高其光合效率的一种有效途径。目前, 已成功的将C4型NADP-ME基因导入到拟南芥、烟草和水稻等上C3植物上, 但大多数结果是对光合产量的提高无实质性的效果[18, 19, 20, 21, 22, 23]。Tsuchida等[21]发现在过量表达玉米叶绿体NADP-ME cDNA的水稻转化体中, NADP-ME酶活性比对照增加30倍左右, 转化体叶色变白并且生长明显受到抑制。迟伟等[22]研究结果显示, 在转高粱C4型NADP-ME cDNA的转基因水稻植株中, NADP-ME酶活性比对照提高了1~7倍, 但光合速率没有提高。Laporte等[23]发现, 转玉米NADP-ME基因的烟草的气孔导度与NADP-ME的水平成反比, 转基因水稻的水分利用效率(鲜重/水耗)比野生型高15%~20%。截至目前, 外源C4型NADP-ME基因对小麦光合效能的影响及其机制研究尚未见报道。
本课题组从玉米高光效材料中分离了C4型NADP-ME基因的cDNA序列[24], 并将其导入拟南芥[25]和小麦中。本研究以T3代转玉米C4型NADP-ME基因小麦株系为材料, 研究该基因在小麦基因组中的整合与表达情况, 光合生理特性和产量性状的表现, 并对其影响机制进行了初步解析, 为合理利用C4型NADP-ME基因改良C3作物光合效能提供依据。
从玉米高光效自交系Z1194中分离了C4型NADP-ME基因的cDNA序列[24], 将其构建到含35S启动子的双元表达载体pCambia3301中, 通过基因枪法导入普通小麦品种周麦23中, 获得的T3代转NADP-ME基因小麦株系10T(9)-1-1和10T(9)-225-4为试验材料。
2012年10月, 将各转基因小麦株系和对照周麦23种植于国家黄淮海转基因小麦中试基地(河南原阳, 35° 00′ N, 113° 40′ E), 每系行长2 m, 行距25 cm, 株距10 cm。田间常规管理。
1.2.1 Southern杂交检测
2013年4月7日, 取拔节期小麦叶片3 g, 用改良的CTAB法提取基因组DNA[26]。取150 μ g基因组DNA和50 ng质粒DNA, 使用限制性内切酶XbalI酶切消化, 纯化后电泳分离, 转移至尼龙膜(N+)上。以质粒pCambia3301-NADP-ME为模板, 使用引物扩增520 bp的NADP-ME基因片段为探针标记模板。取外源基因探针模板1 μ L, 用DIG标记及检测试剂盒(德国Roche公司)进行探针标记和杂交, 暗室中过夜、压片并检测信号。
1.2.2 qRT-PCR分析 2013年分别在抽穗期(4月25日)、开花期(5月2日)、花后第7天(5月13日)和花后第15天(5月20日), 选取100 mg旗叶, 采用植物总RNA提取试剂盒(天根生物科技有限公司)提取总RNA。RNA反转录为cDNA (Promega公司试剂盒), 使用SYBR Green Real-time PCR Master Mix QPK-201 (TOYOBO), 在Bio-Rad iQ5上进行实时荧光定量PCR反应。扩增引物由生工生物工程(上海)有限公司合成。NADP-ME基因引物(QF: 5'-CGTG GAGTACGAAGGAAAGACT-3'; QR: 5'-GAGATCTTTC TGATGCTGGTGA-3')。扩增程序为: 94℃预变性2 min; 94℃变性15 s, 58℃复性10 s, 72℃延伸15 s, 38个循环。以小麦肌动蛋白β -actin为内参照基因(引物序列AF: 5'-GTTCCAATCTATGAGGGATACACGC-3'; AR: 5'-GAA CCTCCACTGAGAACAACATTACC-3')。扩增程序为: 94℃预变性2 min; 94℃变性15 s, 55℃复性15 s, 72℃延伸15 s, 35个循环。所有试验样品均3次重复。
1.2.3 Western杂交检测
为进一步验证玉米C4型NADP-ME基因在小麦中能否正确翻译出相应的蛋白, 在抽穗期对T3代转基因小麦进行Western杂交分析。参照Ku等[17]描述的方法从抽穗期(2013年4月25日)旗叶中提取粗蛋白。取20 μ g粗蛋白样品, 经SDS-PAGE、100 V湿转2 h转移到硝酸纤维素膜上, 新鲜脱脂奶粉封闭1 h后加入一抗(NADP-ME兔多抗)孵育2 h, TBST洗膜3次, 每次5 min, 再用HRP标记的二抗(山羊抗兔)孵育1 h, TBST洗膜6次, 每次5 min, 最后用ECL发光检测试剂盒暗室发光显影, 用Quantity One软件进行杂交条带的灰度分析。NADP-ME兔多克隆抗体由美国Abcam公司制备, HRP标记的二抗购自北京康为世纪生物科技有限公司。
1.3.1 NADP-ME酶活性测定
2013年3月12日小麦返青期, 将转基因植株和对照周麦23移植于直径285 mm、高386 mm花盆内, 每份材料移植30盆, 每盆定苗3株, 常规管理。参照Ku等[5]描述的方法测定抽穗期(4月25日)、开花期(5月2日)、花后第7天(5月13日)和花后第15天(5月20日)旗叶中NADP-ME酶活性。
1.3.2 气体交换参数测定
2013年, 分别在抽穗期(4月25日)、开花期(5月2日)、花后第7天(5月13日)和花后第15天(5月20日), 利用CIRAS-2光合仪测定转基因小麦和对照旗叶的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci)日变化。测定时CO2浓度为大气CO2浓度, 气体流速为300 mL min-1。测定时间从6:00到18:00, 每间隔1 h往返重复测定3次。参照王兰兰等[27]描述的方法计算单位日光合总量。测定时段的光合有效辐射和大气温度见表1。
| 表1 5个生育期的最大光合有效辐射和最高温度 Table 1 Maximal values of photosynthetically active radiation(PAR) and temperatures at five stages |
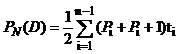
式中, PN(D)为单位日光合总量值(μ mol m-2); Pi为i次测光合速率(μ mol m-2s-1); ti为Pi与Pi+1次测定间隔时间(s); n为测定次数。
1.3.3 旗叶Pn对光强和CO2响应曲线测定
2013年在开花期(5月2日) 9:00— 11:30测定光合速率(Pn)对光强(PPFD)的响应曲线, 叶室温度为29℃, 叶室光强设定梯度依次为1800、1600、1400、1200、1000、800、600、400、200、150、80、40、10 μ mol m-2 s-1, CO2浓度为380 μ mol mol-1。测定光合速率(Pn)对CO2浓度响应的相应曲线, CO2浓度由系统配置的CO2钢瓶提供。利用初始期胞间CO2浓度(Ci) 35~215 μ mol mol-1的Pn与Ci进行回归, 其斜率为羧化效率。叶室光强为1450 μ mol m-2 s-1, 叶室温度为29℃。
气孔张开率是指视野内开放气孔数与总气孔数的百分比。2013年在抽穗期(4月25日), 剪取小麦叶片中部约5 cm长, 迅速投入到FAA固定液(甲醛5 mL+冰醋酸5 mL+70%酒精90 mL)中。将固定好的样品叶片表皮上的溶液擦拭干净, 于其上、下表皮均匀涂上一层薄薄的透明的指甲油。风干后将指甲油层用透明胶粘住剥取, 平放在载玻片上, 在400倍显微镜下观测, 分别统计视野内开放和关闭的气孔数, 所得数据为10个视野的平均值。气孔导度采用CRIAS-2光合仪测定。所有样品各选取5片旗叶进行测试指标分析。
2013年在抽穗期(4月25日)的上午9:30— 11:30, 按照Ji等[30]方法将苹果酸溶液(150 mmol L-1)均匀喷施于转基因小麦株系10T(9)-225-4及对照旗叶表面, 每个处理选取5片旗叶, 喷施蒸馏水作为对照组。将处理后的植株置于暗处(20~30 μ mol m-2 s-1)下处理1 h, 使苹果酸溶液和蒸馏水充分渗入叶片组织内。自然光下适应1 h后, 采用CIRAS-2型光合作用系统测定其光合速率的变化, 重复往返测定3次。
2013年6月, 收获后取自然风干的转基因植株和对照各30株, 调查株高、单株穗数、小穗数、穗粒数、千粒重和单株籽粒重, 使用DPS7.55统计软件对转基因株系和对照各项指标进行统计分析和显著性比较。
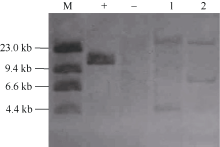
2.1.1 Souhern杂交结果 对3个转基因株系中PCR阳性的T3代植株和对照周麦23的叶片提取基因组DNA进行Southern杂交分析。经限制性内切酶XbalI酶切消化的阳性质粒与探针杂交后出现1条11.5 kb左右的杂交带, 受体对照植株中无杂交信号, PCR检测阳性的转基因植株中出现有杂交带(图1), 表明外源NADP-ME基因已整合到小麦基因组中。
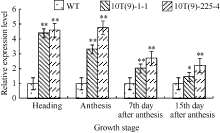
2.1.2 转录水平分析 对不同生育期转基因小麦中NADP-ME的转录水平进行分析, 受体对照中出现cDNA扩增产物, 说明小麦中可能存在有內源NADP-ME基因。外源NADP-ME基因导入后, 2个转基因株系中NADP-ME基因的相对表达量与对照周麦23相比均显著提高(P < 0.01), 且开花期> 抽穗期> 花后第7天> 花后第15天。其中, 转基因株系10T(9)-1-1和10T(9)-225-4的NADP-ME基因表达量, 开花期分别高2.32倍和3.75倍, 抽穗期分别较对照高3.41倍和3.60倍(图2)。
 | 图1 T3代转基因株系的基因组Southern杂交分析 M: λ DNA; +: 质粒pMW003-NADP-ME; -: 受体周麦23; 1~2转基因植株依次为10T(9)-1-1和10T(9)-225-4。Fig. 1 Southern blot analysis of T3 transgenic lines M: λ DNA; +: Plasmid pMW003-NADP-ME; -: recipient Zhoumai 23; 1-2: transgenic plants 10T(9)-1-1 and 10T(9)-225-4. |
2.1.3 翻译水平检测
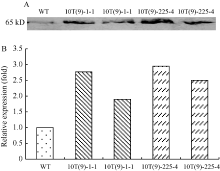
Western杂交结果显示, 转基因小麦和受体对照中均出现65 kD的目的蛋白, 说明NADP-ME抗体能与小麦内源NADP-ME杂交, 不同转基因单株中NADP-ME翻译水平上存在差异(图3-A); 杂交条带灰度分析显示, 转基因株系10T(9)-1-1和10T(9)-225-4分别比对照高0.90~1.77倍和1.50~1.95倍(图3-B)。
外源NADP-ME基因的导入, 使两个转基因株系中NADP-ME酶活性均不同程度的增强。在4个测定生育期, 除10T(9)-1-1在花后第15天与对照未达显著差异外, 两个株系的旗叶NADP-ME酶活性均比对照显著提高(图4-A)。其变化规律为花后第7天> 开花期> 抽穗期> 花后第15天; 其中, 花后第7天转基因小麦NADP-ME酶活性提高幅度最大, 分别较对照高1.33倍和1.13倍。
 | 图2 转基因小麦及其对照(WT)不同生育期旗叶NADP-ME基因的相对表达量 内参基因为β -actin。* * 表示转基因植株与对照之间差异显著(P< 0.01)。Fig. 2 Relative expressions of NADP-MEin flag leaves of transgenic wheat lines and wild type (WT) at different stages Gene β -actinwas used as the internal reference. * * indicates significant difference between the transgenic plant and WT (P< 0.01). |
 | 图3 T3代转基因株系Western杂交(A)和杂交图片灰度分析(B)结果 以WT的表达蛋白含量为1。Fig. 3 Western blot (A) and gray level analysis (B) of T3 transgenic lines The expression protein content of WT was 1. |
外源NADP-ME基因导入后, 4个测定生育期的转基因小麦旗叶Pn均比对照明显下降, 除花后第15天转基因小麦的Pn与对照无显著差异外, 其他生育期两转基因株系Pn均显著低于对照(图4-B); 花后第7天下降幅度最大, 10T(9)-1-1下降了17.26%, 10T(9)-225-4下降了10.35%, 其次为抽穗期, 10T(9)-1-1和10T(9)-225-4 Pn降幅分别为16.15%和9.26%。
与对照周麦23相比, 转基因小麦株系10T(9)-1-1和10T(9)-225-4的各产量性状指标均不同程度降低, 但只有籽粒产量(P=0.015与0.028)和千粒重(P=0.020与0.025)达到显著水平, 其中千粒重平均下降10% (表2)。结果表明, 玉米C4型NADP-ME基因的引入导致转基因小麦千粒重下降, 进而造成籽粒减产。
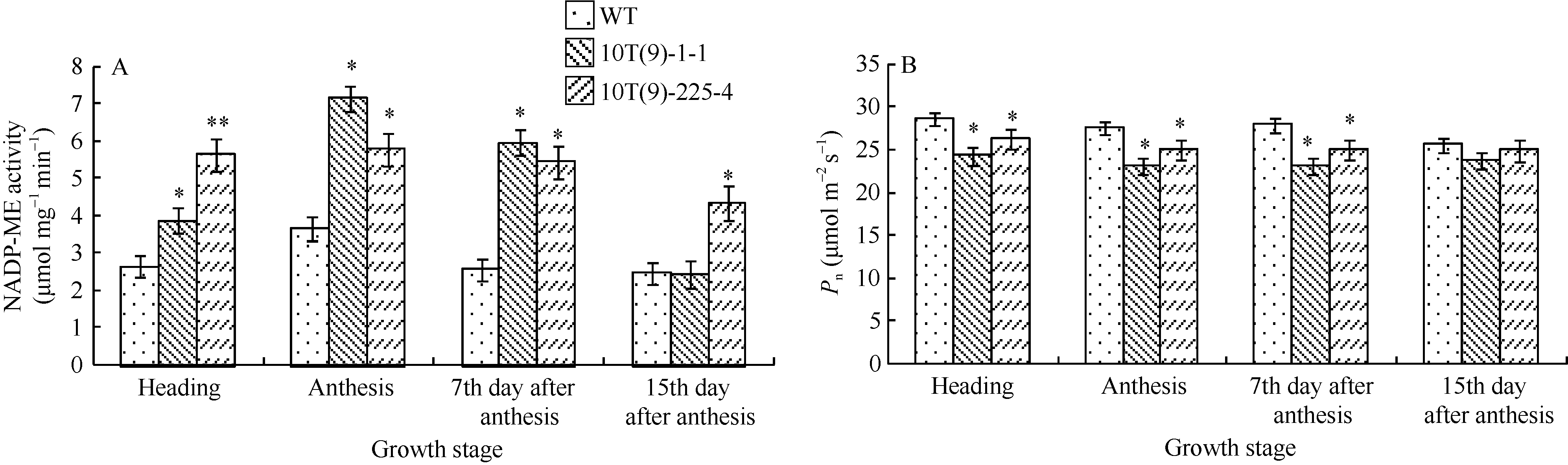 | 图4 不同生育期转基因小麦及对照(WT)旗叶NADP-ME酶活性(A)和净光合速率(B)的变化 * 和* * 分别表示转基因植株与对照在P< 0.05和P< 0.01水平差异显著。Fig. 4 Dynamic changes of NADP-ME enzyme activity(A) and photosynthetic rate (B) in flag leaves of transgenic wheat lines and the wild type (WT) at different stages * and * * indicate significant difference between the transgenic plant and WT at P< 0.05 and P< 0.01 level, respectively. |
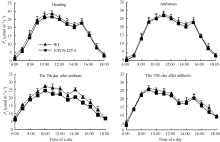
4个测定生育期转基因小麦株系10T(9)-225-4和对照周麦23的Pn日变化均表现为双峰曲线, 转基因小麦的Pn低于对照周麦23, 其差异在花后第7天最明显(图5), 此时转基因小麦旗叶Pn的最大值为24.59 μ mol m-2s-1, 较对照周麦23下降了10.26%。转基因小麦Gs、Tr和Ci等光合特性参数的日变化呈现与对照一致的趋势, 但转基因小麦的Ci明显高于对照周麦23, 而Gs和Tr明显低于对照周麦23 (图6)。
| 表2 转NADP-ME基因小麦和非转基因对照的产量及农艺性状比较 Table 2 Comparison of yield related traits betweenNADP-ME transgenic wheat lines and WT |
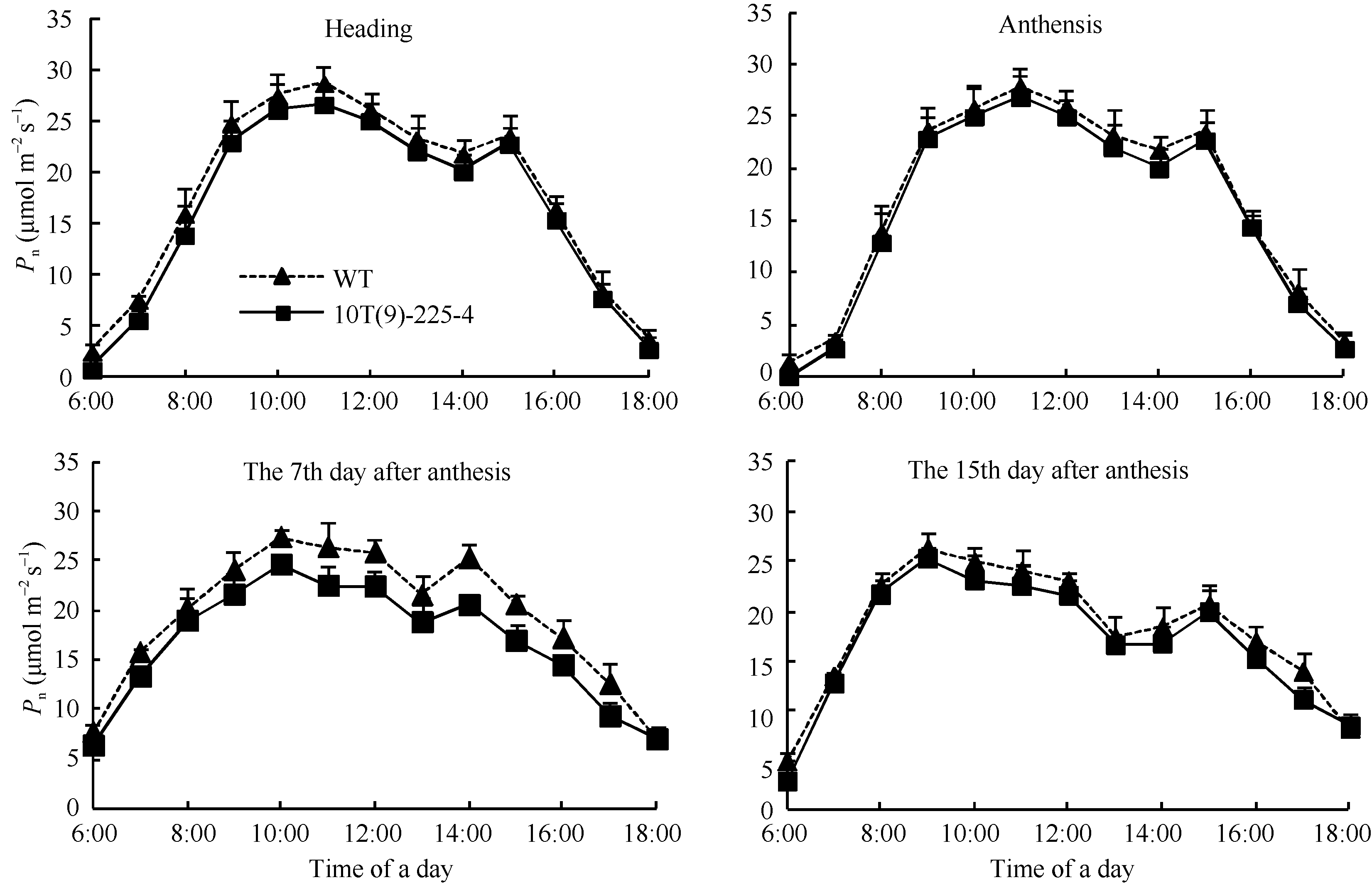 | 图5 转基因株系和对照(WT)不同时期旗叶净光合速率日变化曲线Fig. 5 Diurnal variation of photosynthetic rate of flag leaves in transgenic line and wild type (WT) at different stages |
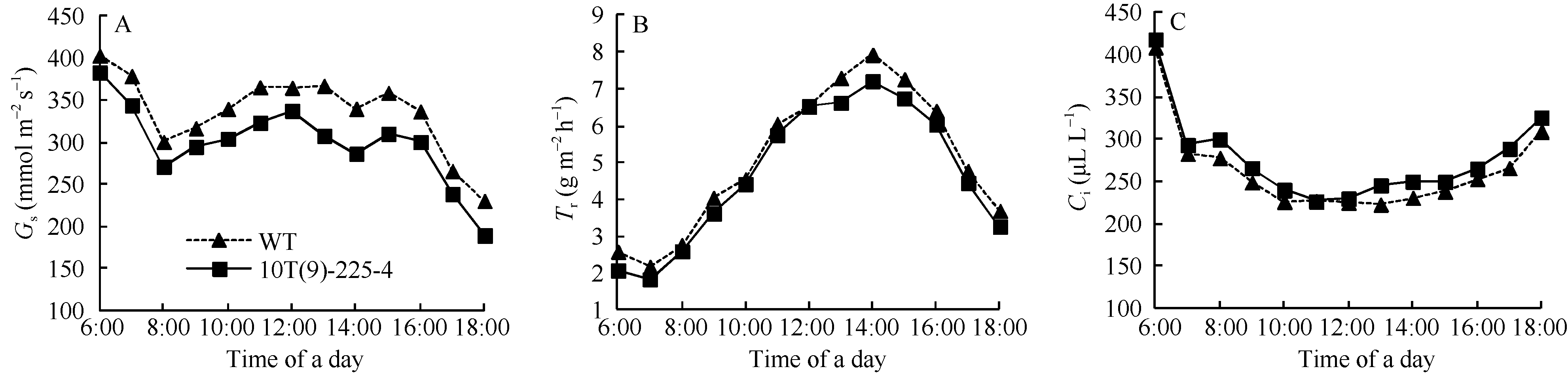 | 图6 转基因株系和对照(WT)不同时期旗叶Gs、Tr和Ci平均日变化曲线 图中数值均为4个生育期测定数值的平均值。Fig. 6 Average diurnal variation of Gs, Tr, and Ci of flag leaves in transgenic line and wild type (WT) at different stages Data are the mean of four growth stages. |
| 表3 转NADP-ME小麦不同生育时期的单位日光合总量 Table 3 Diurnal photosynthesis cumulative Pn in NADP-ME transgenic wheat lines at different growth stages (× 105 μ mol m-2) |
4个测定生育期转基因小麦株系10T(9)-225-4平均日光合总量与对照周麦23相比均明显下降, 尤其在花后第7天, 比对照下降13.93% (表3)。上述结果表明, 转基因小麦株系10T(9)-225-4的光合效率低于对照周麦23。
转基因小麦株系10T(9)-225-4和对照周麦23的饱和光强分别为1400.33 μ mol m-2 s-1和1599.33 μ mol m-2 s-1, 比对照周麦23下降12.44% (图7-A); 羧化效率分别为0.055和0.085, 较对照周麦23下降35.29%; CO2补偿点分别为102.00 μ mol mol-1和86.50 μ mol mol-1, 较对照周麦23高17.92% (图7-B)。上述结果表明, 与对照周麦23相比, 转基因小麦株系10T(9)-225-4利用强光和同化CO2能力下降。
2.7.1 气孔张开率和气孔导度变化
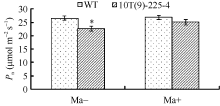
与对照周麦23相比, 转基因小麦株系10T(9)-225-4的气孔并没有完全张开, 气孔张开率为43.54%, 比对照周麦23 (51.32%)下降了15.16%。在气孔数量上两者无明显差异(图8)。转基因小麦株系10T(9)-225-4的气孔导度为253.30 mmol m-2 s-1, 比对照周麦23 (288.49 mmol m-2 s-1)下降了12.18%; 旗叶Pn为22.85 μ mol m-2 s-1, 比对照周麦23 (26.22 μ mol m-2 s-1)下降了12.85%。结果表明, 转基因小麦中气孔导度的下降是其净光合速率降低的原因。
2.7.2 苹果酸和丙酮酸含量变化
与对照周麦23相比, 抽穗期转基因小麦株系10T(9)-225-4的苹果酸含量降低了8.89%, 而丙酮酸含量则和25.00%, 差异达到显著水平(表4)。结果表明, 转基因小麦气孔导度的下降可能是由于其叶片中NADP-ME酶活性的增强, 使得苹果酸浓度降低造成的。
2.7.3 喷施外源苹果酸旗叶Pn变化
喷施苹果酸前, 转基因小麦株系10T(9)-225-4的旗叶Pn为22.77 μ mol m-2 s-1, 较对照周麦23 (26.67 μ mol m-2 s-1)下降了12.20%, 差异达到显著水平; 喷施苹果酸后, 转基因小麦株系10T(9)-225-4的Pn达到25.15 μ mol m-2 s-1, 比喷施前提高了9.46%, 但与喷施苹果酸后的对照周麦23 (26.98 μ mol m-2 s-1)相比两者无明显差异(图9)。外施苹果酸可恢复转基因小麦的光合特性, 由此可见, 转基因小麦气孔导度的下降是由于叶肉细胞苹果酸含量下降所导致的, 进而导致其净光合速率下降。
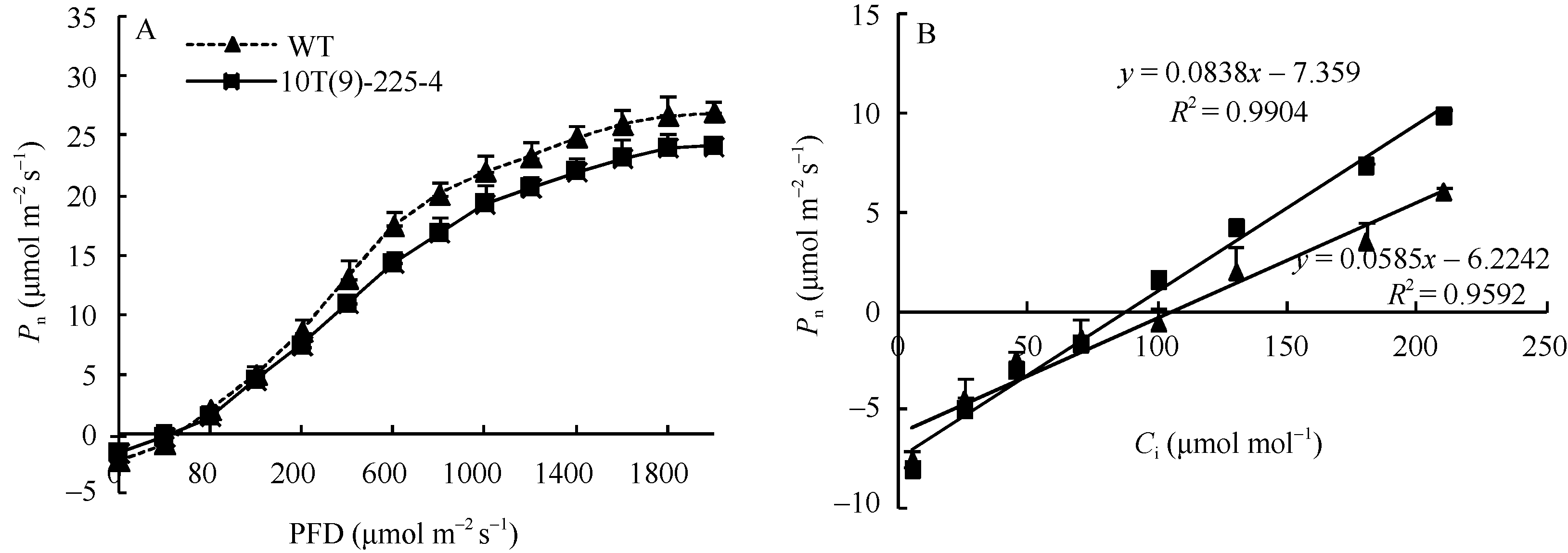 | 图7 转基因小麦旗叶净光合速率对光照强度(A)和胞间CO2浓度(C)的响应曲线Fig. 7 Response curves of photosynthesis to PPDF (A) and Ci (C) in transgenic wheat |
| 表4 转NADP-ME小麦和非转基因对照苹果酸、丙酮酸含量比较 Table 4 Comparison of malic acid and pyruvate acid content betweenNADP-ME transgenic wheat and WT |
 | 图8 400倍目镜下转基因小麦(A)和对照(B)旗叶气孔导度Fig. 8 Stomatal conductance of the flag leaves in transgenic wheat (A) and wild type (B) at a magnification of × 400 |
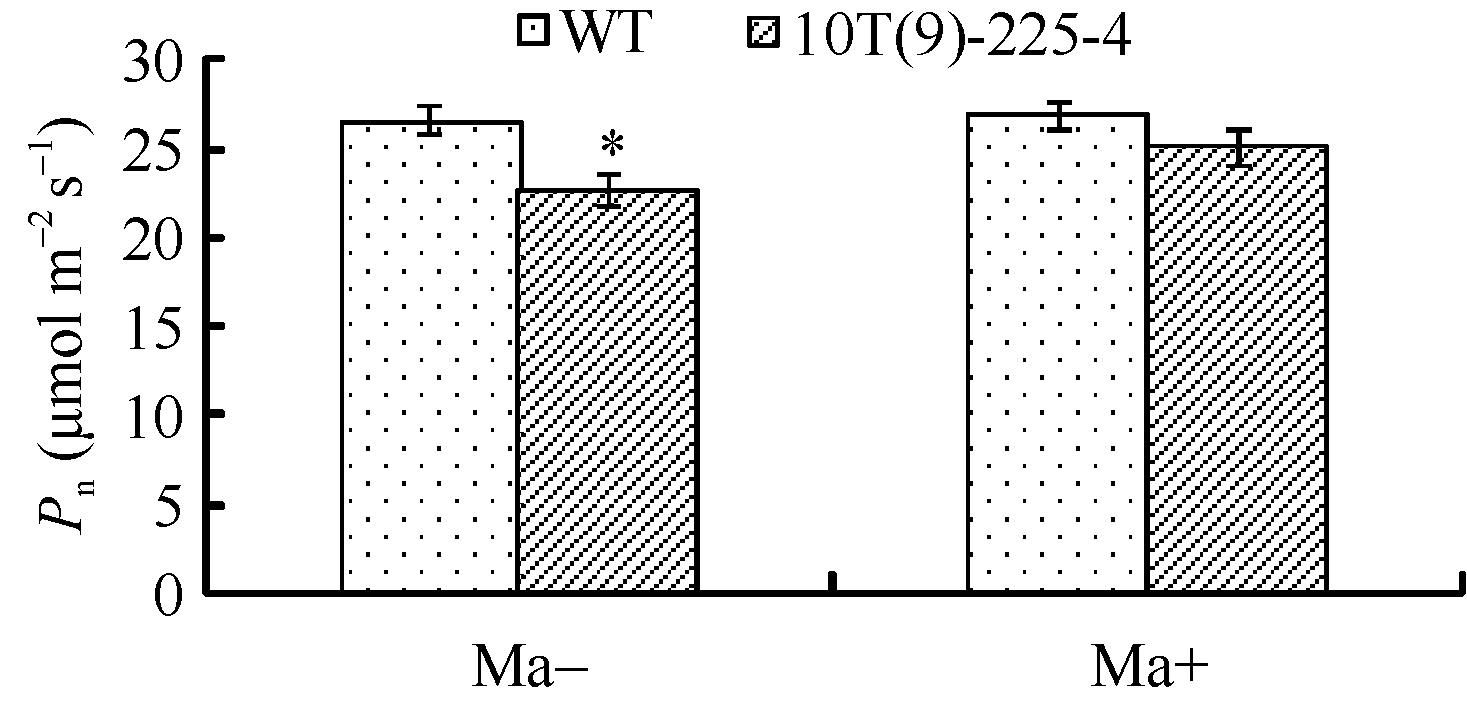 | 图9 外施苹果酸后转基因小麦和对照(WT)旗叶净光合速率比较Fig. 9 Dynamic changes of exogenous malate on photosynthetic rate in transgenic wheat and wild type (WT) |
自Ku等[5]利用农杆菌介导法获得转玉米全长PEPC基因水稻后, 世界各国研究人员开始将C4光合相关基因导入到C3植物中, 以期提高C3植物的光合生产力, 改善其产量潜力[31]。然而将C4光合关键酶基因引入C3植物中提高光合效率的研究一直备受争议[2, 7]。Hu等[13]研究表明, 转玉米C4型PEPC基因小麦光合高达31.95 μ mol m-2 s-1, 较对照提高26%。Takeuchi等[20]将玉米C4型NADP-ME基因全长cDNA导入水稻, 转化植株的NADP-ME活性提高了至少30倍, 叶绿体内高活性的NADP-ME可能影响了叶绿体发育。Jiao等[32]研究发现, 转玉米C4型NADP-ME基因水稻中NADP-ME的活性比对照提高了5倍, 达到玉米的50%, 但转基因水稻的光合效率仍然没有提高。迟伟等[22]的研究也得到相似的结果。本研究以T3代转NADP-ME基因小麦株系10T(9)-1-1和10T(9)-225-4为材料, 证实外源NADP-ME基因已整合到小麦基因组中, 并能够正常转录和翻译; 在4个测定生育期转基因株系的NADP-ME酶活性与对照周麦23相比均有所提高, 而净光合速率(Pn)则明显下降; 尤其在花后第7天, 2个转基因株系的酶活性分别较对照提高1.33倍和1.13倍, 净光合速率(Pn)分别下降17.26%和10.35%; 籽粒产量和千粒重较对照下降显著。
光合速率的提高是一个复杂的过程, 受诸多因素影响。有研究表明, 转PEPC基因水稻光合速率增加可能是由于PEPC过量表达促进了苹果酸的合成, 并通过苹果酸调节了气孔保卫细胞的有机酸代谢, 提高了气孔导度[6, 15]。迟伟等[21]研究发现, 转NADP-ME基因水稻在强光下叶绿体会积累大量还原性的NADPH, 过量的NADPH会造成大量活性氧的产生, 进而对光反应系统PSII的反应中心进行破坏, 使转基因植株光抑制作用增强。苹果酸和丙酮酸分别是NADP-ME酶反应的底物和产物(即: L-苹果酸+ NADP+→ 丙酮酸+CO2+ NADPH+H+), 而苹果酸在叶肉表皮细胞具有调节气孔开闭的作用。陈根云等[33]用外源C4光合原初产物OAA或MA饲喂C3菠菜叶片, 观察到其光合能力提高。朱素琴等[34]在水稻上也得到相似的结果。本研究中, 用150 mmol L-1的苹果酸溶液处理转基因小麦及对照旗叶后, 两者旗叶Pn均有所提高, 其中转基因小麦增幅最大(9.46%), 对照略微提高。对转基因株系10T(9)-225-4的光合效率降低的原因进行分析, 发现转NADP-ME基因小麦中NADP-ME活性提高, 但是其气孔张开率和气孔导度下降; 旗叶苹果酸含量显著低于对照周麦23, 而丙酮酸含量则显著提高; 外施苹果酸则可提高其Pn值, 与对照基本接近。由此可见, 转基因小麦气孔导度的下降可能是由于其苹果酸含量的降低造成的, 进而导致其光合效率下降, 而外施苹果酸可恢复其光合特性。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|